鯛無胄蟲的描述及無胄蟲科的分類地位
2017-08-16 06:06:57周梓華丁雪娟
水生生物學報 2017年4期
關鍵詞:分類
周梓華 蔡 星 丁雪娟
(華南師范大學生命科學學院, 廣東省水產健康安全養殖重點實驗室, 廣州市亞熱帶生物多樣性與環境生物監測重點實驗室,廣州 510631)
鯛無胄蟲的描述及無胄蟲科的分類地位
周梓華 蔡 星 丁雪娟
(華南師范大學生命科學學院, 廣東省水產健康安全養殖重點實驗室, 廣州市亞熱帶生物多樣性與環境生物監測重點實驗室,廣州 510631)
對寄生于大亞灣黑鯛(Acanthopagrus schlegeli)體表的鯛無胄蟲(Anoplodiscus spari)進行了形態描述, 并擴增了該蟲的28S rDNA C1-D2區部分序列, 構建了相關類群的分子系統樹, 探討了無胄蟲科的分類地位。結果顯示: 所獲標本與Ogawa等的描述基本一致, 但交接管基部及末端位置略有不同: 本研究標本的交接管基部始于支持器的下方、末端于支持器三分叉的中支略伸出, 而Ogawa等的標本交接管基部靠近支持器三叉處、末端從三叉處延伸并略彎曲; 無胄蟲屬(Anoplodiscus)為單系起源, 隸屬于指環蟲目、四鉤蟲亞目、無胄蟲科(Anoplodiscidae), 無胄蟲科是一獨立的科, 在分類地位上與四鉤蟲科(Tetraonchidae)互為姐妹群, 與Udonellidae科和微溝科(Microbothriidae)的關系較遠。
鯛無胄蟲; 無胄蟲科; 28S rDNA; 分類地位
無胄蟲科(Anoplodiscidae Tagliani, 1912)隸屬于扁形動物門(Platyhelminthes)單殖吸蟲綱(Monogenea), 目前僅含無胄蟲屬(Anoplodiscus)。無胄蟲屬是由Sonsino (1890)以采自真鯛(Pagrus major)上的Anoplodiscus richiardii為模式種而建立的, 屬征為: 后吸器盤狀, 無幾丁質結構; 腸支單管, 具不對稱側分支; 前吸器1對, 吸槽狀[1]。無胄蟲屬記錄6個種: 寄生于赤鯛(Pagrus pagrus)的A.richiardii Sonsino, 1890和A.longivaginatus Paraguassu, 2002、寄生于黑鯛(Acanthopagrus schlegeli)的A.australis (Johnston, 1930) Ogawa et Egusa, 1981和A.spari (Yamaguti, 1958) Ogawa et Egusa, 1981、寄生于金鯛(Chrysophrys auratus)的A.cirrusspiralis Roubal, Armitage et Rohde, 1983及寄生于真鯛的A.tai Ogawa, 1994[1—5]。張劍英等[6]曾對無胄蟲科無胄蟲屬作過介紹, 吳金英等[7]記述了采自香港海域黑鯛體表的鯛無胄蟲(A.spari), 此為我國報道的唯一一種無胄蟲[7,8], 但未提供該蟲的相關圖片。近年又有該蟲引發養殖魚類病害的報道[9]。
無胄蟲的分類地位一直存在爭議。Monticelli (1905)重描述了無胄蟲屬并將其歸在Anisocotylidae科。Tagliani (1912)在該屬的基礎上建立了無胄蟲亞科Anoplodiscinae。Price (1938)認為Anisocotylidae科名無效, 并將無胄蟲屬歸入微溝蟲科 (Microbothriidae)。Sproston (1946)又將無胄蟲屬歸于鞋口蟲科(Calceostomatidae)。在Yamaguti (1963)的分類系統中, 無胄蟲歸于微溝蟲科。Ogawa等[1]根據無胄蟲的幼蟲具8對邊緣小鉤, 而微溝蟲科的幼蟲只有3對針狀刺(spicules), 以及二者腸支和前吸器的差異, 將無胄蟲亞科從微溝蟲科中獨立出來,并將其提升至科水平。在Boeger和Kritsky[10]的分類系統中, 無胄蟲科歸于三代蟲目, 該三代蟲目包括兩個分支: 一個分支由無胄蟲科、似四鉤蟲科(Tetraonchoididae)和溝穴蟲科(Bothitrematidae)組成,另一個分支包含三代蟲科(Gyrodactylidae)、棘杯蟲科(Acanthocotylidae)和Udonellidae科。Justine等[11]在利用28S rDNA D1區部分序列探討新鞋口蟲科(Neocalceostomatidae)和溝穴蟲科的分類地位時, 發現無胄蟲科、溝穴蟲科和Sundanonchidae科聚為一支, 該結果與Olson等[12]利用18S rDNA構建的進化樹一致, 即無胄蟲科和Sundanonchidae聚為一支。
分子系統學能夠解決缺乏形態特征或形態特征無法解決的某些類群的分類地位問題, 而28S rDNA 5′端序列適合推斷種間、屬間及近緣科間的親緣關系[13—15]。有鑒于此, 本文擬依據所獲標本對鯛無胄蟲的形態特征作一補充描述, 并利用28S rDNA C1-D2區部分序列探討了無胄蟲科的分類地位。
1 材料與方法
1.1 材料來源及形態學研究
宿主魚黑鯛取自廣東大亞灣養殖網箱, 先用肉眼觀察其體表、鰭、眼、鼻、口腔等部位, 再用淡水侵泡3—5min, 后取出魚鰓, 置于盛有過濾海水的培養皿中, 在解剖鏡下進行觀察。檢獲的單殖吸蟲標本, 部分用布氏膠直接封片; 部分用70%酒精固定后經洋紅或胭脂蟲紅染色, 梯度酒精脫水, 松油醇透明后用中性樹膠封片; 其余標本用95%酒精固定保存。照片圖為Olympus顯微數碼相機拍攝, 墨線圖借助Olympus LB描繪器進行手繪、掃描成電子圖片后用Photoshop CS4復墨。在光學顯微鏡下觀察并測量, 量度單位以μm計, 量度取平均數(括號內數值為所測標本的量度變化范圍)。標本保存于華南師范大學生命科學學院魚類寄生蟲學研究室。
1.2 DNA提取與PCR擴增
模板DNA的制備、28S rDNA部分序列的PCR擴增及PCR產物的純化等參見丁雪娟和廖翔華[15]。PCR擴增引物參照Hassouna等[16]: 正向C1 (5′-AC CCGCTGAATTTAAGCAT-3′) 和反向D2 (5′-TG GTCCGTGTTTCAAGAC-3′)。擴增反應體系為50 μL: 10×PCR buffer 5.0 μL, 2 mmol/L dNTP 5.0 μL, 25 mmol/ L MgCl24.0 μL, 10 μmol/L引物各2.0 μL, Taq酶(5 U/μL) 0.5 μL, DNA模板液3.0 μL, 滅菌雙蒸水28.5 μL。擴增反應步驟: 94℃預解鏈5min; 然后94℃ 1min, 56℃ 1min, 72℃ 1min, 30個循環; 最后72℃延伸5min。PCR產物經電泳、回收、純化后送生工生物工程(上海)股份有限公司進行序列測定。
1.3 系統發育樹構建
根據自測的無胄蟲及鱗盤蟲28S rDNA C1-D2區部分序列, 及從NCBI上下載的26種單殖吸蟲的相應序列(表 1), 構建無胄蟲相關類群的系統發育樹。DNA序列用ClustalX[17]軟件比對分析, 并手工刪除兩端無效序列。采用最大似然法(Maximum likelihood, ML) 和貝葉斯分析(Bayesian analysis, BA)構建系統樹。ML分析由在線軟件The CIPPES Science Gateway (http://www.phylo.org) 生成, 使用RAxML-HPC2 on XSEDE工具, 模型選擇GTRCAT, Bootstrap設置為1000次。貝葉斯分析使用軟件Mr-Modeltest2.1.2[18]搜索最佳模型參數(基于hLRT標準)。兩種方法均選用4種寡鉤亞綱單殖吸蟲作為外類群。
2 結果
2.1 鯛無胄蟲Anoplodiscus spari (Yamaguti, 1958) Ogawa & Egusa, 1981
宿主: 黑鯛Acanthopagrus schlegeli (Bloch)
寄生部位: 鰭和體表
標本采集時間和地點: 2014年11月; 廣東惠州大亞灣(22°43′N, 114°28′E)
形態描述: 蟲體呈梭形, 大小為1563 (1129—2062)×652 (370—999)。體前端平截, 上有1對吸槽、邊緣不規則。眼點2對。咽發達, 卵圓形, 大小91 (102—150)×102 (91—138)。腸支單管, 具眾多不對稱的側分支(圖 1A)。
睪丸單個, 圓球形, 大小287 (156—338)×230 (165—281)(圖 1A)。交接器由交接管和支持器組成; 交接管近環形, 管長414 (316—518), 基部位于支持器的下方, 末端位于三分叉中支外側; 支持器一端呈三叉狀展開, 其中一分叉較長且基部明顯膨大(圖 1B)。
卵巢卵圓形, 位于睪丸前, 大小127 (84—166)× 126 (90—166)。陰道開口于蟲體右側, 無幾丁質結構。卵黃腺呈大濾泡狀, 分布自交接器水平至腸支末端之前(圖 1A)。
描述依據3個胭脂蟲紅、2個明礬洋紅染色標本和2個布氏膠封片標本。
2.2 無胄蟲科的分類地位
首次測得鯛無胄蟲的28S rDNA C1-D2部分序列。原始序列由Clustal X比對后經手工刪除兩端無效序列, 最終組成一個由24種多鉤類和4種寡鉤類單殖吸蟲構成的數據集, 序列長度647—963 bp。比對后序列全長1133 bp, 其中簡約信息位點725個,保守位點248個, 變異位點818個。用Mrmodeltest軟件估計的最適替代模型是GTR+I+G模型(Nst=6; Rates=invgamma)。生成的BA和ML樹, 在拓撲結構上沒有差異, 僅節點支持率有所不同。本文選取了ML的50%多數一致樹, 并附有BA的PP (%)和ML的Bootstrap值(圖 2)。
分子系統樹顯示: 各個科以較高的支持率聚為一支, 均是單系群。系統樹由4個分支組成, 其中基部分支由三代蟲科和Udonellidae科構成; 無胄蟲科聚入指環蟲目分支內, 與四鉤蟲科(Tetraonchidae)互為姐妹群, 同樣被納入該分支的還有指環蟲科、錨首蟲科和鱗盤蟲科; 微溝蟲科與單杯科組成一個分支; 分室科獨自構成一個分支。4個分支均擁有較高的分支支持率(70%以上)。
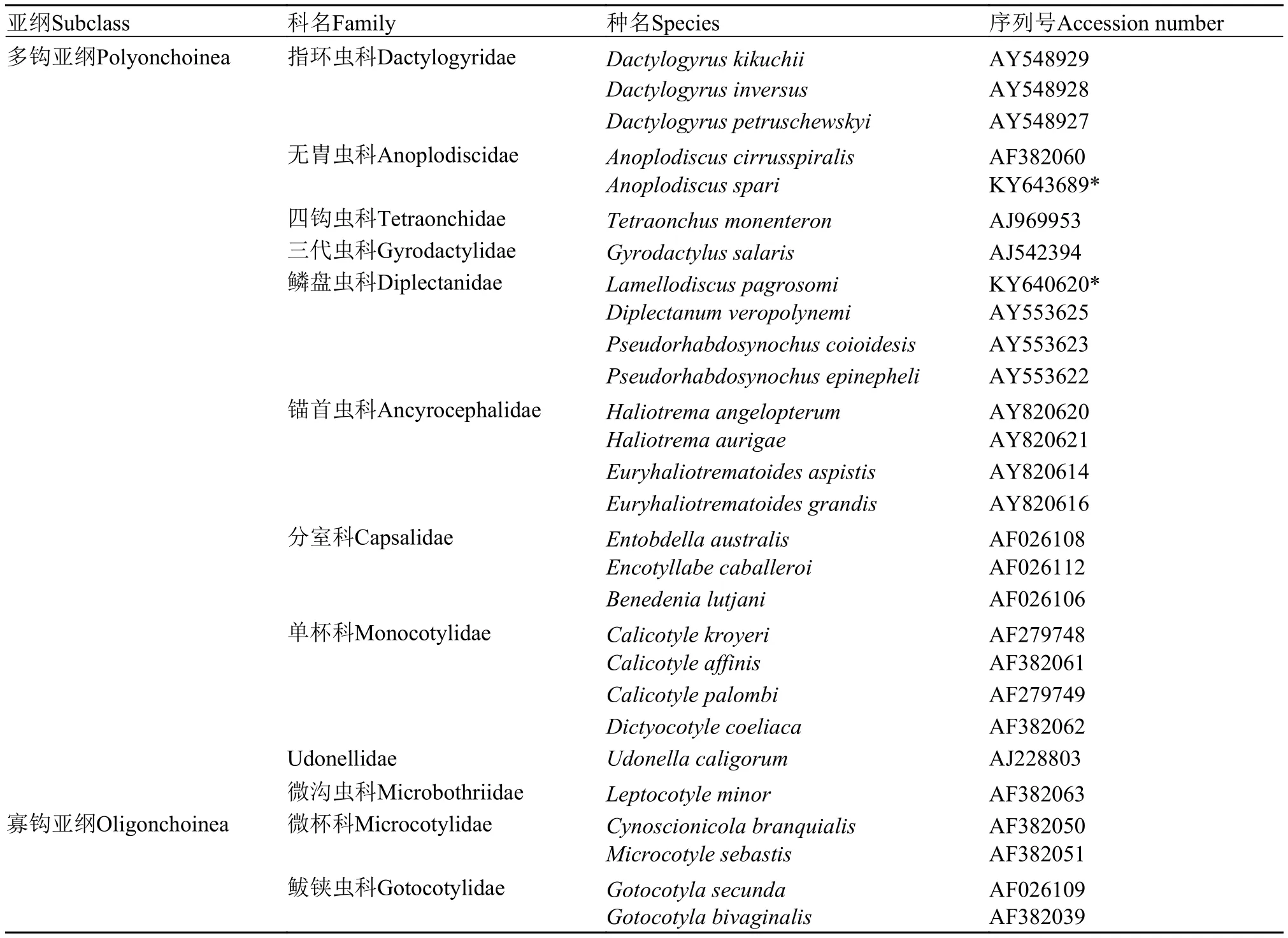
表 1 系統分析所涉及的單殖吸蟲種類及序列信息Tab.1 List of monogenean species and GenBank accession numbers used in the present phylogenetic analyses
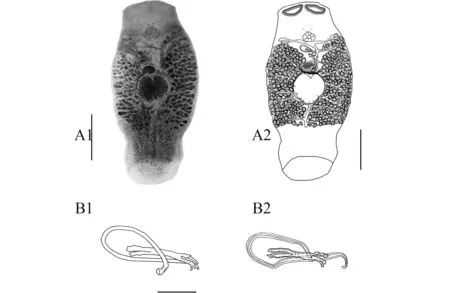
圖 1 鯛無胄蟲Fig.1 Anoplodiscus spari (Yamaguti, 1958) Ogawa et Egusa, 1981
3 討論
本文對采自我國南海大亞灣的鯛無胄蟲作了簡單描述, 并補充了整體圖和交接器結構圖, 所獲標本與Ogawa等(1981)的描述基本一致, 但交接管基部和末端位置略有差異: 在Ogawa等的描述中,交接管基部靠近支持器三叉處、末端從三叉處延伸并略彎曲(圖B2), 而我們的標本中交接管基部在支持器的下方、末端從三分叉的中支略為突出(圖B1)。
在Boeger和Kristky[10]的分類系統中, 無胄蟲科與Udonellidae在同一支內, 均隸屬于三代蟲目, 表明二者具有較近的親緣關系, 且Udonellidae科的后吸器亦缺乏幾丁質結構; 但本研究顯示無胄蟲科與Udonellidae科位于兩個不同的分支內: 無胄蟲科位于指環蟲目內、Udonellidae科屬于三代蟲目。生活史上, 無胄蟲的幼蟲階段具有纖毛和8對幾丁質邊緣小鉤[1], 而Udonella的幼蟲及成蟲階段均沒有纖毛也沒有幾丁質鉤[19]。形態結構上, 無胄蟲的邊緣小鉤和腸支結構與四鉤蟲和似四鉤蟲的更為相似。超顯微結構上, 無胄蟲與指環蟲類的精子都為“4型”(單一軸絲, 無微管) 而Udonella是“2型”(雙軸絲, 無微管)[20]。在本文的分子系統樹上, 無胄蟲科與四鉤蟲科互為姐妹群, 而與三代蟲所在的進化支相距甚遠。無胄蟲曾被歸入微溝蟲科, 后Ogawa等[1]認為其為一獨立的科, 本研究結果顯示無胄蟲科是一獨立的科, 與微溝蟲科分屬于不同的進化支內。綜上所述, 我們認為無胄蟲成蟲幾丁質結構的缺失為一種同型性狀, 而非共同衍征, 支持Justine等[11]的觀點, 即無胄蟲科應歸入指環蟲目。

圖 2 基于28S rDNA部分序列構建的無胄蟲科及相關類群的分子系統樹Fig.2 The phylogenetic tree of Anoplodiscidae and some related groups based on the partial 28S rDNA sequences
本研究證實了無胄蟲科為一獨立的科, 支持將無胄蟲科納入指環蟲目、四鉤蟲亞目。無胄蟲成蟲后吸器幾丁質缺失是一種次生現象[21], 提示在研究探討單殖吸蟲的分類地位時, 不能僅依據成蟲的形態結構, 還應考慮其生活史各個階段的形態特征。另幾丁質結構是單殖吸蟲分類鑒定的主要依據[15], 但對于缺乏或生活史某個階段缺乏幾丁質結構的蟲種(如本文的無胄蟲), 單純依賴形態特征的分類就顯得尤為不足, 此時分子系統學能有效地彌補這一缺陷, 為單殖吸蟲的正確分類提供科學依據。
致謝:
華南師范大學生命科學學院張劍英教授提供相關文獻, 劉琳副教授協助英文潤色, 2013級研究生方偉協助采集標本, 謹致謝忱。
[1]Ogawa K, Egusa S.The systematic position of the genus Anoplodiscus (Monogenea: Anoplodiscidae) [J].Systematic Parasitology, 1981, 2(4): 253—260
[2]Paraguassú A R, Luque J L, Alves D R.A new species of Anoplodiscus (Monogenea: Anoplodiscidae) parasitic on Pagrus pagrus (Osteichthyes: Sparidae) from the coastal zone of the State of Rio de Janeiro, Brazil [J].Memórias do Instituto Oswaldo Cruz, 2002, 97(8): 1197—1199
[3]Roubal F R, Armitage J, Rohde K.Taxonomy of Metazoan ectoparasites of snapper, Chrysophrys autratus (Family Sparidae), from southern Australia, eastern Australia and New Zealand [J].Australian Journal of Zoology, 1983, 31(94): 1—68
[4]Ogawa K.Anoplodiscus tai sp.nov.(Monogenea: Anoplodiscidae) from cultured red sea bream Pagrus major [J].Fish Pathology, 1994, 29(1): 5—10
[5]Roubal F R.The taxonomy and site specificity of the metazoan ectoparasites on the black bream, Acanthopagrus australis (Günther), in northern New South Wales [J].Australian Journal of Zoology, Supplementary Series, 1981, 30(84): 1—100
[6]Zhang J Y, Qiu Z Z, Ding X J.Parasites and Parasitic Diseases of Fishes [M].Beijing: Science Press.1999, 205 [張劍英, 邱兆祉, 丁雪娟.魚類寄生蟲與寄生蟲病.北京: 科學出版社.1999, 205]
[7]Wu J Y, Lu J Y, Hu Y S.A new species and a new Chinese record of monogeneans from marine fishes in the South China Sea [J].Acta Zootaxonomica Sinica, 2002, 27(4): 677—684 [吳金英, 呂軍儀, 胡應劭.寄生于南海魚類中的單殖吸蟲一新種和一新紀錄(吸蟲綱: 單殖目).動物分類學報, 2002, 27(4): 677—684]
[8]Zhang J Y, Yang T B, Liu L, et al.A list of monogeneans from Chinese marine fishes [J].Systematic Parasitology, 2003, 54(2): 111—130
[9]Xie J J, Xu W J, Shi H, et al.The diagnosis and prevention of Anoplodicus spari [J].Scientific Fish Farming, 2014, 2: 63—64 [謝建軍, 許文軍, 施慧, 等.黑鯛無胄蟲病的診斷與防治.科學養魚, 2014, 2: 63—64]
[10]Boeger W A, Kritsky D C.Phylogenetic relationships of the Monogenoidea [A].In: Littlewood D T J, Bray R A(Eds.), Interrelationships of the Platyhelminthes [C].London: Taylor & Francis.2001, 92—102
[11]Justine J L, Jovelin R, Neifar L, et al.Phylogenetic positions of the Bothitrematidae and Neocalceostomatidae (Monopisthocotylean Monogeneans) inferred from 28S rDNA sequences [J].Comparative Parasitology, 2002, 69: 20—25
[12]Olson P D, Littlewood D T J.Phylogenetics of the Monogenea - evidence from a medley of molecules [J].International Journal for Parasitology, 2002, 32(3): 233—244
[13]Jovelin R, Justine J L.Phylogenetic relationships within the polyopisthocotylean monogeneans (Platyhelminthes) inferred from partial 28S rDNA sequences [J].International Journal for Parasitology, 2001, 31(4): 393—401
[14]Mollaret I, Jamieson B G M, Justine J L.Phylogeny of the Monopisthocotylea and Polyopisthocotylea (Platyhelminthes) inferred from 28S rDNA sequences [J].International Journal for Parasitology, 2000, 30(2): 171—185
[15]Ding X J, Liao X H.Phylogenetic position of the monogeneans Pseudodactylogyrus, Heternchocleidus and Trianchoratus inferred from the 5’ terminal sequences of 28S rDNA [J].Acta Zootaxonomica Sinica, 2005, 30(2): 244—251 [丁雪娟, 廖翔華.偽指環蟲、異鉤蟲和三鉤蟲的系統位置.動物分類學報, 2005, 30(2): 244—251]
[16]Hassouna N, Mithot B, Bachellerie J P.The complete nucleotide sequence of mouse 28S rRNA gene.Implications for the process of size increase of the large subunit rRNA in higher eukaryotes [J].Nucleic Acids Research, 1984, 12(8): 3563—3583
[17]Thompson J D, Gibson T J, Plewniak F, et al.The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools [J].Nucleic Acids Research, 1997, 25(24): 4876—4882
[18]Posada D, Crandall K A.Modeltest: testing the model of DNA substitution [J].Bioinformatics, 1998, 14(9): 817—818
[19]Littlewood D, Rohde K, Clough K A.The phylogenetic position of Udonella (Platyhelminthes) [J].International Journal for Parasitology, 1998, 28(8): 1241—1250
[20]Justine J L, Lambert A, Mattei X.Spermatozoon ultrastructure and phylogenetic relationships in the monogeneans (Platyhelminthes) [J].International Journal for Parasitology, 1985, 15(6): 601—608
[21]Whittington I D.The capsalidae (Monogenea: Monopisthocotylea): a review of diversity, classification and phylogeny with a note about species complexes [J].Folia Parasitologica, 2004, 51(2—3): 109—122
DESCRIPTION OF ANOPLODICUS SPARI (MONOGENEA: ANOPLODISCIDAE) AND THE TAXONOMIC POSITION OF ANOPLODISCIDAE
ZHOU Zi-Hua, CAI Xing and DING Xue-Juan
(School of Life Science, South China Normal University, Guangdong Provincial Key Laboratory for Healthy and Safe Aquaculture, Guangzhou Key Laboratory of Subtropical Biodiversity and Biomonitoring, Guangzhou 510631, China)
The current study described the whole worm and male copulatory organ of Anoplodicus spari collected from Acanthopagrus schlegeli off Daya Bay, South China Sea.Our specimens of Anoplodicus spari from South China Sea was basically consistent with the description of Ogawa et al.(1981) except for the slight difference in the position of the base and distal end of copulatory tube.The proximal end of the copulatory tube lied beneath the accessory piece and the distal end protruded from the three rami in our specimens, while the distal end extended from the rami and curved in the specimen of Ogawa (1981).The partial 28S rDNA sequences (C1-D2 domain) of Anoplodicus spari and other related groups were used to study the taxonomic position of Anoplodiscidae.The results showed the independent position of Anoplodiscidae, a valid family belonging to the clade of Tetraonchinea, Dactylogyridea.Anoplodiscidae and Tetraonchidae were a sister group in the phylogenetic tree, and this clade was distantly different from Udonellidae and Microbothriidae.
Anoplodicus spari; Anoplodiscidae; 28S rDNA; Taxonomic position
Q959.19
A
1000-3207(2017)04-0891-05
10.7541/2017.111
2016-07-01;
2016-11-07
國家自然科學基金項目(31172051和31572232)資助 [Supported by the National Natural Science Foundation of China (31172051, 31572232)]
周梓華(1992—), 男, 廣東南海人; 碩士研究生; 主要從事海水魚類單殖吸蟲研究。E-mail: 289462085@qq.com
丁雪娟, E-mail: dingxj@scnu.edu.cn
猜你喜歡
西北民族大學學報(自然科學版)(2021年4期)2021-12-29 02:54:24
數學小靈通(1-2年級)(2021年4期)2021-06-09 06:25:56
大眾健康(2021年6期)2021-06-08 19:30:06
小聰仔(科普版)(2020年12期)2021-01-18 09:16:52
東方少年·布老虎畫刊(2020年4期)2020-06-08 15:48:10
學生天地(2019年32期)2019-08-25 08:55:22
中學生數理化·七年級數學人教版(2019年4期)2019-05-20 10:06:32
中學生數理化·七年級數學人教版(2018年6期)2018-06-26 08:36:06
小天使·一年級語數英綜合(2017年11期)2017-12-05 18:49:56
初中生世界·七年級(2017年9期)2017-10-13 22:27:46
