東方粘蟲 U6啟動子的克隆及功能驗證
2017-07-18 11:26:39王高振吳文君祁志軍
西北農業學報 2017年6期
王高振,金 朵,吳文君,祁志軍
(西北農林科技大學 植物保護學院農藥研究所,陜西楊凌 712100)
東方粘蟲 U6啟動子的克隆及功能驗證
王高振,金 朵,吳文君,祁志軍
(西北農林科技大學 植物保護學院農藥研究所,陜西楊凌 712100)
為了持續獲得大量短片段干擾RNA(short interference RNA, siRNA),通過染色體步移技術克隆得到東方粘蟲 U6 snRNA 基因5′-端側翼啟動子序列1 426 bp。經生物信息學分析,該序列含SPH元件、八聚體序列(OCT)、遠端序列元件(DSE)、近端序列元件(PSE)及TATA box等RNA聚合酶Ⅲ(RNA polymerase Ⅲ,RNA Pol Ⅲ) U6啟動子的特征元件。通過構建RNA干擾(RNA interference, RNAi)載體的方式,對增強型綠色熒光蛋白EGFP基因進行RNAi,證明克隆得到的東方粘蟲 U6啟動子可成功驅動shEGFP表達,具有 U6啟動子功能。為后續構建以東方粘蟲V-ATP酶H亞基為靶基因的沉默載體奠定基礎。
東方粘蟲;RNA干擾; U6啟動子;功能驗證
RNA干擾(RNA interference, RNAi)是近年來快速發展起來的一種序列特異的轉錄后基因沉默技術[1],在功能基因組研究[2-3]、疾病防治[4-5]及新藥研發[6-7]等方面具有廣闊的應用前景。其基本原理是短片段干擾RNA(short interference RNA, siRNA)與細胞內同源的mRNA結合導致其降解,從而使目的基因表達沉默[8]。獲得siRNA的方法主要有化學合成法、Dice酶切割法和短片段發卡RNA(short-hairpin RNA, shRNA)表達載體法[9]等。其中,構建shRNA表達載體的方法是唯一適合長期研究的方法,而且由于質粒可以復制擴增,相比起化學合成來說,能夠顯著降低制備siRNA的成本[10]。目前,介導siRNA表達的啟動子主要是RNA聚合酶Ⅲ(RNA polymerase Ⅲ, RNA pol Ⅲ)啟動子,常用人源和鼠源的 U6啟動子及人的H1啟動子等[11]。這類啟動子可以促進快速合成大量shRNA,可以實現對細胞內基因的快速沉默[12]。RNA pol Ⅲ啟動子序列上包含明確的起始和終止序列,轉錄出的RNA形成發夾結構會在3′-端形成2個突出的尿嘧啶,這類似于天然的siRNA,因而有利于誘發RNA沉默[13]。目前,通過構建基于 U6啟動子的基因沉默載體在不同的細胞和生物體內已經取得了良好的基因沉默效果。例如,由家蠶 U6啟動子驅動的shRNA表達載體轉染家蠶BmN細胞后,顯著降低家蠶核多角體病毒對細胞的傷害[14];河豚雙 U6啟動子雙鏈RNA(double-stranded, dsRNA)表達載體轉染鮭魚胚胎CHSE-214細胞后,有效抑制敗血癥病毒在該細胞內的擴散[15];CMV-U6混合啟動子shRNA表達載體介導斑馬魚相關基因的沉默[16]。由此可見,基于 U6啟動子的RNAi技術已經成為基因沉默的主要手段。
目前,人們對 U6啟動子的結構已經研究較為清楚,發現在轉錄起始位點上游包含3個啟動子元件:DSE、PSE和TATA box[17]。Lambeth等[18]研究發現 U6啟動子各個元件的間距、序列和結構之間存在種屬差異;Wise等[19]通過比較同一載體上的鼠源和雞 U6啟動子,發現不同啟動子的序列和結構對啟動效率有很大影響。也就是說某一物種的 U6啟動子對其他物種體內RNA pol Ⅲ的驅動能力可能降低甚至喪失。因此,要獲得理想的啟動效果,利用物種本源 U6啟動子是最佳選擇。
東方粘蟲 (Mythimnaseparata(Walker)) 是一種鱗翅目害蟲,其幼蟲主要危害玉米和小麥等禾本科作物,具有遠距離遷飛、暴食和集中危害等特點。嚴重發生時,短時間內就可將成片的玉米、谷子、水稻吃成根茬,防治不及時會造成嚴重損失,甚至絕產[20]。目前,防治東方粘蟲以噴灑殺蟲劑為主,但大面積使用化學農藥,不但造成農藥殘留、殺傷害蟲天敵,而且會導致害蟲產生抗藥性。近年來,隨著基因沉默技術的發展,基于RNAi的害蟲防治策略逐漸受到人們的關注[21],已經在玉米根葉甲(DiabroticavirgiferavirgiferaLeConte)[22]和煙粉虱(Bemisiatabaci)[23]等害蟲防治上獲得成功。在前期研究中,本實驗室克隆出東方粘蟲V-ATP酶H亞基,設計合成dsRNA并微量注射3齡幼蟲,試蟲出現生長發育受阻和死亡等典型癥狀[24]。為了持續獲得大量的siRNA,本研究克隆出東方粘蟲的 U6啟動子并對其進行功能驗證,為后續構建以東方粘蟲V-ATP酶H亞基為靶基因的沉默載體奠定基礎。
1 材料與方法
1.1 供試材料與試劑
東方粘蟲,在溫度為(22±1) ℃,相對濕度為60%~70%,光周期為L∶D=16 h∶8 h,光強為30 000 lx的條件下室內人工飼養,試驗中選取5齡初蛻幼蟲。293 T細胞系由西北農林科技大學農藥研究所提供,并在含胎牛血清(φ=10%)、0.1 mg/mL硫酸鏈霉素和100 U/mL青霉素的DMEM高糖培養基中置于37 ℃、φ=5% CO2生化培養箱中培養,取對數生長期的細胞進行試驗。
Genome walker universal試劑盒及pEGFP-N1載體購自Clontech公司,T4DNA連接酶、pGEM-T載體等購自Promega公司,ExTaqDNA聚合酶(5 U/μL)、10×ExTaqbuffer (20 mmol/L, Mg2+plus)、dNTP Mixture(2.5 mmol/L)、DNA marker、pMD18-T載體、DH5α感受態細胞等購自TaKaRa公司;限制性內切酶EcoRⅠ、BamHⅠ、HindⅢ購自NEB公司;脂質體Lipofectamine 2000、DMEM高糖培養基、胎牛血清等購自Invitrogen公司;凝膠回收純化試劑盒、質粒提取試劑盒、昆蟲基因組DNA提取試劑盒、無內毒素質粒提取試劑盒等購自Omega公司;24孔細胞培養板購自ThermoFisher scientific公司;引物及測序均由上海英駿生物技術有限公司完成。
1.2 東方粘蟲基因組DNA的提取
基因組DNA的提取按照昆蟲基因組DNA提取試劑盒說明書進行。
1.3 染色體步移引物設計與合成
將家蠶 U6 snRNA基因全序列(AY649381.1)與果蠅的 U6 snRNA基因全序列(AH004871.1)經在線軟件Clustal Omega(http://www.ebi.ac.uk/tools/msa/clustalo/)比對,分析它們的同源區域并設計兩條下游引物U6R1 和 U6R2。上游兩條引物使用Genome walker universal試劑盒中的AP1和AP2。
引物序列如下:
AP1:5′-GTAATACGACTCACTATAGG-GC-3′;AP2:5′-ACTATAGGGCACGCGTGG- T-3′;U6R1:5′-CGATTTTGCGTGTCATCCTTGCGCAG-3′;U6R2:5′-TGCCTGGCTGTAAGTGCTCAGATTCC-3′。
在用戶擔心余額寶的自身潛在風險的時,阿里巴巴正在努力加強用戶的資金提供安全和保障。例如發表承諾,如果客戶賬戶資金被盜,支付寶方面將承諾全額賠償。
1.4 染色體步移文庫的建立
將已測定質量濃度的東方粘蟲基因組DNA經4種限制性內切酶DraⅠ、EcoR Ⅴ、StuⅠ和PvuⅡ過夜酶切,消化基因組DNA,回收純化消化產物。將純化后的DNA與Genome walker 接頭于16 ℃過夜連接,70 ℃下5 min終止反應。用TE緩沖液稀釋至一定質量濃度作為PCR擴增的模板。
1.5 U6 snRNA基因上游5′-端側翼序列的克隆
以構建好的染色體步移文庫為模板,分別以AP1/U6R1和AP2/U6R2為引物,進行兩輪巢式PCR。PCR程序按照染色體步移試劑盒說明書進行。將4個不同文庫對應的兩輪PCR產物經10 g/L瓊脂糖凝膠電泳分析,并將目的條帶用凝膠回收試劑盒回收純化,再將其連接至pGEM-T載體上,轉化至大腸桿菌DH5α感受態細胞中,對陽性克隆測序分析。
1.6 U6啟動子的shEGFP重組載體的構建
1.6.1 pMD18-N載體的構建 根據pMD18-T載體圖譜,通過T-A克隆導入一段不包含EcoRⅠ、BamHⅠ及HindⅢ酶切位點的片段N(515 bp),經雙酶切檢測并測序。
1.6.3 pMD18-U6與pMD18-shEGFP載體的構建 根據染色體步移法克隆得到的 U6 snRNA上游5′-端側翼序列,設計上游引物F1:GGATCCCTAGTGCACGGTTCCAAATTGGT(斜體為BamHⅠ酶切位點),下游引物R1 :AAGCTTCATGAAAACTTTACTTGCGCGA(斜體為Hind Ⅲ酶切位點),以東方粘蟲基因組DNA為模板,克隆得到上游序列1 360 bp,即為包含BamHⅠ、Hind Ⅲ酶切位點的 U6啟動子。將得到的pMD18-N載體經BamHⅠ、Hind Ⅲ酶切,并用凝膠回收試劑盒回收,回收產物分別與 U6啟動子及獲得的雙鏈shRNA連接,得到pMD18-U6與pMD18-shEGFP載體,并送上海英駿生物技術有限公司測序。
1.6.4 pMD18-U6-shEGFP載體的構建 按照“1.6.3”方法,設計上游引物F2:5′-GAATTCCTAGTGCACGGTTCCAAATTGGT-3′(斜體為EcoRⅠ酶切位點),下游引物R2 :5′-GGATCCCATGAAAACTTTACTTGCGCGA-3′(斜體為BamHⅠ酶切位點),克隆得到包含EcoRⅠ、BamHⅠ酶切位點的 U6啟動子。將pMD18-N載體經EcoRⅠ、BamHⅠ酶切,并用凝膠回收試劑盒回收,回收產物與 U6啟動子連接,得到pMD18-U6-N質粒;將pMD18-U6-N經BamHⅠ、HindⅢ酶切,凝膠回收后與shRNA連接,得到 pMD18-U6-shEGFP載體,并送上海英駿生物技術有限公司測序。
1.7 細胞轉染
將處于對數期的293 T細胞鋪至24孔板內培養,在倒置顯微鏡下觀察,當細胞融合度達70%左右時進行下一步試驗。將構建好的pMD18-U6-shEGFP、pMD18-U6、pMD18-shEGFP載體分別與pEGFP-N1載體共轉染293 T細胞。具體方法見表1。
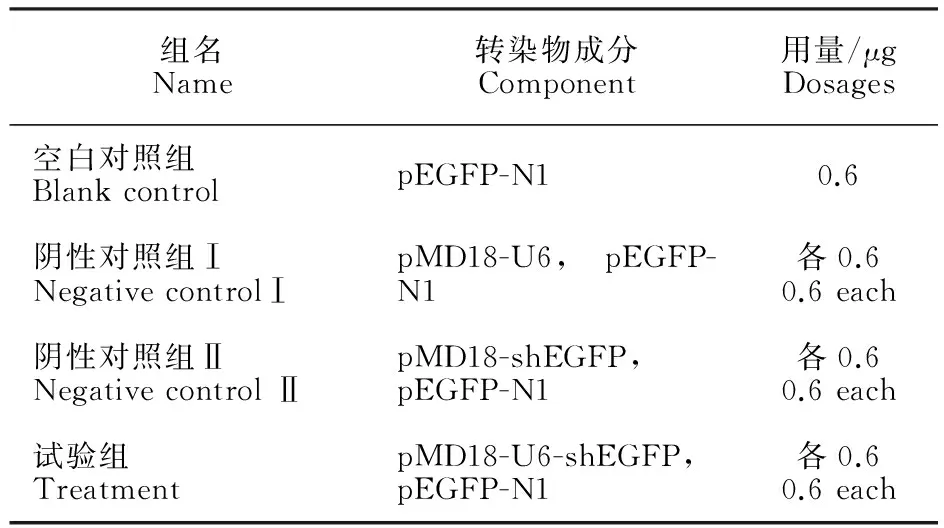
表1 轉染組及轉染成分 Table 1 Transfected group and compositions
2 結果與分析
2.1 U6 snRNA基因上游5′-端側翼基因克隆與序列分析
采用染色體步移法獲得東方粘蟲 U6 snRNA基因上游5′-端側翼序列并測序,該序列長1 426 bp。經NCBI數據庫序列比對發現,該序列下游3′-端與家蠶(Bombyxmori) U6 snRNA基因序列(AY649381)相似度為100%,與黑腹果蠅(Drosophilamelanogaster) U6 snRNA基因序列(M24605)相似度為96%,與豹紋蝶(Melitaeacinxia) U6 snRNA基因序列(JX878560.1)相似度為96%。由此推斷這些相似序列為東方粘蟲 U6 snRNA基因序列。經啟動子在線分析軟件NNPP v. 2. 2 (http://www.fruitfly.org/ seq_tools/ promoter.html) 預測,東方粘蟲 U6 snRNA 基因上游5′-端側翼序列有4個可能的轉錄起始位點(圖1),其轉錄起始位點上游存在著 TATA box,符合真核生物基因轉錄起始位點所具有的特征。
參照Domitrovich等[25]的方法尋找東方粘蟲 U6啟動子元件,結果發現,該序列中包含RNA聚合酶Ⅲ啟動子的典型基序,包括遠端序列元件DSE,近端序列元件PSE及TATA box,DSE由1個能夠與轉錄因子Oct-1相結合的八聚體序列(OCT),和1個與刺激反式作用因子(stimulated trans-acting factor, STAF)相結合的SPH元件組成(圖2)。
2.2 東方粘蟲 U6啟動子的克隆
根據東方粘蟲 U6 snRNA基因上游5′-端側翼序列,設計兩對引物F1/R1、F2/R2,以基因組為模板,得到分別含BamHⅠ、Hind Ⅲ與含EcoRⅠ、BamHⅠ的 U6啟動子(圖3)。

加粗字母代表可能的轉錄起始位點 The bold letters mean prediction of transcription start site

OCT, SPH, PSE, TATA-box都用下劃線標出,預測的轉錄起始位點作為+1,并被框出 OCT, SPH, PSE, TATA box were underlined, and start site of the predicted transcription were recorded as +1, and was out of the box.
圖2 東方粘蟲 U6 snRNA基因5′-端側翼序列分析
Fig.2 Analysis of 5′-flanking region of U6 snRNA gene inM.separata

M. Marker DL 5000; 1. 含BamHⅠ、Hind Ⅲ的 U6啟動子 U6 promoter withBamHⅠ andHind Ⅲ; 2. 含EcoRⅠ、BamHⅠ的 U6啟動子 U6 promoter withEcoRⅠ andBamHⅠ
圖3 具有酶切位點的東方粘蟲 U6啟動子
Fig.3 U6 promoters ofM.separatawith restriction sites
2.3 基因沉默載體的構建與雙酶切驗證
為了構建基因沉默載體,首先需要對pMD18-T載體進行環化,為此設計并克隆一段515 bp的片段N,該片段不包含構建載體所需要的3個酶切位點:EcoRⅠ、BamHⅠ及Hind Ⅲ。經T-A克隆后,雙酶切驗證N片段成功導入pMD18-T載體中(圖4)。隨后分別將 U6、shEGFP及U6-shEGFP片段重組到pMD18-N,構建載體pMD18-U6、pMD18-shEGFP及pMD18-U6-shEGFP,雙酶切結果表明目的片段成功轉入載體(圖5)。2.4 RNAi載體轉染293 T細胞的熒光顯微鏡檢測
將載體pMD18-U6、pMD18-shEGFP與pMD18-U6-shEGFP分別與pEGFP-N1共轉染293 T細胞。將各孔細胞置于倒置熒光顯微鏡下,觀察同一視野下細胞生長情況及各組熒光強度。結果如圖6所示:在轉染24 h、48 h與72 h后,與pEGFP-N1空白對照、pMD18-U6陰性對照和pMD18-shEGFP陰性對照相比,pMD18-U6-shEGFP的熒光強度均顯著降低,表明shEGFP可以在293 T細胞中成功表達并引起熒光蛋白的表達量降低。
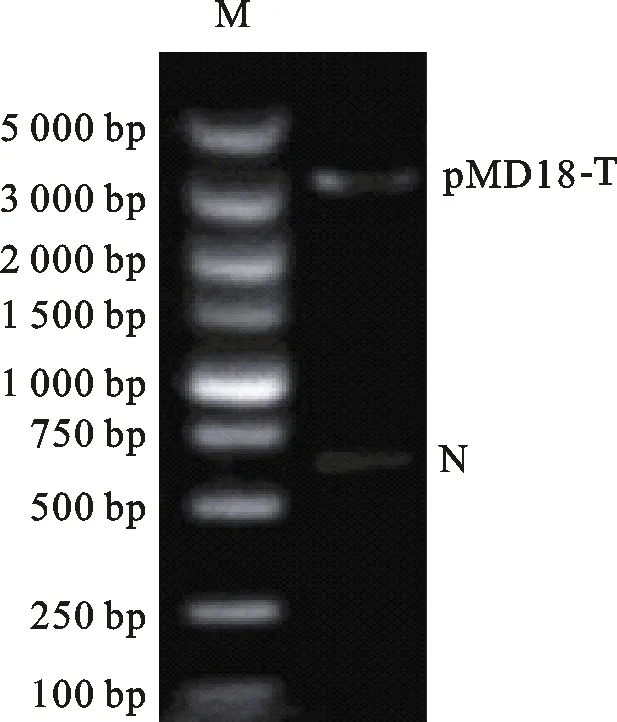
M. Marker DL 5000
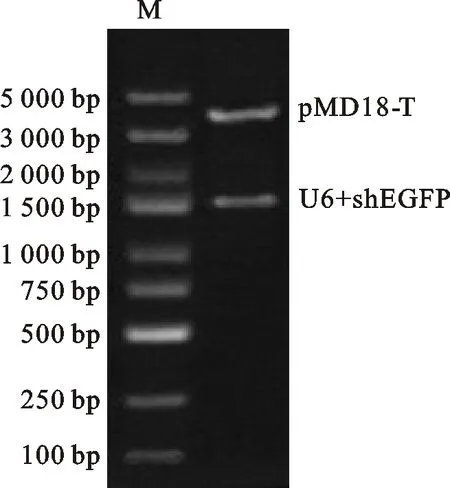
M. Marker DL 5000
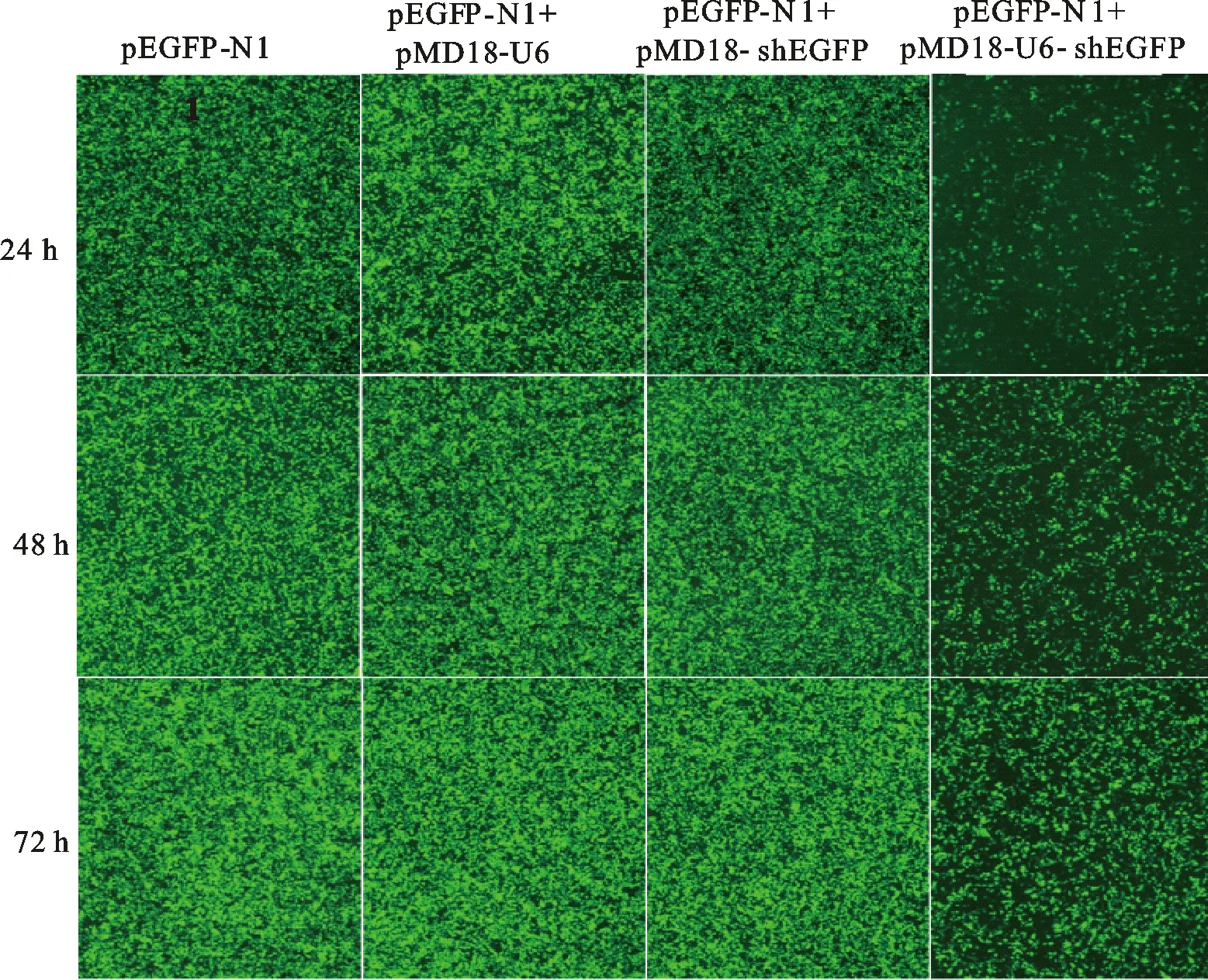
圖6 轉染后不同時間293 T細胞熒光Fig.6 293 T cell fluorescence after transfected by different vectors with pEGFP-N1
3 討 論
U6啟動子的3′-端序列一般較為保守,但5′-端側翼序列及其長度往往差異較大,用常規的RACE等技術很難克隆到啟動子的全長序列。本研究采用染色體步移技術,首先分別用4個限制性內切酶(DraⅠ、EcoRⅤ、PvuⅡ和StuⅠ)對東方粘蟲基因組DNA進行酶切,將其分成4個帶相應酶切位點的DNA文庫,添加特異性接頭后設計上游引物,再參照近源物種家蠶和果蠅 U6 snRNA的3′-端共有序列設計下游特異性引物,用染色體步移試劑盒獲得5′-端側翼片段并逐級向前推動,最終克隆出東方粘蟲 U6啟動子序列。
對克隆的 U6啟動子序列分析表明,該序列包含OCT,SPH, DSE, PSE及TATA-box等RNA pol Ⅲ啟動子特有的序列元件,與有關文獻[25-27]報道的人類、雞及水牛等動物的 U6啟動子特征元件一致。經與NCBI數據庫比對發現,該 U6啟動子3′-端序列與家蠶及果蠅 U6 snRNA基因部分序列均有很高的同源性(分別為100%和96%)。因此,確定此序列為東方粘蟲 U6 snRNA基因,其上游5′-端側翼序列包含東方粘蟲的 U6啟動子。
U6啟動子能否有效驅動RNA的表達是檢驗 U6 snRNA是否包含整個啟動子序列的關鍵。為了驗證東方粘蟲 U6啟動子序列是否完整,本研究用siDicer軟件對pEGFP-N1基因序列進行siRNA預測分析,隨機挑選1個siRNA進行shRNA合成,并將其構建到由東方粘蟲 U6啟動子驅動的shRNA載體中。細胞轉染證實,在24、48、72 h時,試驗組熒光與空白對照組和陰性對照組均有顯著差異,而空白對照組和各陰性對照組之間熒光則無明顯差異。由此表明,本研究克隆獲得的東方粘蟲 U6 snRNA包含完整的 U6啟動子序列,能成功驅動shRNA的表達并可用于后續基因沉默載體的構建。
Reference:
[1] FIRE A,XU S Q,MONTGOMERY M K,etal.Potent and specific genetic interference by double-stranded RNA inCaenorhabditiselegans[J].Nature,1998,391(6669):806-811.
[2] HARBORTH J,ELBASHIR S M,BECHERT K,etal.Identification of essential gene in cultured mammalian cells using small interfering RNAs [J].JournalofCellScience,2001,114(24):4557-4565.
[3] AZA-BLANC P,COOPER C L,WAGNER K,etal.Identification of modulator of TRAIL-induced apoptosis via RNAi-based phenotypic screening [J].MolecularCell,2003,12(3):797-801.
[4] GE Q,MCMANUS M T,NGUYEN T,etal.RNA interference of influenza virus production by directly targeting mRNA for degradation and indirectly inhibiting all viral RNA transcription [J].ProceedingsoftheNationalAcademyofSciencesoftheUSA,2003,100(5):2718-2723.
[5] LE H,KUMAR V.Specific inhibition of gene expression aid trans-activation functions of hepatitis B virus X protein and c-myc by small interfering RNAs [J].FEBSLetters,2004,560(1-3):210-214.
[6] SHIRANE D,SUGAO K,NAMIKI S,etal.Enzymatic production of RNAi libraries from cDNAs [J].NatureGenetics,2004,36(2):190-196.
[7] DYKXHOORN D M,LIEBERMAN J.Running interference:prospects and obstacles to using small interfering RNAs as small molecule drugs [J].AnnualReviewofBiomedicalEngineering,2006,8:377-402.
[8] TIJSTERMAN M,PLASTERK R H A.Dicers at RISC; the mechanism of RNAi [J].Cell,2004,117(1):1-3.
[9] HU S W,NI W,HAZI W,etal.Cloning and functional analysis of sheep U6 promoters[J].AnimalBiotechnology,2011,22(3):170-174.
[10] 楚莉輝,劉龍丁,馬紹輝,等.利用人 U6 snRNA啟動子構建RNA干擾質粒載體[J].中國生物化學與分子生物學報,2006,22(3):228-233.
CHU L H,LIU L D,MA SH H,etal.Constructing of an RNAi plasmid based on human U6 snRNA[J].ChineseJournalofBiochemistryMolecularBiology,2006,22(3):228-233(in Chinese with English abstract).
[11] MIYAGISHI M,TAIRA K. U6 promoter-driven siRNAs with four uridine 3'overhangs efficiently suppress targeted gene expression in mammalian cells [J].NatureBiotechnology,2002,20(5):497-500.
[12] ZHOU H,HUANG C,XIA X G.A tightly regulated Pol Ⅲ promoter for synthesis of miRNA genes in tandem [J].BiochimicaetBiophysicaActa,2008,1779(11):773-779.
[13] ELBASHIR S M,HARBORTH J,WEBER K,etal.Analysis of gene function in somatic mammalian cells using small interfering RNAs [J].Methods,2002,26(2):199-213.
[14] OHTSUKA D,NAKATSUKASA T,FUJITA R,etal.Use of bombyx mori U6 promoter for inducing gene-silencing in silkworm cells [J].JournalofInsectBiotechnologyandSericology,2008,77(3):125-131.
[15] KIM M S,JEE B Y,CHO M Y,etal.Fugu double U6 promoter-driven long double-stranded RNA inhibits proliferation of viral hemorrhagic septicemia virus (VHSV) in fish cell lines [J].ArchivesofVirology,2012,157(6):1029-1038.
[16] SU J G,ZHU Z Y,XIONG F,etal.Hybrid cytomegalovirus- U6 promoter-based plasmid vectors improve efficiency of RNA interference in zebrafish [J].MarineBiotechnology,2008,10(5):511-517.
[17] CARBON P,MURGO S,EBEL J P,etal.A common octamer motif binding protein is involved in the transcription of U6 snRNA by RNA polymerase Ⅲ and U2 snRNA by RNA polymeraseⅡ[J].Cell,1987,51(1):71-79.
[18] LAMBETH L S,WISE T G,MOORE R J,etal.Comparison of bovine RNA polymerase Ⅲ promoters for short hairpin RNA expression [J].AnimalGenetics,2006,37(4):369-372.
[19] WISE T G,SCHAFER D J,LAMBETH L S,etal.Characterization and comparison of chicken U6 promoters for the expression of short hairpin RNAs [J].AnimalBiotechnology,2007,18(3):153-162.
[20] RENJU T,GIRADDI R,HUNJE R,etal.Evaluation of new insecticidal poison baits againstMythimnaseparata(Walker) in sorghum[J].KarnatakaJournalofAgriculturalSciences,2009,22(4):7773-776.
[21] GU L Q,KNIPPLE D C.Recent advance in RNA interference research in insects:Implications for future insect pest management strategies [J].CropPrtection,2013,45:36-40.
[22] BAUM J A,BOGAERT T,CLINTON W,etal.Control of coleopteran insect pests through RNA interference [J].NatureBiotechnology,2007,25(11):1322-1326.
[23] THAKUR N,UPADHYAY S K,VERMA P C,etal.Enhanced whitefly resistance in transgenic tobacco plants expressing double stranded RNA of V-ATPaseAgene [J].PloSone,2014,9(3):e87235.
[24] 張淑靜,王高振,劉 爽,等.利用RNAi技術沉默東方粘蟲V-ATP酶H亞基研究[J].西北農業學報,2015,24(1):170-174.
ZHANG SH J,WANG G ZH,LIU SH,etal.Silencing of V-ATPase subunit H ofMythinmaseparatathrough RNA interference [J].ActaAgriculturalBoreali-occidentalisSinica,2015,24(1):170-174(in Chinese with English abstract).
[25] DOMITROVICH A M,KUNKEL G R.Multiple,dispersed human U6 small nuclear RNA genes with varied transcriptional efficiencies [J].NucleicAcidsResearch,2003,31(9):2344-2352.
[26] 陳新雨.雞RNA聚合酶Ⅲ型啟動子7SK克隆和轉錄shRNA活性研究[D].陜西楊陵:西北農林科技大學,2012:21-23.
CHEN X Y.Cloning of chicken RNA polymerase Ⅲ type Ⅲ promoter 7SK and characterization of its shRNA transcription activity in vitro[D].Yangling Shaanxi:Northwest A&F University,2012,21-23(in Chinese with English abstract).
[27] 張曉溪,劉慶友,鄧彥飛,等.水牛RNA聚合酶Ⅲ啟動子的克隆與鑒定[J].南方農業學報,2014,45(5):858-863.
ZHANG X X,LIU Q Y,DENG Y F,etal.Cloning and identification of bufalo RNA polymerase Ⅲ promoters [J].JournalofSouthernAgriculture,2014,45(5):858-863(in Chinese with English abstract).
(責任編輯:郭柏壽 Responsible editor:GUO Baishou)
Cloning and Functional Verification of U6 Promoter fromMythimnaseparata(Walker)
WANG Gaozhen, JIN Duo, WU Wenjun and QI Zhijun
(Institute of Pesticide Science, College of Plant Protection, Northwest A&F University, Yangling Shaanxi 712100,China)
In order to obtain constantly short interference RNA(siRNA) , the 1 426 bp 5′- flanking promoter sequence of U6 snRNA gene ofMythimnaseparata(Walker) was cloned by the method of Genome Walking, as well as its function was verified in this study. Based on bioinformatics analysis, the results showed that the promoter fragment contained polymerase Ⅲ core promoter elements SPH, OCT, DSE, PSE and TATA box. The observation of expression level enhanced green fluorescent protein EGFP after being transfected with RNA interference (RNA interference, RNAi) , the vectors by fluorescence microscope showed that the U6 promoter fragment drived the expression of shEGFP successfully. This study had established a foundation for further construction of RNAi vectors which can silence V-ATPase subunit H.
Mythimnaseparate(Walker); RNAi; U6 promoter; Functional verification
2016-05-19 Returned 2016-06-28
The National Natural Science Foundation of China (No. 31371958).
WANG Gaozhen, male, master student. Research area:pesticide toxicology. E-mail:wanggaozhen@nwsuaf.edu.cn
QI Zhijun, male, associate professor. Research area:pesticide molecular toxicology. E-mail:qzhij@nwsuaf.edu.cn
日期:2017-06-05
2016-05-19
2016-06-28
國家自然科學基金(31371958)。 第一作者:王高振,男,碩士研究生,從事農藥毒理學研究。E-mail:wanggaozhen@nwsuaf.edu.cn 通信作者:祁志軍,男,副教授,主要從事農藥分子毒理學研究。E-mail:qzhij@nwsuaf.edu.cn
Q789
A
1004-1389(2017)06-0939-07
網絡出版地址:http://kns.cnki.net/kcms/detail/61.1220.S.20170605.1728.036.html
