金黃色葡萄球菌中的群感效應研究
2017-07-31 16:19:42鐘玉心關潔婷王偉
農產品加工 2017年11期
鐘玉心,洪,關潔婷,王偉
(1.廣州市食品檢驗所,廣東廣州510000;2.東莞出入境檢驗檢疫局,廣東東莞523000;3.廣州市怡臻原文化傳播有限公司,廣東廣州510000)
金黃色葡萄球菌中的群感效應研究
鐘玉心1,洪2,關潔婷3,王偉2
(1.廣州市食品檢驗所,廣東廣州510000;2.東莞出入境檢驗檢疫局,廣東東莞523000;3.廣州市怡臻原文化傳播有限公司,廣東廣州510000)
金黃色葡萄球菌是一種能夠引起繼發性感染的典型機會致病菌,可以引起局部化膿感染,也可以引發肺炎、心內膜炎甚至敗血癥等全身感染。而群感效應在金黃色葡萄球菌所引起的疾病和毒素表達中都起著關鍵性作用。群感效應是一種細胞間的信息交流過程,它可以使得細菌之間相互傳遞關于外界環境和種群數量的信息,進而相應地調整基因表達,以確保自身能夠最大程度地適應外界環境。介紹金黃色葡萄球菌中的群感效應系統,并對其4種基因元件——AgrBDCA及其作用進行了闡述。
金色葡萄球菌;群感效應;基因表達
0 引言
群感效應(Quorum sensing)是細菌為了應對種群數量變化或是外界環境限制的一種調節相關基因表達的作用機制[1-5]。1979年,人們首次對費氏弧菌(Vibrio fischeri)和哈氏弧菌(Vibrio harveyi)中的群感效應進行了報道[1,3,6]。費氏弧菌寄生夏威夷短尾魷魚(Euprymna scolopes)的發光器官中,當細菌的數量達到一定程度會誘導與生物發光相關基因的轉錄,從而發光。夏威夷短尾魷魚利用所發出的光進行交流、捕食、躲避天敵[6-7]。正是由于群感效應的存在,細菌可以在整個菌群的范圍內調整基因表達,使整個菌群的行為協調一致,這使得細菌不再是簡單相互獨立的個體,更像是一個多細胞的生命體[5,6,8]。
細菌會產生一種小分子的化學物質作為群感效應中的信號[9]。在相對封閉的環境中,這種信號分子會隨著細菌數量的增多而增多,當信號分子的濃度達到一定閾值時,將會對細菌的基因表達產生一些調節作用,這種化學信號稱之為自誘導物(Autoinducer,AIs)。有些自誘導物能夠促進自身的生成,而有些則不能[2-3,5,9]。一般來說,革蘭氏陰性菌的自誘導物是N-酰基高絲氨酸內酯(Nacyl-homoserine lactones,HSLs),而革蘭氏陽性菌是一種小分子短肽(Autoinducer peptides,AIPs)[2-3,9,10],這2類自誘導物的共同點就是在構型上二者都具有內酯環。但是HSLs以自由擴散的方式自由地出入細胞,當HSLs濃度達到閾值后,會與存在于細胞質中特殊的受體蛋白結合,活化后的受體蛋白可以作為一種轉錄激活因子調控基因的表達[2,9]。而AIPs并不能直接進入細胞質,它需要與位于細胞膜上的受體蛋白結合,通過受體蛋白來與其他蛋白的作用用調控基因表達[2]。
1 金黃色葡萄球菌中的群感效應通路
對于葡萄球菌這類機會致病菌來說經常會分泌一些胞外物質,這些胞外物質能夠有效地保護細菌免受宿主免疫系統的侵襲,使細菌牢固地黏附在宿主細胞表面,而且在一定程度上能夠降解宿主組織細胞,為細菌提供營養[9]。這些胞外物質的分泌與基因表達高度協調,并且與細菌的生理需要密切相關。對于金黃色葡萄球菌來說,群感效應系統——Agr系統作為核心調節元件之一,可以對胞外物質與毒素的分泌進行有效調控。也有研究發現,有一些金黃色葡萄球菌會利用LuxS的產物AI-2(Autoinducer-2),一種呋喃酰硼酸二酯來進行種間的信息傳遞。
Agr系統所編碼出的蛋白構成了金黃色葡萄球菌中的群感效應通路,其中包括了2個不同的啟動子,即P2與P3。P2的轉錄本包含了AgrD,AgrB,AgrC與AgrA 4個基因,P3的轉錄本為RNAIII。AgrC與AgrA共同構成了經典的二元應答調節系統,而AgrD與AgrB負責AIP的產生與加工。成熟的AIP會被分泌到細胞外,當AIPs的濃度達到一定閾值會活化位于細胞膜上的AgrC受體。細胞質內的AgrA被AgrC激活后,會與P2,P3啟動子結合調節基因表達。RNAIII具有雙重作用,一方面可以促進毒素基因的表達,另一方面抑制了表面蛋白相關基因的表達。在革蘭氏陽性菌中,大多數都具備這一群感效應通路。
在體外培養中,金黃色葡萄球菌分泌胞外蛋白可以分為2個階段。初期主要是負責黏附的蛋白,如蛋白A(ProteinA)、凝固酶、纖維蛋白結合蛋白(Fibronection binding proteins),后期主要是一些蛋白酶、降解酶、溶血素和毒素。研究結果也顯示,在細菌生長的指數后期,負責編碼表面蛋白的基因表達量有所下降,而負責編碼毒素蛋白和降解蛋白的基因表達量有所上升。在這不同階段的轉變中,Agr系統起著重要的作用。
金黃色葡萄球菌中的Agr系統見圖1。
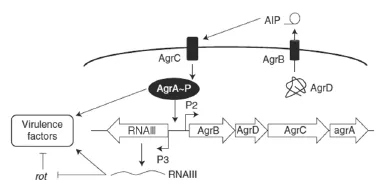
圖1 金黃色葡萄球菌中的Agr系統
2 Agr介導群感效應通路的組成元件
2.1 AgrB
在葡萄球菌中,AgrB的N端1/3和AgrC的C端組氨酸激酶域是保守區域,其他則是可變區,這使得在Agr系統在金黃色葡萄球菌中可以分為4類。每一類Agr系統都會產生特異的AIPs信號和與之相對應的AgrC受體。AIPs會特異性地活化與之同源的AgrC受體,抑制異源的AgrC,即使AIP上1個氨基酸的改變都會影響這種特異性。
葡萄球菌中的4類AIP見圖2。
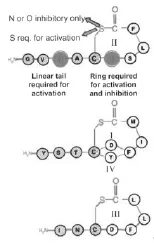
圖2 葡萄球菌中的4類AIP
2.2 AgrD
AgrD負責編碼出AIPs的前體,這種前體AIPs會在相關酶的作用下在C段和N段接受加工,在半胱氨酸和C端形成1個硫代內酯環。所形成的硫代內酯環對AIPs至關重要,而且是AIPs的特征之一。AgrB是位于細胞膜上的肽鏈內切酶,對于AIP的合成加工具有重要作用。AgrB可以特異性地識別AgrD的C端序列,并催化硫代內酯環的形成。經研究發現,這種作用與AgrB上保守組氨酸和半胱氨酸殘基有關。
AgrB識別AgrD的特異性略低于AgrC識別AIP的特異性,所以不同類別的AgrB與AgrD可以相互作用。研究發現,AgrB-I可以以相同的速率對A-grD-I與AgrD-III進行加工。對比金黃色葡萄球菌的4種AgrD序列可以發現,AIPs由7~9個氨基酸殘基所組成,其中包含了1個保守的半胱氨酸,在C末端有2個疏水氨基,由半胱酸與C末端的疏水氨基酸形成五元硫代內酯環。
在金黃色葡萄球菌中4類AgrD的序列見表1。
4種AIPs的結構如圖2所示,結構中的硫代內酯環是整個AIPs活性的中心,其所用涉及整個Agr系統的激活與抑制。如果把AIPs中的硫代內酯環用內酯環所取代,那么其激活同源AgrC受體的功能將會消失,但仍具有抑制異源AgrC的功能。當然AIPs的N端尾部結構也能夠影響其功能,將AIPsII N端第3位的丙氨酸用天冬氨酸代替之后,它會抑制所有的4類AgrCs受體。由此可見,AIPs的功能與其結構密切相關。
2.3 AgrC
AgrC作為Agr系統中的受體屬于組氨酸蛋白激酶的一種,它有2個部分構成:N端的跨膜感受域(Transmeme brane sensor domain)和C端的組氨酸蛋白激酶域(Histidine protein kinase domain)。AgrC的組氨酸蛋白激酶域其氨基酸序列高度保守,但是跨膜感受域涉及AIPs的特異性識別,其序列是可變的。
AgrCI的氨基酸序列見圖3。

表1 在金黃色葡萄球菌中4類AgrD的序列
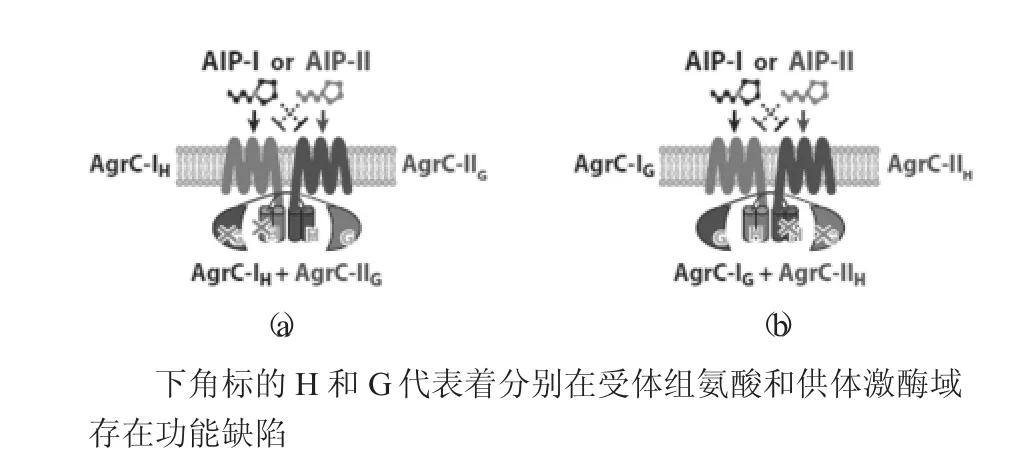
圖4 AgrC的二聚體結構
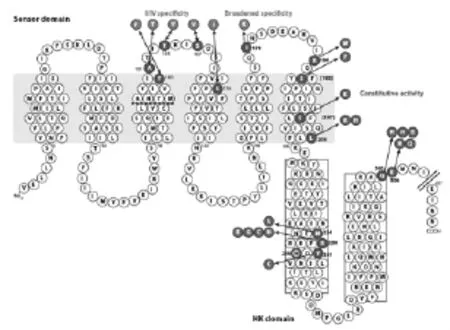
圖3 AgrC I的氨基酸序列
有人將4類AgrC的近端感受域與遠N端感受域進行了互換,以研究其具體功能。結果顯示,AIPs的特異性識別主要涉及遠端感受域,非特異性的連接和信號傳遞涉及跨膜螺旋3,4,6和膜外第3個環結構。
George等人認為,AgrC受體是以二聚體的方式在細胞膜上聚集的。將在受體組氨酸(Receptorhistindine)或是供體激酶結構域(Donorkinasedomain)存在變異的AgrC I和AgrC II導入不存在AgrC基因的細菌中,會發現本應該沒有活性的Agr系統也出現了一定的活性。相關研究認為AgrC是以2聚體的方式在一起,至使不同受體的激酶域卷曲螺旋緊密相連,組成4-螺旋束結構域(4-helixbundle)。這樣當活化后的感受域影響4-螺旋束結構域時,才會進一步活化2個受體的激酶結構域[9]。
AgrC的二聚體結構見圖4。
2.4 AgrA
AgrA是一種反應調節蛋白(Response regulator),是LytTR反應調節蛋白家族中的一員,可以與Agr系統的2個啟動子結合,促進基因的轉錄。Robbin等人的研究表明,P2和P3啟動子在下游重復序列存在著2 bp的不同。這種不同導致了AgrA與2個啟動子的親和力不同,AgrA更容易與P2結合。SarA也有著可以和P2啟動子結和,促進基因轉錄的功能,在一些特定的條件下AgrA可以和SarA共同作用,促進基因轉錄。
2.5 RNAIII
RNAIII是位于細胞內Agr系統的效應子(Effector),大小為514個核苷酸,其半衰期在45 min以上。RNAIII的序列在不同種微生物中表現不同,但是它的結構類似,這使得種間的信息交流成為可能。
關于RNAIII的功能研究揭示了其3'端對于抑制蛋白A(spa編碼)的表達十分必要,沒有重疊的5'和3'區域可以獨立地促進α-溶血素(hla編碼)的表達。也就是說,RNAIII可以幫助實現細菌從表達表面蛋白到表達毒素的轉變。在第7,13和14個莖環結構中,那些富含胞嘧啶的環結構與典型的SD區(Shine-Dalgarnosequecnce)互補,這也意味著RNAIII的基本功能使作為反義RNA參與胞外毒素的轉錄后調控。RNAIII與hlamRNA的SD區結合后,可以改變hlamRNA的二級結構,使其更易于翻譯。而RNAIII和蛋白A及纖維素結合蛋白(Fibrinogen-binding protein)的轉錄本起始位點結合,阻礙翻譯的發生。同時研究表明,RNAIII也可以通過類似的機制來阻礙另外一個轉錄影響因子Rot的合成。Rot的功能和RNAIII的正好相反,Rot可以促進表面蛋白的產生,抑制分泌蛋白的產生[2]。
RNAIII的二級結構見圖5。
3 Agr系統的作用
生物被膜(Biofilm)的形成是細菌體內極其復雜的一個過程,同時它也受到自身基因的調控和試驗條件的影響。被膜的形成可以分為2種不同的機制:①以PIA為基本物質的被膜;②以黏附蛋白為基本物質的被膜[2]。
有研究表明,Agr缺陷性的金黃色葡萄球菌和Agr缺陷型的表面葡萄球菌能比攜帶Agr的菌株形成更加厚重的被膜,這也許是和Agr系統可以上調δ溶血素的表達,下調黏附蛋白的表達[2]。黏附蛋白的表達在被膜形成的初期起著關鍵性作用,而δ溶血素則能夠裂解成熟的生物被膜。有研究表明,Agr系統的主要作用是通過上調蛋白酶產生引起生物被膜的裂解,蛋白對保持生物被膜的完整性具有重要作用。
總的來說,QS對于金黃色葡萄球菌生物被膜形成的動態過程具有重要作用,但是簡單地認為QS是可以促進被膜生成或是可以抑制被膜生成又太過武斷。有研究表明,RIP(YSPWTNF)一種七肽,可以干擾金黃色葡萄球菌生物被膜的形成,而且這種干擾是通過抑制Agr系統來實現的。而這正與之前的Agr系統可以抑制生物被膜的研究結果相矛盾,所以RIP干擾生物被膜的具體機制需要進一步探究。
在動物感染試驗中,Agr也是一種重要的影響因子。在皮膚膿腫(Skin abscesses)、心內膜炎(Endocarditis)等動物模型中,發現Agr突變有所減弱。而且醫學數據也顯示出,Agr系統對于金黃色葡萄球菌所引起的感染具有重要作用。在由葡萄球菌所引起的肺炎中,20%分離出的菌種不攜帶Agr。雖然Agr缺陷型菌株也有治病能力,但是所引起疾病的嚴重程度明顯低于具有Agr的菌株[9]。
4 結論
群感效應對于細菌調節機體行為至關重要,同時也可以使其探索新的適宜生存環境。基于自誘導物的產生和檢測,QS系統可以在整個菌群范圍內調整基因的表達。而其他調節機制正是在QS的整體調節基礎上,確保調節基因表達都能準確應對外界環境的變化。
同時,QS系統也和生物被膜的形成相關。在生物被膜包被下的細菌,都能夠從這種彼此的信息傳遞中受益。在QS中所涉及到的自誘導物小分子是一種核心調節子,現在這種生物小分子調節細菌行為的作用已漸漸被人們賦予了新的意義。人們可以通過調控這些生物小分子來調節細菌行為,進而控制疾病,而且這種新的治療方法其目的是減少病原菌毒素的表達,而不是抑制細菌的生長。所以,通過干擾生物小分子調節通路的治療方法是在抗生素治療方法之后的新型方法。
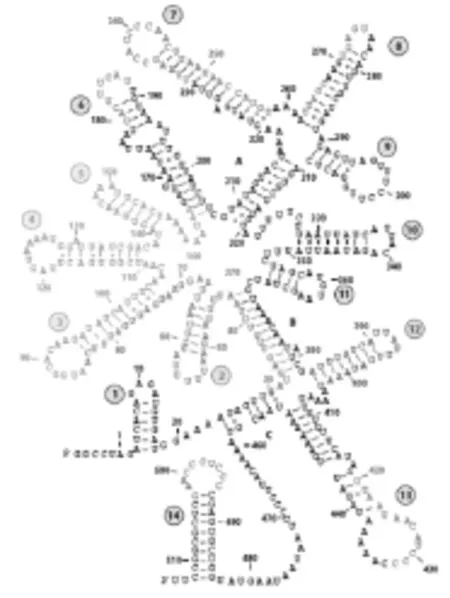
圖5 RNAIII的二級結構
[1]Melissa B Miller,Bonnie L Bassler.Quorum sensing in bacteria[J].Annual Review of Microbiology,2001(5):165-199.
[2]Steben T Rutherford,Bonnie L Bassler.Bacterial quorum sensing:its role in virulence and possibilities for its control[J].Cold Apring Harbor Perspectives in Medicine,2012,2(11):705-709.
[3]Antunes L C,Ferreira R B,Buckner M M,et al.Quorum sensing in bacterial virulence[J].Micobiology,2010(7):2 271-2 282.
[4]Mattgew D S,Jessica L Terrell,Chen-Yu Tsao,et al.Directed assembly of a bacterial quorum[J].ISME Journal,2016(10):158-169.
[5]Laurent Keller,Michael G Surette.Communication in bacteria:an ecological and evolutionary persepectvie[J]. Nature Reviews Microbiology,2006,4(4):249-258.
[6]Chrostopher M Waters,Bonnie L Bassler.Quorum sensing:cell-to-cell communication in bacteria[J].The Annual Review of Cell and Developmental Biology,2005(2):319-346.
[7]Tom Defoirdt,Nico Boom,Patrick Sorgeloos,et al.Quorun sensing and quorum quenching in Vibrio harveyi:lessons learned from in vivo work[J].ISME Journal,2008(2):19-26.
[8]Li Yung Hua,Tian Xiaolin.Quorum sensing and bacterial social interactions in biofilms[J].Sensors,2012,12(3):2 519-2 538.
[9]Richard P Novick,Edward Geisinger.Quorum sensing in staphylococci[J].Annual Review of Genetics,2008,42(1):541-564.
[10]Burkhard A Hense,Christina Kutttler,Johannes Muller,et al.Dose efficiency sensing unify diffusion and quorum sensing[J].Nature Reviews Microbiology,2007,5(3):230-239.◇
Research on the Quorum Sensing of Staphylococcus aureus
ZHONG Yuxin1,HONG Yuan2,GUANG Jieting3,WANG Wei2
(1.Guangzhou Institute for Food Control,Guangzhou,Guangdong 510000,China;2.Dongguan Entry-exit Inspection and Quarantine Bureau,Dongguan,Guangdong 523000,China;3.Guangzhou Yizhenyuan Cultural Diffusion Co.,Ltd.,Guangzhou,Guangdong 510000,China)
Staphylococcus aureus is a kind of typical opportunistic pathogens that can cause secondary infection,it can cause local purulent infection,also can cause pneumonia,endocarditis,and even sepsis systemic infection.The quorum sensing in diseases caused by Staphylococcus aureus and toxin expression plays a key role.Quorum sensing is a kind of communication process between the cells,it can make bacteria pass each other information about the external environment and population,and then adjust gene expression,to ensure that itself adapt to the environment to a great extent.The quorum sensing of Staphylococcus aureus is reviewed in this paper,and the four kinds of genetic elements——AgrBDCA and their effect are expounded.
Staphylococcus aureus;quorum sensing;gene expression
R392
A
10.16693/j.cnki.1671-9646(X).2017.06.018
1671-9646(2017)06a-0055-04
2017-04-15
鐘玉心(1986—),女,本科,助理工程師,研究方向為食品添加劑的檢驗。
猜你喜歡
工業設計(2022年8期)2022-09-09 07:43:20
核科學與工程(2021年4期)2022-01-12 06:30:26
軍民兩用技術與產品(2021年10期)2021-03-16 06:05:30
北京測繪(2020年12期)2020-12-29 01:33:58
今日農業(2020年19期)2020-12-14 14:16:52
小學生必讀(中年級版)(2020年9期)2020-12-04 02:07:22
裝備制造技術(2019年12期)2019-12-25 03:06:46
中國洗滌用品工業(2019年4期)2019-05-11 09:27:34
家庭影院技術(2017年9期)2017-09-26 03:41:45
中學物理·高中(2016年12期)2017-04-22 11:53:03
