不同穗型小麥小花發育過程中幼穗內同化物分配與穗粒數的關系
2017-09-01 00:29:02文祥朋孫克剛鄭春風
江西農業學報 2017年8期
關鍵詞:差異
文祥朋,任 偉,孫克剛,杜 君,鄭春風
(1.河南省新鄉縣農牧局,河南 新鄉 453700;2.河南省農業科學院 小麥研究所,河南 鄭州 450002; 3.河南省農業科學院 植物營養與資源環境研究所/河南農業生態與環境重點實驗室,河南 鄭州 450002)
不同穗型小麥小花發育過程中幼穗內同化物分配與穗粒數的關系
文祥朋1,任 偉2,孫克剛3*,杜 君3,鄭春風3
(1.河南省新鄉縣農牧局,河南 新鄉 453700;2.河南省農業科學院 小麥研究所,河南 鄭州 450002; 3.河南省農業科學院 植物營養與資源環境研究所/河南農業生態與環境重點實驗室,河南 鄭州 450002)
分別以大穗型、多穗型小麥品種周麥16、豫麥49-198為試驗材料,重點研究了大穗型和多穗型小麥品種在小花發育過程中,小花發育成粒與同化物分配之間的關系。在小麥幼穗發育過程中,通過比較分析2個穗型品種的同化物分配差異發現,在小花發育中后期,大穗型品種周麥16轉移到穗部的同化物比多穗型品種豫麥49-198多。花期完善小花數與穗干重、穗氮積累量、莖干重、莖氮積累量呈正相關關系,但相關性不顯著,與穗/莖干重、穗/莖氮積累量呈顯著正相關,與穗重/穗氮積累量呈極顯著正相關。大穗型周麥16在穗/莖干重、穗/莖氮積累量、穗重/穗氮積累量上顯著高于豫麥49-198,且在穗重/穗氮積累量上達到極顯著水平。前者在花期單位面積完善小花數較后者增加3.85%,且差異達到極顯著水平。花期完善小花的形成與穗器官同化物供給總量的關系不大,而與同化物在穗部和營養器官的分配比例密切相關,與同化物在穗部的分配比例關系更加密切。大穗型周麥16較高的穗粒數與花期較高的完善小花數密切相關,且以穗部和營養器官間及穗部自身較高的同化物分配比例(尤其是穗重/穗氮積累量)取得大穗多粒。
不同穗型品種;小花發育;同化物分配;穗粒數
在當前高產水平下,需在穩定穗數的前提下,把提高粒重和增加穗粒數作為進一步提高小麥產量的突破口[1-4]。但由于受環境條件和品種遺傳特性的制約,增加粒重相對比較有限,穗粒數的提高應成為提高小麥產量的關鍵[5-8]。由于小麥穗粒數的形成是小花發育和結實成粒等一系列復雜生理過程的最終體現[9-10],因此,明確小花發育的生理變化特征,探明小花發育結實成粒的規律,對于深化小麥產量生理的研究和提高穗粒數,具有十分重要的理論和實際意義。李存東等[11]對冬、春性小麥品種小花發育的動態模式進行了研究,王兆龍等[12]對大穗型和小穗型小麥品種的小花結實特性進行了報道,王俊英等[13]對小花發育規律進行了研究。朱云集等[14]指出合理施用氮肥能夠增加小花的數目,減少小花退化,增加穗粒數。Ferrante等[15]認為硬粒小麥可孕花結實成粒與氮肥有效性密切相關。Demotes等[16]研究發現小花發育期穗部氮素積累量與籽粒數呈顯著正相關。前人有關植株碳氮代謝與小麥產量關系的研究已有較多報道[17-18],然而,對2個不同穗型小麥品種在小花發育成粒過程中同化物分配與穗粒數的關系研究少見報道。本試驗重點研究了大穗型和多穗型小麥品種在小花發育過程中,小花發育成粒與同化物分配間的關系,旨在為大穗型小麥品種的栽培調控和培育提供理論依據。
1 材料與方法
1.1 試驗材料與設計
本研究于2012~2014年在河南農業大學科教示范園區(3486′ N,11359′ E)進行了田間試驗,土壤質地為壤土,0~20 cm土層有機質含量為10.6 g/kg、全氮為0.9 g/kg、堿解氮為82.1 mg/kg、速效磷為25.6 mg/kg、速效鉀為124.5 mg/kg、有效硼為0.41 mg/kg。總施氮量146 kg/hm2(尿素,含N 46%),50%于播前基施,其余50%于拔節期追施。播前每公頃施磷肥(P2O5)150 kg,鉀肥(K2O)120 kg。以大穗型品種周麥16、多穗型品種豫麥49-198為供試材料,2年試驗材料均于10月8日播種,基本苗為2.25×106株/hm2,行距為20 cm,每處理小區面積為20 m2,隨機區組設計,重復3次。試驗田栽培管理同一般高產田。
1.2 測定內容與方法
自小麥3葉期開始取樣,每隔7 d取樣1次,以有完整綠色花藥的小花為完善小花,每小區選擇生長均勻一致的小麥植株5株(每處理共計15株),在EMZ-TR解剖鏡下觀察記載主莖幼穗分化進程,并觀察記載分化小穗數、小花數及幼穗分化各階段特征。
播后累積生長度日(GDD)=[(Tmax+Tmin)/2]-Tb
公式中,Tmax:最高溫度,Tmin:最低溫度,Tb:基礎溫度。
分別于拔節期、拔節后7 d、拔節后14 d、拔節后21 d、拔節后28 d、孕穗期、抽穗期、開花期、成熟期取樣,105 ℃下殺青30 min,80 ℃烘至恒重,粉碎保存,用于植株干物質和氮含量的測定。
產量及其構成因素:成熟期每小區隨機取20株,按常規考種法記載每穗結實粒數,實收5 m2計產。
1.3 數據分析
采用Microsoft Excel 2003和SPSS 17.0軟件對2012~2013年度和2013~2014年度2個年度的數據進行處理分析及繪圖。
2 結果與分析
2.1 小花發育過程中幼穗內同化物的分配變化
由圖1可知,在拔節后14 d之前,大穗型周麥16與多穗型豫麥49-198的穗/莖干物重無顯著差異。拔節后14 d之后,周麥16的穗/莖干物重顯著高于豫麥49-198。由此表明,在小花發育中后期,大穗型小麥品種周麥16轉移到穗部的同化物比多穗型小麥品種豫麥49-198多,為大穗的形成提供了良好的物質基礎。
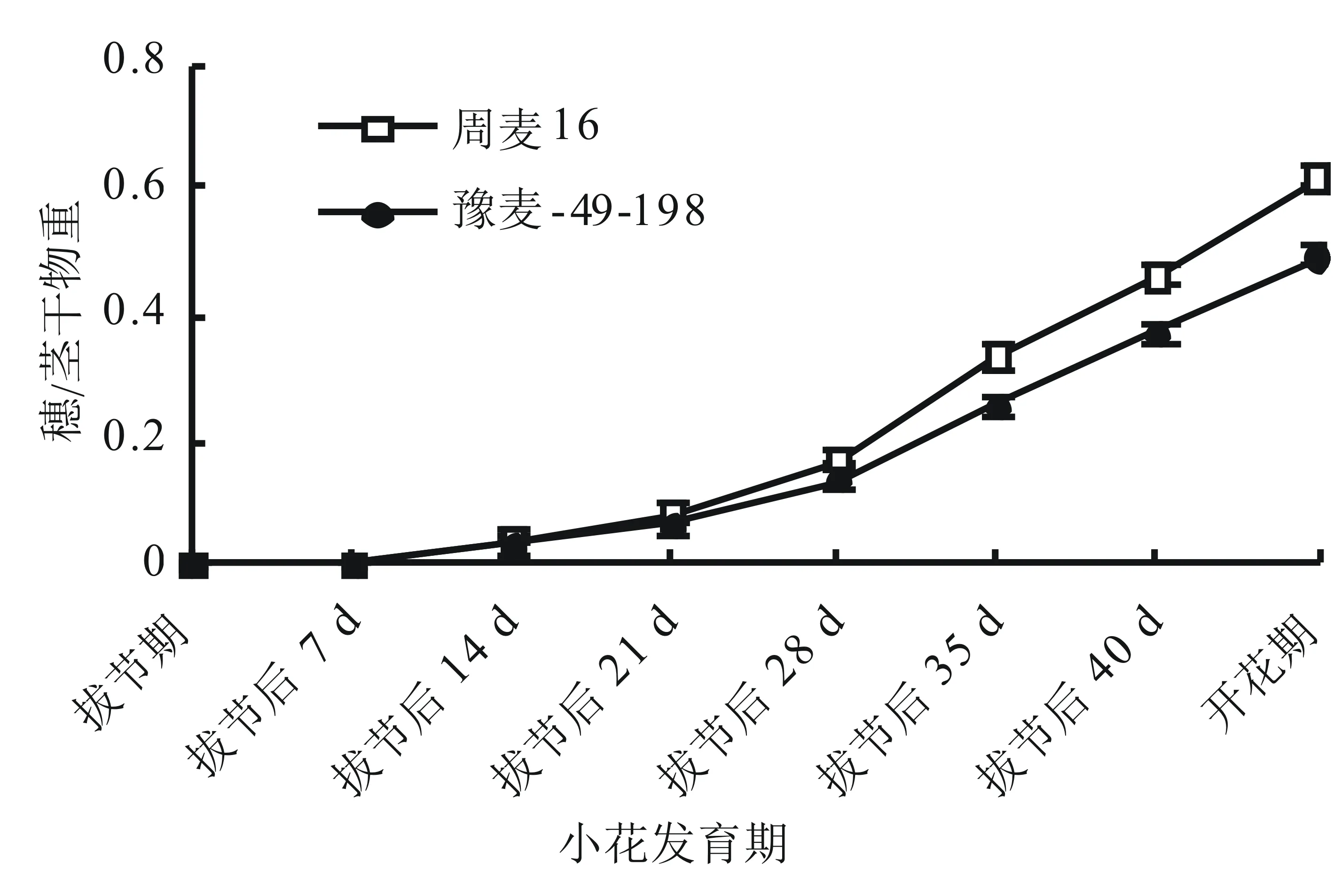
莖包含莖、葉、鞘,下同。
由圖2可知,在拔節后21 d之前,大穗型周麥16的穗/莖氮積累量顯著低于多穗型豫麥49-198。而在拔節后21 d之后,周麥16的穗/莖氮積累量顯著高于豫麥49-198。這表明在小花發育后期,大穗型小麥品種周麥16轉移到穗部的氮量比多穗型小麥品種豫麥49-198多。
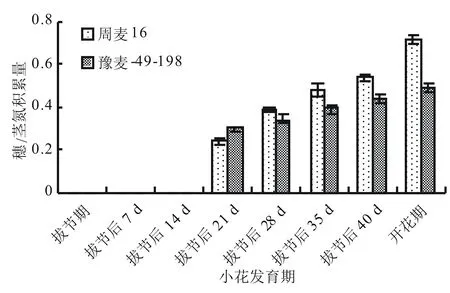
圖2 2種穗型小麥在小花發育期主莖的穗/莖氮積累量
2.2 開花期2種穗型小麥的同化物分配與完善小花數的差異分析
由表1可知,多穗型豫麥49-198在花期單位面積的莖干重、穗氮積累量和莖氮積累量上顯著高于大穗型周麥16,但在穗/莖干重、穗/莖氮積累量、穗重/穗氮積累量上顯著低于周麥16,且在穗重/穗氮積累量上達到極顯著水平。進一步分析可知,大穗型周麥16在開花期單位面積完善小花數較多穗型豫麥49-198增加3.85%,且差異達到極顯著水平。
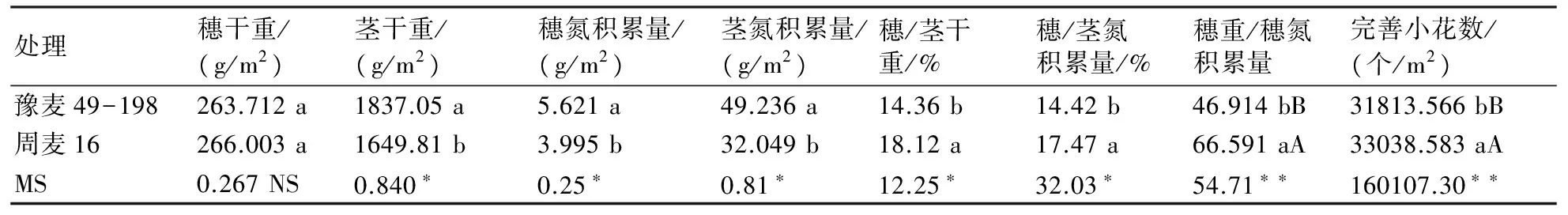
表1 開花期同化物分配與完善小花數及穗粒數的差異分析
注:同列小、大寫字母分別表示在0.05、0.01水平上的差異顯著性,字母相同則差異不顯著,不同則顯著。
2.3 開花期同化物分配與完善小花數的相關分析
由表2可知,開花期單位面積的完善小花數與穗干重、穗氮積累量、莖干重、莖氮積累量呈正相關關系,但相關性不大;與穗/莖干重、穗/莖氮積累量呈顯著正相關;與穗重/穗氮積累量的相關系數達到0.9以上,呈極顯著正相關。
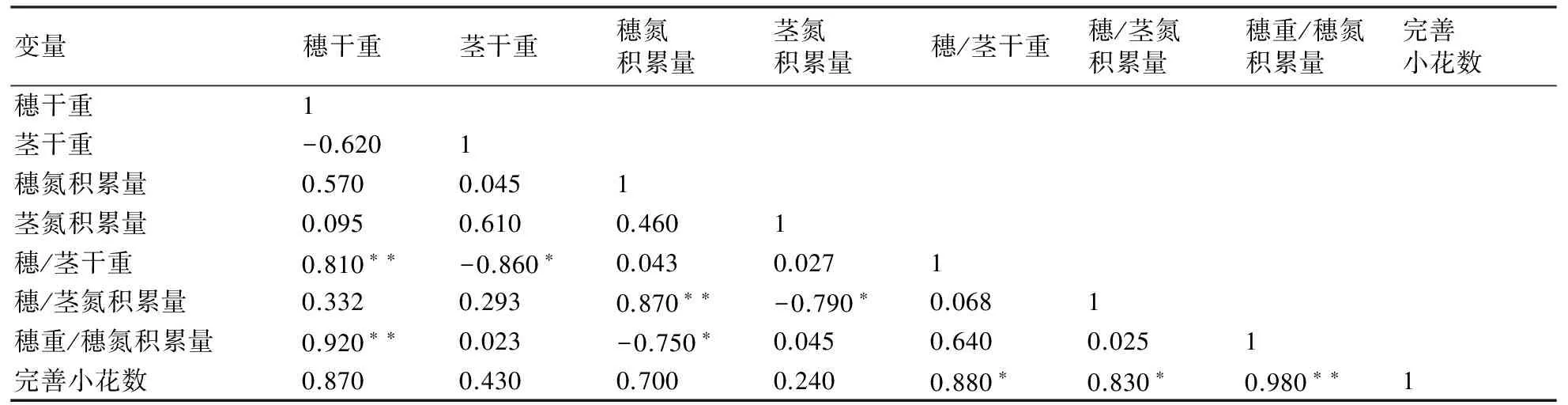
表2 開花期同化物分配與完善小花數及穗粒數的相關分析
注:*、**分別表示在0.05、0.01水平上的相關顯著性。
2.4 2種穗型小麥總分化小花數、完善小花數和結實粒數的差異
從圖3可知,大穗型周麥16和多穗型豫麥49-198在完善小花數和結實粒數上存在較大差異,且差異達到顯著水平。大穗型周麥16單位面積完善小花數、結實粒數分別為38018.82個和22439.87個,多穗型豫麥49-198單位面積完善小花數、結實粒數分別為25498.38個和14852.65個。然而,在分化小花數上,大穗型周麥16單位面積分化小花數高于多穗型豫麥49-198,但差異不顯著。
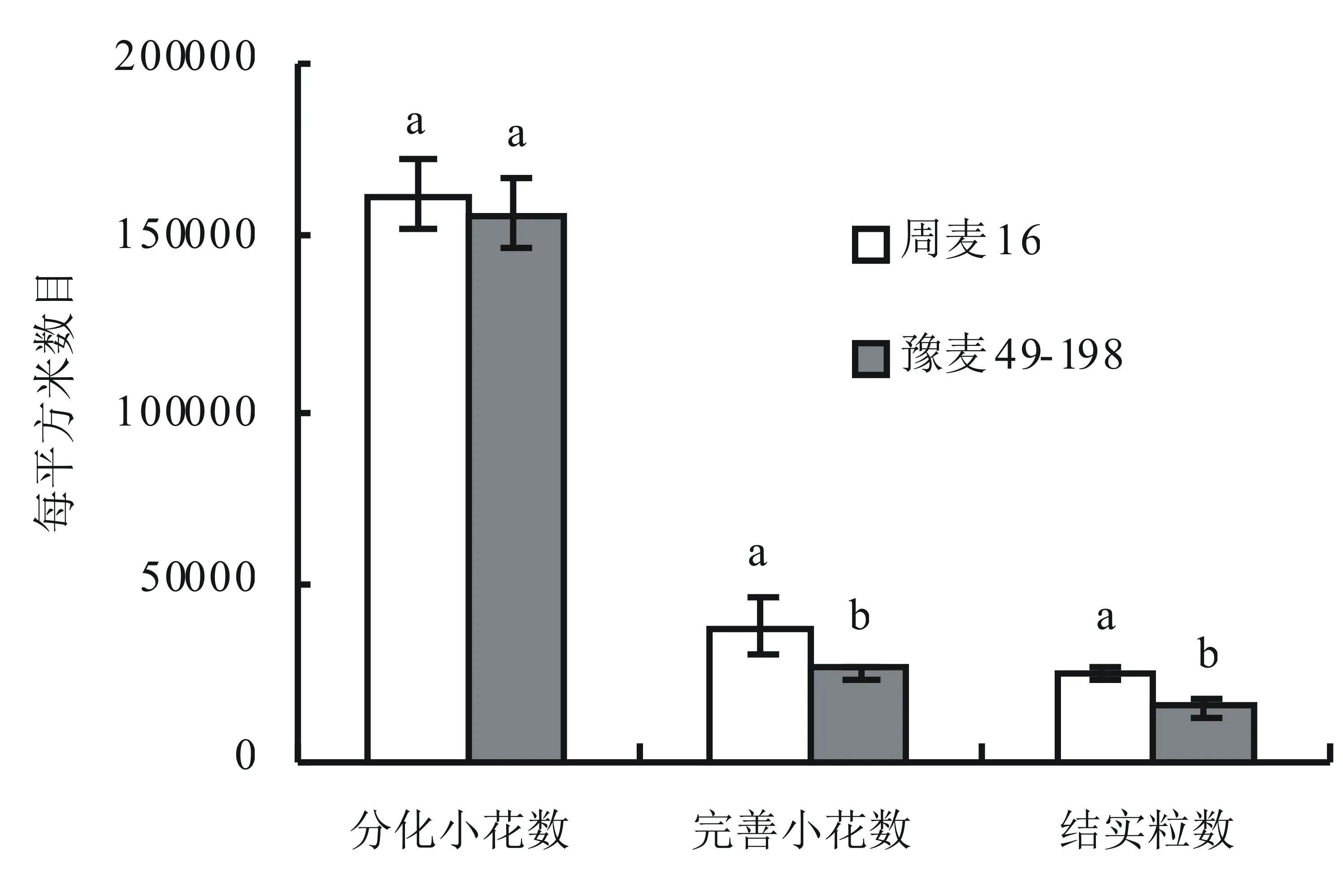
圖3 分化小花數、可孕小花數和結實粒數的差異
3 結論與討論
從我國華北地區小麥產量及其構成因素實際調查情況分析,成穗數一般為600萬/hm2左右,千粒重在40 g以上,但目前所推廣小麥品種每穗粒數一般在30粒上下[5]。根據生態學最小因子限制定律,穗粒數少已成為提高產量的最小因子,即限制小麥籽粒產量提高的短板。若在產量構成因素成穗數、穗粒數和千粒重三者之間各增加1個單位,以穗粒數對產量的貢獻絕對值最高[19]。因此,在我國華北的小麥生產中,發揮穂分化時間長的優勢,成大穂是增加產量的主要途徑。李存東等[20]通過對不同播期下小麥冬、春性品種小花結實特性研究,提出穗粒數與可孕小花數及其結實率密切相關,而與總小花數及其結實率無明顯聯系。本研究結果表明,大穗型周麥16和多穗型豫麥49-198兩種穗型小麥品種在完善小花數和結實粒數上存在較大差異,且差異達到顯著水平,但是,在總分化小花數上,兩者差異不明顯。這說明穗粒數與花期完善小花的形成存在著密切聯系,且與總分化小花數關系不大,此結果與前人研究相一致。
在開花前,小麥的營養器官處于快速生長期,尤其是在拔節后節間迅速伸長時期[21-22]。Miralles等[23]認為,限制穗粒數形成的一個重要因素是幼穗與莖稈在快速生長期對同化物的激烈競爭。本研究發現,在拔節后14 d之后,大穗型周麥16的穗/莖干物重顯著高于多穗型豫麥49-198;在拔節后21 d之后,大穗型周麥16的穗/莖氮積累量顯著高于多穗型豫麥49-198。這表明在小花發育中后期,大穗型小麥品種周麥16有較多的同化物轉移到穗部,這為大穗的形成奠定了良好的物質基礎,此結果與Slafer等[24]認為形成大穗的重要物質基礎需要較多的同化物轉移到穗部的研究結果相一致。Ellen[25]指出致使小花急劇退化的原因與碳水化合物總供給量關系不大,而與營養器官和穗部的同化物分配比例有很大聯系。本研究發現,花期單位面積的完善小花數與穗干重、穗氮積累量呈正相關關系,但相關性不顯著,而與穗/莖干重、穗/莖氮積累量呈顯著正相關,與穗重/穗氮積累量呈極顯著正相關,這表明花期完善小花的形成與穗器官同化物供給總量的關系不大,而與同化物在穗部和營養器官的分配比例密切相關,此結果與其研究結果一致,并進一步指出,花期完善小花數的形成與同化物在穗重/穗氮積累量上的分配比例呈極顯著正相關。
[1] 于振文,田奇卓,潘慶民,等.黃淮麥區冬小麥超高產栽培的理論與實踐[J].作物學報,2002,28(5):577-585.
[2] 王兆龍,曹衛星,戴廷波.小麥穗粒數形成的基因型差異及增粒途徑分析[J].作物學報,2001,27(2):236-242.
[3] Fischer R A. The importance of grain or kernel number in wheat: A reply to Sinclair and Jamieson[J]. Field Crops Research, 2008, 105(1): 15-21.
[4] Sinclair T R, Jamieson P D. Grain number, wheat yield, and bottling beer: an analysis[J]. Field Crops Research, 2006, 98(1): 60-67.
[5] 崔金梅,郭天財,朱云集,等.小麥的穗[M].北京:中國農業出版社,2008:284-286.
[6] 馬元喜,王晨陽,朱云集.協調小麥幼穗發育三個兩極分化過程增加穗粒數[M]//盧良恕.中國小麥栽培研究新進展.北京:農業出版社,1991:119-126.
[7] 王志敏.小麥穗粒數的調節[D].北京:北京農業大學,1994.
[8] 鄭春風,朱慧杰,朱云集,等.冬小麥小花發育及結實特性對葉面噴硼的響應[J].植物營養與肥料學報,2016,22(2):550-556.
[9] Zheng C F, Zhu Y J, Zhu H J, et al. Floret development and grain setting characteristics in winter wheat in response to pre-anthesis foliar applications of 6-benzylaminopurine and boron[J]. Field Crops Res, 2014, 169: 70-76.
[10] Ferrante A, Savin A, Slafer G A. Floret development of durum wheat in response to nitrogen availability[J]. Journal of Experimental Botany, 2010, 61(15): 4351-4359.
[1] Li C D, Cao W X, Dai T B. Dynamic characteristics of floret primordium development in wheat[J]. Field Crops Res, 2001, 71(1): 71-76.
[12] 王兆龍,曹衛星,戴廷波,等.不同穗型小麥品種小花發育與結實特性研究[J].南京農業大學學報,2000,23(4):9-12.
[13] 王俊英,趙春江,楊寶祝.小麥小花發育與退化的研究[J].華北農學報,1996,11(2):9-13.
[14] 朱云集,崔金梅,王晨陽,等.小麥不同生育時期施氮對穗花發育和產量的影響[J].中國農業科學,2002,35(11):1325-1329.
[15] Ferrante A, Savin A, Slafer G A. Floret development and grain setting differences between modern durum wheat under contrasting nitrogen availability[J]. Journal of Experimental Botany, 2013, 64: 169-184.
[16] Demotes-Mainard S, Jeuffroy M H. Effects of nitrogen and radiation on dry matter and nitrogen accumulation in the spike of winter wheat[J]. Field Crops Res, 2004, 87(2/3): 221-233.
[17] 王彥麗,朱云集,郭天財,等.冬小麥碳氮積累、轉運和籽粒產量對小花發育期追氮的響應[J].麥類作物學報,2011,31(1):98-105.
[18] 張敏,戴廷波,姜東,等.6-BA對小麥花后C/N物質運轉和籽粒品質的影響[J].南京農業大學學報,2006,29(4):6-10.
[19] 朱云集,崔金梅,郭天財,等.河南省小麥生產發展中幾個關鍵技術問題的商榷[J].河南農業科學,2011,40(8):54-57.
[20] 李存東,曹衛星,張月晨,等.不同播期下小麥冬春性品種小花結實特性及其與植株生長性狀的關系[J].麥類作物學報,2000,20(1):59-62.
[21] González F G, Slafer G A, Miralles D J. Floret development and spike growth as affected by photoperiod during stem elongation in wheat[J]. Field Crops Res, 2003, 81(1): 29-38.
[22] Slafer G A, Abeledo L G, Miralles D J, et al. Photoperiod sensitivity during stem elongation as an avenue to raise potential yield in wheat[J]. Euphytica, 2001, 119(1/2): 191-197.
[23] Miralles D J, Katz S D, Colloca A et al. Floret development in near isogenic wheat lines differing in plant height[J]. Field Crops Res, 1998, 59(1): 21-30.
[24] Slafer G A, Andrade F H. Physiological attributes to the generation of grain yield in bread wheat cultivars released at different eras[J]. Field Crops Res, 1993, 31(3/4): 351-367.
[25] Ellen J. Effects of nitrogen and plant density on growth, yield and chemical composition of two winter wheat cultivars[J]. Agron and Crop Sci, 1990, 164(3): 174-183.
(責任編輯:曾小軍)
Relationship between Photosynthate Distribution in Young Ear at Floret Development Stage and Grain Number per Ear of Different Ear-type Wheat Varieties
WEN Xiang-peng1, REN Wei2, SUN Ke-gang3*, DU Jun3, ZHENG Chun-feng3
(1. Farming and Animal Husbandry Bureau of Xinxiang County in Henan Province, Xinxiang 453700, China; 2. Wheat Research Institute, Henan Academy of Agricultural Sciences, Zhengzhou 450002, China; 3. Institute of Plant Nutrition, Resources and Environmental Sciences, Henan Academy of Agricultural Sciences / Henan Provincial Key Laboratory of Agricultural Ecology and Environment, Zhengzhou 450002, China)
The relationships between photosynthate distribution in young ear at floret development stage and grain number per ear of large-ear-type wheat variety Zhoumai 16 and multiple-ear-type wheat variety Yumai 49-198 were studied. It was found that Zhoumai 16 transferred more photosynthate to ear than Yumai 49-198 in middle and later period of floret development. The number of perfect florets in flowering period was significantly positively correlated with the ear-stem weight ratio and ear-stem nitrogen accumulation ratio, and was very significantly positively correlated with the ratio of ear weight to ear nitrogen accumulation. The ear-stem dry weight ratio, and ear-stem nitrogen accumulation ratio of Zhoumai 16 were significantly higher than those of Yumai 49-198, and the ear weight / ear nitrogen accumulation of the former was very significantly higher than that of the latter. The number of perfect florets per unit area of Zhoumai 16 in flowering period was 3.85% higher than that of Yumai 49-198, and the difference reached a very significant level. The formation of perfect florets in flowering period was closely related to the distribution proportion of photosynthates in ear and vegetation organ (especially in ear). As for Zhoumai 16, higher grain number per ear was closely related to more perfect florets in flowering period, and the acquirement of large ears and more grains per ear depended on high distribution proportion of photosynthates in ear (especially ear weight / ear nitrogen accumulation).
Different ear-type wheat varieties; Floret development; Photosynthate distribution; Grain number per ear
2017-06-01
河南省科技開放合作項目(172106000044)。
文祥朋(1981─),男,河南新鄉人,農藝師,碩士,主要從事植物營養與作物施肥推廣工作。*通訊作者:孫克剛。
S512.1
A
1001-8581(2017)08-0011-04
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
