茉莉酸甲酯調控下人參發狀根皂苷合成相關基因表達的研究
2017-07-13 14:41:00王康宇于麗莉張美萍尹銳林彥萍趙
中國中藥雜志 2017年12期
王康宇+于麗莉+張美萍+尹銳+林彥萍+趙明珠+孫春玉+王義

[摘要] 通過外源添加茉莉酸甲酯(MeJA),研究其調控液體培養條件下人參發狀根中皂苷生物合成途徑相關酶基因在誘導子作用時表達情況。以四年生吉林人參主根誘導的人參發狀根無性系為材料,利用香草醛-硫酸比色法測定MeJA 處理前后人參發狀根的總皂苷含量;同時,利用熒光實時定量PCR測定MeJA 處理前后人參發狀根中鯊烯合成酶(squalene synthase,SQS)、鯊烯環氧酶(squalene epoxidase,SQE)、氧化鯊烯環化酶(oxidized squalene cyclase,OSC)、達瑪烷二醇合成酶(dammarenediol synthase,DS)、β-香樹脂合成酶(β-amyrin synthase,β-AS)和環阿屯醇合成酶(cycloartenol synthase,CAS)基因的相對表達量。結果表明:獲得MeJA添加到人參發狀根中最佳添加濃度為6×10-4μmol·L-1、添加時間為22 d、作用時間為5 d;MeJA的添加可以提高人參發狀根中過氧化物酶(PPD)、過氧化氫酶(CAT)和過氧化物酶(PPD)的酶活性;經過MeJA處理后人參發狀根中人參皂苷生物合成途徑的SQS,SQE,OSC,DS,β-AS基因的表達變化均有顯著提高,然而CAS基因的表達變化不顯著。因此說明在添加MeJA處理后,皂苷生物合成途徑中SQS,SQE,OSC,DS,β-AS基因表達情況與PPD,CAT,PPO的酶活性的變化趨勢也相一致。
[關鍵詞] 人參發狀根; 人參皂苷生物合成; 茉莉酸甲酯(MeJA); 基因表達
[Abstract] In order to obtain the expression of ginsenoside biosynthetic pathway related enzyme gene in ginseng hairy root under the control of elicitors, methyl jasmonate (MeJA) was added exogenously as elicitors. Ginseng hairy root clones induced by 4-year-old ginseng root was used as material, total saponin content in ginseng hairy root before and after MeJA treatment was determined by vanillin-sulfuric acid colorimetry, Meanwhile, relative expression of squalene synthase genes, squalene epoxidase genes, oxidized squalene cyclase genes, dammarenediol synthase genes, β-amyrin synthase genes, cycloartenol synthase genes before and after MeJA treatment were determined by Real-time PCR. The optimum conditions of MeJA which added to ginseng hairy root were obtained, the optimum additional concentration was 6×10-4μmol·L-1, the optimum additional time was 22 d, and the optimum action time was 5 d. The addition of MeJA could improve the enzymatic activity of peroxidase (PPD), catalase (CAT) and peroxidase (PPD) in ginseng hairy root. The expression of SQS,SQE,OSC,DS and β-AS genes of ginsenoside biosynthetic pathway increased significantly after MeJA treatment, while the change of CAS gene expression were not significant. The expression of key enzyme SQS,SQE,OSC,DS and β-AS genes in ginsenoside biosynthetic pathway was consistent with the changes of PPD,CAT,PPO enzymatic activity.
[Key words] ginseng hairy root; ginsenoside biosynthesis; methyl jasmonic acid(MeJA); gene expression
人參Panax ginseng C. A. Meyer是馳名中外的五加科人參屬植物,聞名遐邇的“東北三寶”之一[1]。吉林省是我國人參的主要產地,占全球人參總產量的70%,人參產業不僅是吉林省重要的經濟組成部分,更是我國國民經濟中最具特色的新興產業之一。人參中的主要活性成分為人參皂苷,按其化學性質可分為:人參皂苷二醇型(A型),包括Ra,Rbl,Rb2,Rc,Rd,Rh2等;人參皂苷三醇型(B型),包括Re,Rf,Rg1,Rg2,Rh1等;齊墩果酸型(C型),如Ro[2-3]。按苷元的基本類型可以分為:達瑪烷型、齊墩果酸型和奧克梯隆型[3-4]。人參組織培養中人參發狀根具有生長速度快,周期短,皂苷含量較高等特點,似的利用人參發狀根組織培養作為細胞工廠定向生產人參皂苷的研究已經成為熱點問題。但目前,我國對人參發狀根的研究主要集中在發狀根的誘導[5-7]、有效成分的提取分析[8-9],關于人參皂苷生物合成途徑中調控皂苷合成的關鍵酶在人參發狀根中的研究甚少。
研究表明茉莉酸類化合物可以在植物生長發育過程起到脅迫信號的重要作用[10]。外源性添加茉莉酸類物質(茉莉酸或茉莉酸甲酯等)不僅對植物的生長起作用,也能顯著的影響植物的次級代謝活動[11-12]。當外源添加時激活信號轉導通路,迫使植物產生應激反應,產生大量次生代謝產物應對各種脅迫病害[14-16]。因此,目前使用茉莉酸甲酯等小分子化合物作為誘導子成為了研究的熱門方法,刺激植物合成大量具有商業價值的次生代謝產物[13,17]。
本研究以吉林人參為材料,誘導和篩選獲得人參發狀根無性系,通過篩選MeJA調控人參發狀根生物合成人參皂苷的最佳作用濃度,最佳添加時間、最佳作用時間的條件,從而研究MeJA調控人參皂苷生物合成途徑中重要的相關酶基因的表達情況。利用熒光實時定量PCR技術實時檢測MeJA調控下人參皂苷生物合成途徑中重要相關酶基因的表達,不僅可以為研究誘導子調控人參皂苷的生物合成提供數據支持,更可以為人參發狀根體系作為細胞工廠定向生產人參皂苷提供理論依據和技術手段。
1 材料
實驗材料為四年生人參主根誘導獲得的人參發狀根,由吉林農業大學-吉林省人參基因資源開發與利用工程研究中心提供。
人參皂苷Rg1單體對照品(批號110754-200826)采購于中檢所、茉莉酸甲酯購于美國Sigma公司、RevertAidTM first strand cDNA Synthesis Kit反轉錄試劑盒采購于美國MBI公司、熒光實時定量qPCR試劑盒購于德國Qiagen公司、其他試劑均購于百泰克公司和北京化工廠(分析純)。
熒光實時定量PCR擴增儀為美國ABI公司的7500型、高速冷凍離心機為美國Thermo公司的Fresco21型、恒溫振蕩培養箱為哈東聯公司的HZQ-F160型、超微量核酸蛋白濃度檢測儀為德國耶拿公司的Scandrop250型。
2 方法
2.1 篩選吉林人參發狀根無性系
配置1/2MS固體培養基,倒入培養皿中,在培養皿背面畫1.0 cm×1.0 cm的正方格用于測量人參發狀根生長速度。選取生長良好的人參發狀根切取發狀根根尖部位1.0 cm長度,接于畫好正方格的平板上于黑暗處培養。選擇生長狀況良好的,接種于1/2MS液體培養基中,培養條件為22 ℃,110 r·min-1振蕩培養,以備后續實驗。
2.2 茉莉酸甲酯(MeJA)的制備及添加
利用本實驗室條件下構建好的人參發狀根動力培養模型[18],分別考察MeJA最佳添加濃度、時間、作用時間對人參發狀根的影響。選擇配置MeJA溶液的基礎濃度為200 μmol·L-1,設置7個添加濃度梯度分別為0,1,2,3,4,5,6 μL,篩選出MeJA最佳添加濃度;設置7個添加時間梯度分別為第14,16,18,20,22,24,26天,添加已經篩選出最佳添加濃度的MeJA,篩選出MeJA最佳添加時間;在已經確定MeJA最佳添加濃度和最佳添加時間的條件下,設置7個MeJA作用時間分別為1,2,3,4,5,6,7 d,作用時間過后更換新鮮1/2MS培養基后繼續培養,篩選出MeJA最佳作用時間。每次篩選實驗時,均設置相同條件下不做任何處理的5瓶對照組,篩選實驗重復3次。
2.3 人參發狀根生長量及皂苷含量的測定
以干、鮮重為指標進行發狀根生長量的測定,干重為將新鮮材料取出后置于40 ℃以下條件的烘箱中烘干至恒重。
鮮重增加量(g/瓶)=收獲時鮮重(g/瓶)- 接種時鮮重(g/瓶)
干重增加量(g/瓶)=收獲時干重(g/瓶)- 接種時干重(g/瓶)
人參發狀根中總皂苷提取方法是利用的索氏回流提取法、總皂苷含量檢測方法是使用的硫酸香草醛-硫酸比色法 [18-19]。以人參皂苷Rg1對照品質量濃度為橫坐標,不同濃度對照品吸光度為縱坐標,繪制人參皂苷的標準曲線,并獲得標準曲線回歸方程為Y=0.230 7X+0.002 7,R2=0.996 8,從而可以檢測出待測樣本中皂苷含量。
2.4 過氧化物酶(PPD)、多酚氧化酶(PPO)、過氧化氫酶(CAT)活力的測定
稱取1.0 g發狀根于研缽中,加入2 mL磷酸緩沖液(PPO活力pH 8.0,POD活力pH 5.5,CAT活力pH 7.8)充分研磨后,將勻漿后轉入離心管中,再次向研缽中加入1.5 mL磷酸緩沖液,充分沖洗研缽后,將其轉入離心管中,8 000 r·min-1離心10 min(POD,CAT),4 000 r·min-1離心15 min(PPO),取上清液3 mL至于干凈試管中。向空白管和測定管中依次加入0.5 mol·L-1磷酸緩沖液2.5 mL(POD活力pH 5.5),4 mL(PPO活力pH 8.0),3 mL(CAT活力pH 7.8)。隨后分別對POD,CAT,PPO活力進行測定。
2.4.1 POD活力測定[20] 在已經準備好的預處理溶液中加入1.0 mL 2% H2O2溶液、1.0 mL 0.5 mol·L-1愈創木酚溶液和0.5 mL酶溶液。先將對照組置于沸水浴中5 min使加入的酶失活,然后對照組和待測組同時置于37 ℃水浴處理10 min,取出后分別加入2 mL 20% TCA溶液終止反應,最在470 nm處檢測A,并計算POD活力。POD活力=ΔA470 nm /(0.01×T) ×D(T為反應時間;D為稀釋倍數)。
2.4.2 PPO活力測定[20] 在已經準備好的預處理溶液中加入1 mL 0.2 mol·L-1兒茶酚溶液,向對照組和待測組中分別加入1 mL酶溶液。置于37 ℃水浴處理10 min,取出后立即加2 mL 20% TCA溶液混勻終止反應,最后在525 nm處檢測A,并計算PPO活力。PPO活力=ΔA525 nm/(0.01×t) ×D(T為反應時間;D為稀釋倍數)。
2.4.3 CAT活力測定[20] 在已經準備好的預處理溶液中加入2 mL蒸餾水,在待測組中加入0.4 mL酶溶液,置于25 ℃水浴中預熱處理3 min后加入0.6 mL 0.1 mol·L-1的H2O2,最后在240 nm處檢測A,并計算CAT活力。CAT活性=(ΔA240×VT)/(0.1×V1×t×FW)(VT為粗酶溶液總體積;V1為使用粗酶溶液體積;FW為待測樣本鮮重;t為反應時間)。
2.5 MeJA對人參發狀根中皂苷生物合成相關基因表達的影響
2.5.1 吉林人參發狀根的總RNA的提取 利用Trizol法提取吉林人參發狀根的總RNA。對總RNA使用RNA純化試劑盒進行純化處理,并將其回溶于20 μL DEPC處理過的ddH2O中,最后利用核酸濃度檢測儀Scandrop250測定RNA濃度。
2.5.2 人參發狀根RNA的反轉錄 利用美國MBI公司的RevertAidTM first strand cDNA Synthesis Kit反轉錄試劑盒,將Total RNA反轉錄為cDNA,保存備用。
2.5.3 實時定量PCR的引物設計與合成 利用Vector NTI Advance 11.0軟件設計GADPH,SQS,SQE,OSC,DS,β-AS,CAS基因的實時定量PCR擴增引物,并委托由華大基因(BGI)公司完成合成,引物序列見表1。
2.5.4 熒光實時定量PCR 熒光實時定量PCR反應體系參照試劑盒,根據常規PCR結果設置反應參數,每個模板3次重復。PCR循環參數為:預變性 95 ℃ 10 min;變性 95 ℃ 15 s;退火60 ℃ 1 min;延伸72 ℃ 30 s。將熒光實時定量PCR過程中收集的全部數據,使用美國ABI公司提供的SDS V2.3軟件進行系統分析,并計算2-ΔΔCT,獲得每個基因的相對表達量。
3 結果與分析
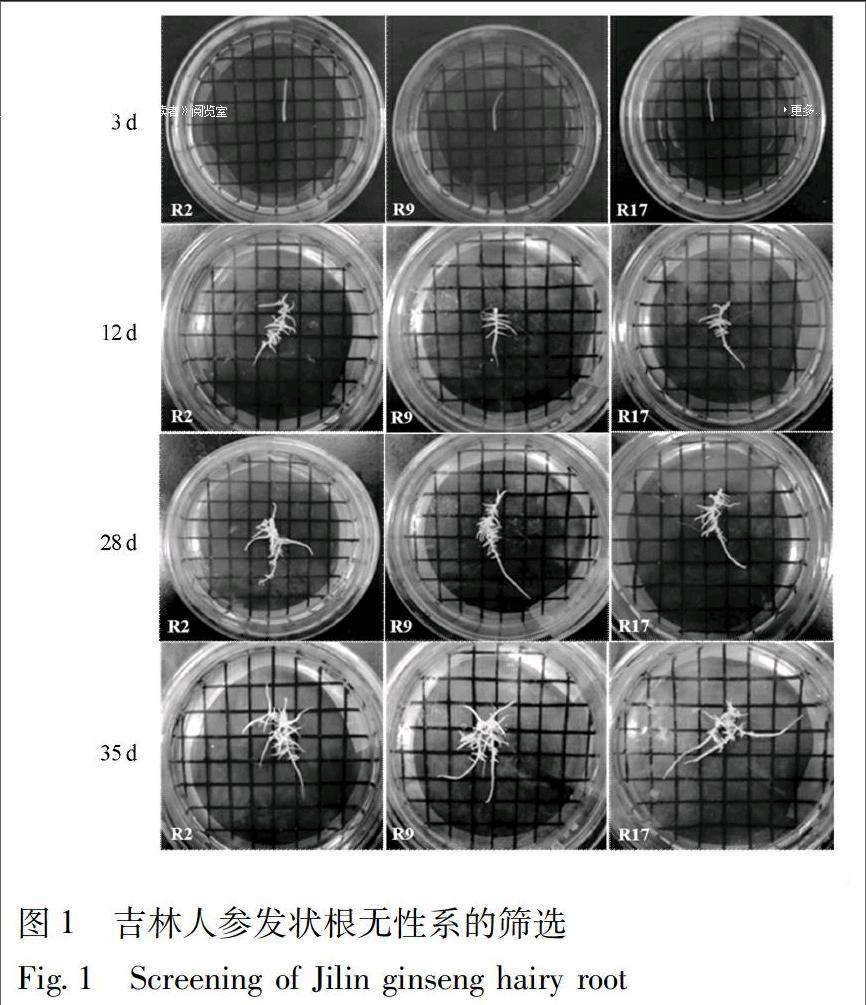
3.1 吉林人參發狀根無性系的篩選
本實驗對誘導獲得的吉林人參的發狀根通過單根培養,共篩選出23個人參發狀根無性系,并將其命名為R1-R23。當培養到第3天時,人參發狀根在1/2MS平板培養中開始有明顯伸長;當培養到第12天時,人參發狀根的伸長長度約為剛培養時長度的2倍,且出現較多分枝;當培養到第28天時,人參發狀根伸長長度約為剛培養時長度的4倍,且出現根系的分枝上再次分枝,伴隨愈傷化現象出現;當培養到第35天,發狀根的分枝可達到30根以上,見圖1。綜合比對結果,人參發狀根系無性系R9在35 d培養周期中,其生長速度較快,能夠產生較多分枝,主根與分枝均呈現較高透明程度,并且顏色為乳白色,愈傷化程度較低,與其他22個人參發狀根無性系相比優勢顯著,因此,確定R9人參發狀根無性系為最優材料,并將其轉移到1/2MS液體培養基中在22 ℃,150 r·min-1,暗培養條件下繼續培養,以備后續實驗使用。
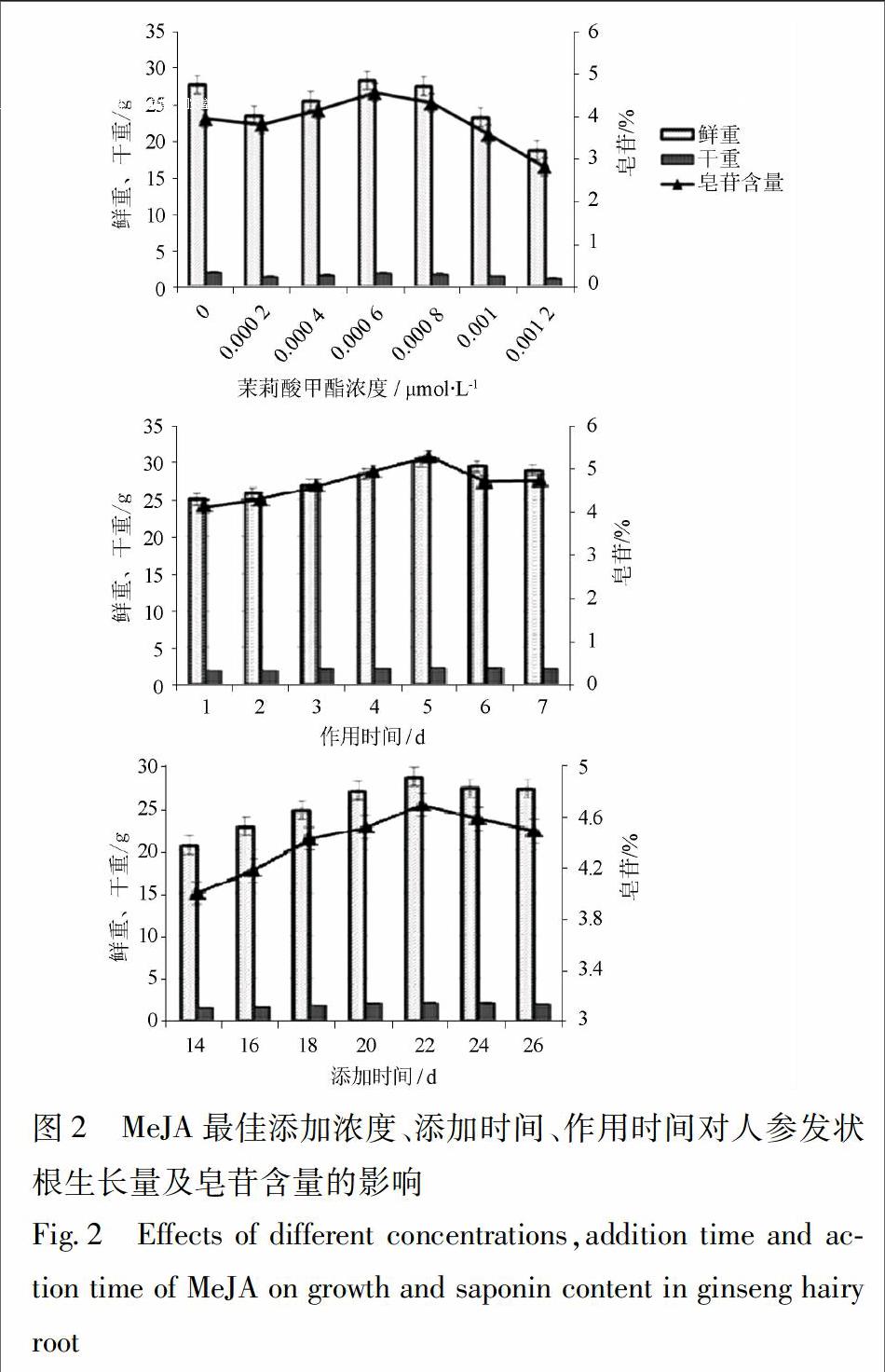
3.2 MeJA對人參發狀根生長量與皂苷含量的影響
在篩選最佳添加濃度、添加時間和作用時間時MeJA對人參發狀根的生長量、皂苷含量的影響顯著,最佳條件下處理組的生長量和皂苷含量高于對照組。本實驗通過篩選獲得MeJA最佳添加濃度為6×10-4μmol·L-1、最佳添加時間為22 d,最佳作用時間為5 d,見圖2。
3.3 MeJA對人參發狀根中PPD,PPO,CAT 3種酶活力的影響
在MeJA的最佳添加條件下,分別檢測了POD,CAT,PPO的酶活性,見圖3。結果顯示:POD酶活力在第6小時出現第1個小峰值。隨著MeJA作用時間的增加,POD酶活力也升高,當POD酶活力在第20小時達到了最大值,測試組POD酶活力比對照組高1.40倍。隨著作用時間繼續增加,POD酶活力不斷降低,當36 h后時POD酶活力與對照組的基本接近。PPO酶活力隨著MeJA作用時間增加逐漸升高,當到第18小時 PPO的活力達到了最大值,隨著酶活力開始下降,24 h后PPO酶活力與對照組的逐漸接近。CAT酶活力在第6小時出現第1個小峰值,隨著MeJA作用時間的不斷增加,測試組CAT酶活力也不斷升高,CAT酶活力在第14小時達到了最大值,測試組CAT酶活力比對照組高1.43倍。隨著作用時間不斷增加,CAT酶活力開始逐漸降低,24 h后CAT酶活力與對照組的基本接近。因此,POD,PPO,CAT這3種作為保護植物面對外界刺激啟動應激反應的酶類,同時還可以清除應激反應引發的活性氧。隨著MeJA在調控人參發狀根的作用時間的不斷增加,POD,CAT,PPO這3種保護酶類的活性也隨之升高,清除MeJA刺激人參發狀根應激反應產生活性氧能力也不斷提升,最終可以起到保護發狀根細胞的膜結構,從而增加人參發狀根對外界的抗性。
3.4 MeJA調控人參發狀根中皂苷生物合成相關基因表達研究
本實驗研究了MeJA對人參發狀根中人參皂苷生物合成的SQS,SQE,OSC,DS,β-AS,CAS基因表達的影響,見圖4。0 h作用的作為未經處理的對照組,經過MeJA處理后的人參發狀根與對照組相比,SQS,SQE,OSC,DS,β-AS基因的表達變化均有顯著提高。結果表明:在添加MeJA調控后第12小時,SQS基因的表達量達到最大值,高于對照組8.2倍;在添加MeJA調控后第24小時,SQE基因的表達量達到最大值,高于對照組11.04倍;在添加MeJA調控后第4小時,OSC基因的表達量呈現小幅度的上升,隨后又呈現下降,在調控后第8小時,OSC基因表達量再次呈現上升,直到第16小時OSC基因的表達量達到最大值,高于對照組11.38倍,隨著作用時間繼續增加,其表達量逐步下降,但始終高于對照組。在添加MeJA調控后第20小時,DS和β-AS基因的表達量分別達到最大值,且分別高于對照組13.91,12.59倍。然而在添加MeJA調控后的人參發狀根與未經處理的對照組發狀根相比CAS基因的表達量變化不顯著。
4 討論
外源性添加茉莉酸類物質(茉莉酸或茉莉酸甲酯等)不僅對植物的生長起作用,也能顯著的影響植物的次級代謝活動。當外源添加時激活信號轉導通路,迫使植物產生應激反應,產生大量次生代謝產物應對各種脅迫病害[10-16]。因此,目前使用茉莉酸甲酯等小分子化合物作為誘導子成為了研究的熱門方法,刺激植物合成大量具有商業價值的次生代謝產物。
本實驗通過對吉林人參發狀根外源添加MeJA作為誘導子,從結果可以發現:在添加MeJA調控人參發狀根液體懸浮培養下,不僅可以促進懸浮培養條件下的人參發狀根的生長,更可以定向影響人參發狀根中皂苷生物合成。在MeJA添加刺激作用下,人參皂苷生物合成途徑中SQS,SQE,OSC,DS,β-AS,CAS等關鍵酶基因的表達是一種動態的變化,通過MeJA刺激作用SQS,SQE,OSC,DS,β-AS基因的表達能發生快速響應,在添加作用6 h后,這5種關鍵酶基因的表達迅速升高。黃超等[21]利用釩酸鹽小分子化合物作為誘導子,添加到人參細胞懸浮培養物中,研究發現:處理組中SQS,SQE,DS基因的表達量分別高于對照組7.6,10.3,17.8倍,同時處理組中CAS基因的表達量與對照組相比變化不顯著。該實驗結果與本實驗得出的處理組中SQS,SQE,OSC,DS,β-AS基因的表達量分別高于對照組的結論基本相一致。因此,在添加MeJA處理后,人參皂苷生物合成途徑中關鍵酶SQS,SQE,OSC,DS,β-AS基因的表達與總皂苷含量變化的趨勢相一致。
在人參皂苷生物合成代謝過程中,SQS基因被認定是一個起十分關鍵作用的限速酶基因,是人參皂苷合成的關鍵調控位點,控制皂苷中碳元素的流向。通過添加MeJA刺激時,SQE基因的表達比SQS基因的表達具有更高的持續性和更強的表達作用,推測當受到MeJA刺激時,SQE基因在人參皂苷合成途徑中的關鍵作用比SQS基因更為重要。DS和β-AS基因表達量的最高峰值出現于MeJA刺激后的20 h左右,比SQS,SQE,OSC基因表達最大峰值稍晚,見圖5。此外,CAS基因是萜類化合物中甾醇類中的相關基因,從而出現表達量變化平穩,變化不顯著。因此,這些基因表達規律的結果體現了人參皂苷生物合成代謝過程中關鍵酶基因表達存在著空間時序性的相關理論。
外源添加MeJA作為誘導子,可以引發植物的防御反應。本實驗結果發現,在添加MeJA 調控作用6 h后PPD與CAT的酶活力在開始升高,當添加20 h和14 h后分別達到酶活力的最大峰值,隨著添加MeJA調控時間繼續增長,到達36~72 h時測試組中PPD與CAT的酶活力逐漸恢復到與對照組一致的水平;在添加MeJA 調控作后從開始到第22小時測試組中PPO的酶活力始終高于對照組,達到24 h時測試組中PPO的酶活力逐漸恢復到與對照組一致的水平。
同時,在人參發狀根關鍵酶基因的表達方面也有著變化,在添加MeJA調控作用的第6~8小時中,人參皂苷生物合成代謝途徑中的SQS,DS,β-AS關鍵酶基因的表達量開始升高,當作用時間達到12~14 h這3個基因的表達量處于最大峰值,隨后開始逐漸下降;在添加MeJA調控作用的第2~24小時中,OSC關鍵酶基因表達量始終高于對照組,分別在第4和第16小時表達量出現峰值,在添加24 h后開始逐漸降低;在添加MeJA調控作用的第12~24小時中,SQE關鍵酶基因表達量始終于高于對照組,當作用時間達到第24小時表達量出現最大峰值,隨后開始下降。因此,在添加MeJA調控人參發狀根后,PPD,CAT和PPO的酶活力變化趨勢與人參皂苷生物合成代謝途徑中關鍵酶SQS,SQE,OSC,DS,β-AS基因的表達量變化趨勢是相一致的。
[參考文獻]
[1] Taik K Y, Yun S L, You H L, et al. Anticarcingenie effect of Panax ginseng C.A. Meyer and identification of active compounds[J]. J Korean Med Sci, 2001,16:6
[2] 王菡,富強,王麗思,等. 人參皂苷生物合成的相關酶及其基因的研究進展[J]. 人參研究,2012(1):40.
[3] Kim J Y, Zhang D, Yang D C. Biosynthesis and biotechnological production of ginsenosides[J]. Biotechnol Adv,2015,33:717.
[4] 李平亞.人參皂苷NMR標準圖譜[M].北京:化學工業出版社,2012.
[5] 李天航,楊晶,王艷芳,等. 發狀根在藥用植物基因工程研究進展[C].天津: 2010年中國藥學大會暨第十屆中國藥師周,2010.
[6] 張獻龍,唐克軒.植物生物技術[M].北京:科學出版社,2004:279.
[7] 王建華,薛健,侯春喜,等.人參發狀根的愈傷組織誘導[J].吉林農業大學學報,2008,30(5):700.
[8] 羅玉.利用毛狀根的培養生產植物次生代謝物[J].云南農業科技,2002(2): 40.
[9] 張興,劉曉娟,呂巧玲.毛狀根生產次生代謝產物的研究進展[J].化工進展,2007,26(9):1228.
[10] 蔣科技,皮妍,侯嶸,等.植物內源茉莉酸類物質的生物合成途徑及其生物學意義[J].植物學報,2010,45 (2):137.
[11] Howe G A. Jasmonates as signals in the wound response[J]. J Plant Growth Regul, 2004, 23: 223.
[12] Wasternack C, Stenzel I, Hause B, et al. The wound response in tomato-role of jasmonic acid[J]. J Plant Physiol, 2006, 163:297.
[13] M Kawauchi, T Arima, O Shirota, et al. Production of sesquiterpene-type phytoalexins by hairy roots of HyOS Cyamus albus co-treated with cupper sulfate and methyl jasmonate[J]. Chem Pharm Bull, 2010, 58: 934.
[14] 蔡昆爭,董桃杏,徐濤.茉莉酸類物質(JAs) 的生理特性及其在逆境脅迫中的抗性作用[J].生態環境,2006,15(2):397.
[15] 傅俊范,孫嘉曼,周如軍,等.茉莉酸甲酯對人參β-1,3-葡聚糖酶和幾丁質酶活性的影響[J].中國農學通報,2012,28(9):214.
[16] 張智慧,聶燕芳,何磊,等.外源茉莉酸甲酯誘導水稻抗瘟性相關防御酶和內源水楊酸的變化[J].植物病理學報,2010,40(4):395.
[17] 徐立新,趙壽經,梁彥龍,等.外源調節物質對人參毛狀根生長及皂苷合成的影響[J].吉林大學學報:工學版,2010,40(6):1619.
[18] 張丹.人參發狀根懸浮培養體系的建立及發酵動力學研究[D].長春:吉林農業大學,2012.
[19] 蓋玉紅,王文慧,王瀝浩,等.人參發狀根與栽培人參有效成分的比較研究[J].北方園藝,2013(7):166.
[20] 王穎.前體物質對西洋參愈傷組織生長及代謝的影響研究[D].長春:吉林農業大學,2007.
[21] 黃超.小分子化合物的添加提高人參皂苷產量及其作用機制探索[D].上海:華東理工大學,2013.
[責任編輯 呂冬梅]
