純種發(fā)酵對小米淀粉分子結(jié)構(gòu)及老化特性的影響
2017-09-03 10:06:34葛云飛寧冬雪夏甜天王維浩曹龍奎
食品科學(xué) 2017年16期
關(guān)鍵詞:質(zhì)量
寇 芳,葛云飛,沈 蒙,寧冬雪,夏甜天,王維浩,2,曹龍奎,2,*
純種發(fā)酵對小米淀粉分子結(jié)構(gòu)及老化特性的影響
寇 芳1,葛云飛1,沈 蒙1,寧冬雪1,夏甜天1,王維浩1,2,曹龍奎1,2,*
(1.黑龍江八一農(nóng)墾大學(xué)食品學(xué)院,黑龍江 大慶 163319;2.國家雜糧工程技術(shù)研究中心,黑龍江 大慶 163319)
利用自然發(fā)酵液中對淀粉老化特性起主要作用的菌種發(fā)酵,研究發(fā)酵對小米淀粉分子結(jié)構(gòu)及老化特性的影響。采用0.2 g/100 mL的NaOH溶液提取發(fā)酵后的小米淀粉,研究不同菌種發(fā)酵后對小米淀粉顆粒特性、官能團(tuán)、分子質(zhì)量、糊化及老化特性的影響。結(jié)果表明:發(fā)酵未改變淀粉的偏光十字;植物乳桿菌、戊糖片球菌和屎腸球菌發(fā)酵后小米淀粉表面被侵蝕,釀酒酵母及芽孢菌發(fā)酵后淀粉顆粒表面侵蝕跡象變重,孔道加深且數(shù)量增多;釀酒酵母及芽孢菌發(fā)酵后小米淀粉官能團(tuán)區(qū)的峰位未變,但特征峰強(qiáng)度減弱,植物乳桿菌、戊糖片球菌及屎腸球菌發(fā)酵后小米淀粉指紋區(qū)圖譜部分消失;植物乳桿菌發(fā)酵后Ⅰ區(qū)、Ⅱ區(qū)的重均、數(shù)均分子質(zhì)量較小米淀粉降低。戊糖片球菌、屎腸球菌、釀酒酵母發(fā)酵后Ⅰ區(qū)的重均分子質(zhì)量升高,數(shù)均分子質(zhì)量降低,Ⅱ區(qū)重均、數(shù)均分子質(zhì)量降低。芽孢菌發(fā)酵后Ⅰ區(qū)的數(shù)均分子質(zhì)量略有升高,Ⅱ區(qū)重均、數(shù)均分子質(zhì)量降低。植物乳桿菌、戊糖片球菌及屎腸球菌發(fā)酵后淀粉的糊化溫度、回生值及最終黏度降低,熱焓值升高。釀酒酵母發(fā)酵后糊化溫度及回生值降低,最終黏度及熱焓值升高。發(fā)酵使淀粉的分子結(jié)構(gòu)、支鏈淀粉及直鏈淀粉分子發(fā)生改變,短期抗老化性能提高。
純種發(fā)酵;淀粉;分子結(jié)構(gòu);老化
隨著社會經(jīng)濟(jì)及加工水平的迅速發(fā)展,谷物精細(xì)化加工造成的富貴病越來越受到人們的重視,因此如何進(jìn)行膳食搭配是當(dāng)今社會關(guān)注的焦點(diǎn)[1]。小米的營養(yǎng)價值高,易被人體消化吸收,是我國孕婦產(chǎn)后滋補(bǔ)的主要能量來源,具有清熱、滋陰、補(bǔ)脾腎、清腸胃、利小便、治水瀉等功效[2],是一種藥食兩用的雜糧。雖然小米有悠久的歷史,但我國關(guān)于小米食品的開發(fā)還處于初開發(fā)階段[3],因此開發(fā)以小米為主、精面為輔的雜糧食品來改善人們的膳食結(jié)構(gòu)將有廣闊的發(fā)展前景。
發(fā)酵可顯著提高小米淀粉及蛋白質(zhì)的消化率,提高營養(yǎng)價值,改善口感及風(fēng)味[4],且發(fā)酵后淀粉的回生值顯著降低,有利于改善淀粉老化特性,但發(fā)酵小米的生產(chǎn)條件難控制,難以實(shí)現(xiàn)產(chǎn)業(yè)化生產(chǎn),且國內(nèi)外學(xué)者對發(fā)酵小米的研究主要集中在發(fā)酵產(chǎn)品的研發(fā)[5-7]、發(fā)酵小米制品[8-13]、菌種的分離鑒定[14]、發(fā)酵特定產(chǎn)品的微生物特性[15-20]等,而關(guān)于小米自然發(fā)酵及發(fā)酵對淀粉改性的研究鮮有報道。若能掌握自然發(fā)酵對淀粉老化特性的改性機(jī)理,將對生產(chǎn)抗老化小米開拓新途徑。本實(shí)驗(yàn)以黑龍江省廣泛銷售的紅谷小米為研究對象,利用16S rDNA、26S rDNA鑒定對淀粉老化特性起主要作用的菌種發(fā)酵,研究發(fā)酵過程中小米淀粉分子結(jié)構(gòu)及老化特性的變化,旨在為探究自然發(fā)酵對淀粉老化特性的改性機(jī)理及不同菌株對小米淀粉性質(zhì)的影響提供理論支持,為開發(fā)發(fā)酵小米新途徑提供依據(jù)。
1 材料與方法
1.1 材料與試劑
菌種為實(shí)驗(yàn)室利用16S rDNA、26S rDNA鑒定技術(shù)從自然發(fā)酵液中篩選并鑒定所得;紅谷小米購置于大慶。
氫氧化鈉(分析純) 天津市大茂化學(xué)試劑廠;鹽酸 廣州萬從化工有限公司;葡聚糖標(biāo)準(zhǔn)品(Dextran T-20000(mw2 000 000 g/mol)、DextranT-150(mw133 800 g/mol)、DextranT-4099(mw36 800 g/mol)、DextranT-10(mw9 700 g/mol)、Dextran T-5(mw2 700 g/mol))美國Sigma公司;蒸餾水 實(shí)驗(yàn)室自制。
1.2 儀器與設(shè)備
DSC1型差示掃描量熱儀、AR2140型分析天平 瑞士梅特勒-托利多儀器有限公司;DK-S24型恒溫水浴鍋、DGG-9053A型電熱鼓風(fēng)干燥箱 上海森信實(shí)驗(yàn)儀器有限公司;TGL16B型臺式離心機(jī) 上海安亭科學(xué)儀器廠;MJ-10A型磨粉機(jī) 上海市浦恒信息科技有限公司;掃描電子顯微鏡 荷蘭FEI公司;Nicolet 6700傅里葉變換紅外光譜儀 美國Thermo Fisher公司;1525高效液相色譜儀(配2410示差折光檢測器和Empower工作站)美國Waters公司;高壓不銹鋼坩堝 上海瑾恒儀器有限公司;壓樣機(jī) 美國Perkin-Elmer公司;RVA4500型快速黏度分析儀、RVA專用鋁盒 瑞典波通儀器公司。
1.3 方法
1.3.1 預(yù)處理
1.3.1.1 菌種制備
將自然條件下小米發(fā)酵液中菌株分離純化,利用16S rDNA、26S rDNA對分離菌種鑒定,結(jié)果為:乳酸菌(戊糖片球菌、屎腸球菌、植物乳桿菌)、芽孢菌和釀酒酵母,將鑒定后的菌種擴(kuò)大培養(yǎng)。具體方法為:乳酸菌、芽孢菌活化后接種至肉湯液體培養(yǎng)基,37 ℃增殖培養(yǎng)24~48 h,直到菌懸液濃度達(dá)到5×108CFU/mL。釀酒酵母活化后,接種到酵母浸出粉胨葡萄糖(yeast extract peptone dextrose,yPD)液體培養(yǎng)基,28 ℃增殖培養(yǎng)24~48 h,直到酵母菌菌懸液濃度達(dá)到5×108CFU/mL。
1.3.1.2 純種發(fā)酵小米
用滅菌蒸餾水將小米清洗一遍后置于500 mL燒杯中,加入無菌蒸餾水(小米與無菌水的比例為1∶1.2(g/mL))。分別吸取擴(kuò)大培養(yǎng)后的微生物菌懸液(戊糖片球菌、屎腸球菌、植物乳桿菌、芽孢菌、釀酒酵母)5 mL接種到小米中,用保鮮膜密封,在最適溫度(乳酸菌37 ℃,酵母菌28 ℃,芽孢菌37 ℃)條件下培養(yǎng)120 h進(jìn)行發(fā)酵。
1.3.1.3 小米淀粉的制備
將發(fā)酵(未發(fā)酵)小米干磨法過80目篩,得到的小米粉按料液比1∶3(g/mL)置于0.2 g/100 mL的NaOH溶液中,提取3 h,3 000 r/min離心10 min,棄去上清液,除去沉淀區(qū)中上層黃褐色的物質(zhì),水洗,連續(xù)離心4 次,直至淀粉漿呈白色。用1 mol/L HCl溶液調(diào)漿至pH 7.0中性,離心,30 ℃干燥,過80 目篩,即得發(fā)酵(未發(fā)酵)小米淀粉。
1.3.2 發(fā)酵對小米淀粉結(jié)構(gòu)及老化特性的影響
1.3.2.1 發(fā)酵對小米淀粉偏光十字的影響
配成1%的淀粉懸液,混勻使淀粉顆粒充分分散,滴1滴于潔凈的載玻片上,置于偏光顯微鏡的載物臺上觀察不同發(fā)酵處理的小米淀粉,并拍攝其照片。
1.3.2.2 發(fā)酵對小米淀粉顆粒形貌的影響
用導(dǎo)電膠將分散均勻的小米淀粉樣品固定,利用離子濺射鍍膜儀噴金,用掃描電子顯微鏡進(jìn)行觀察并拍攝有代表性的照片。
1.3.2.3 發(fā)酵對小米淀粉基團(tuán)的影響
取小米淀粉0.5~2 mg,再加入100~200 mg經(jīng)過磨細(xì)干燥的KBr粉末,混合研磨均勻后,放入傅里葉變換紅外光譜儀光束中進(jìn)行全波段的掃描,掃描范圍為4 000~400 cm-1,分辨率為4 cm-1,最后得到小米淀粉的紅外光譜圖[21]。
1.3.2.4 發(fā)酵對小米淀粉分子質(zhì)量的影響
稱取適量樣品于容量瓶中,用流動相溶解,定容。色譜條件:色譜柱:Ultra hydrogel? Linear(300 mm×7.8 mm,兩根柱子串聯(lián)),流動相:0.1 mol/L硝酸鈉,流速:0.8 mL/min,柱溫:30 ℃,進(jìn)樣量:20 μL(有時可調(diào)整至30 μL)。
1.3.2.5 發(fā)酵對小米淀粉糊化特性的影響
利用差示掃描熱量儀研究發(fā)酵后小米淀粉糊化特性的變化。準(zhǔn)確稱樣品3.0 mg于坩堝中,加入7 μL蒸餾水,用壓片器反復(fù)壓3~4 次至坩堝邊緣密封完好。室溫均衡12 h,在N2流量150 mL/min、壓力0.1 MPa、升溫速率5 ℃/min[22]的條件下測定不同樣品的糊化特性曲線。
1.3.2.6 發(fā)酵對小米淀粉老化特性的影響
利用快速黏度儀研究發(fā)酵后小米淀粉老化特性的變化。準(zhǔn)確稱取樣品3.500 g(干基)于樣品盒中,加水25 mL,35 ℃保溫3 min,以6 ℃/min的速率加熱到95 ℃,保溫5 min,以6 ℃/min的速率降溫到50 ℃。用儀器配套的軟件分析得到曲線[22]。
1.4 數(shù)據(jù)統(tǒng)計(jì)分析
采用SPSS軟件對數(shù)據(jù)統(tǒng)計(jì)分析,用Origin軟件進(jìn)行繪圖處理。
2 結(jié)果與分析
2.1 發(fā)酵對淀粉偏光十字的影響
淀粉顆粒內(nèi)部存在結(jié)晶結(jié)構(gòu)和無定形結(jié)構(gòu),淀粉分子結(jié)晶區(qū)具有高度有序的排列,而在非結(jié)晶區(qū)淀粉分子呈無序的排列,正是這兩種不同的結(jié)構(gòu)在密度和折射率上存在差別,使得淀粉顆粒產(chǎn)生各向異性的現(xiàn)象,當(dāng)偏振光通過淀粉顆粒時就會產(chǎn)生偏光十字[23]。在偏光顯微鏡下觀察淀粉顆粒呈現(xiàn)黑色的十字,將顆粒分成四個白色的區(qū)域,稱為偏光十字或馬耳他十字[24]。
淀粉顆粒通過改性后,其內(nèi)部的分子鏈有序排列的結(jié)晶結(jié)構(gòu)就會遭到破壞,偏光十字就會發(fā)生改變或消失,所以淀粉顆粒內(nèi)部偏光十字的變化在一定程度上可以反映出淀粉顆粒結(jié)晶結(jié)構(gòu)是否發(fā)生變化。不同發(fā)酵時間小米淀粉的偏光十字如圖1所示。不同菌種發(fā)酵方式所得淀粉都有明顯的偏光十字,說明乳酸菌、酵母菌及芽孢菌發(fā)酵都未使淀粉顆粒的結(jié)晶區(qū)發(fā)生改變。發(fā)酵過程中微生物代謝產(chǎn)生的有機(jī)酸和酶可能主要作用于淀粉的無定形區(qū)。

圖1 不同發(fā)酵處理小米淀粉偏光十字Fig. 1 Polarized cross of millet starch fermented by different pure cultures
2.2 發(fā)酵對淀粉顆粒結(jié)構(gòu)的影響

圖2 不同發(fā)酵處理小米淀粉顆粒形態(tài)的影響Fig. 2 Effects of different fermentation processes on the morphology of millet starch granules
小米淀粉顆粒大部分呈多角形,少數(shù)為球形[25]。由圖2可以看出,小米淀粉顆粒表面較為光滑,且有細(xì)小的孔道。利用戊糖片球菌發(fā)酵的小米淀粉表面侵蝕跡象較植物乳桿菌和屎腸球菌輕,但三者發(fā)酵后小米淀粉顆粒表面均有明顯被侵蝕的跡象,并伴有明顯的孔洞,說明乳酸菌發(fā)酵可使淀粉表面被侵蝕。芽孢菌發(fā)酵后淀粉顆粒表面被侵蝕跡象嚴(yán)重,形成較多深孔,而釀酒酵母發(fā)酵后淀粉顆粒表面的孔道數(shù)增多的同時,其孔道加深且變大,說明酵母菌發(fā)酵使淀粉顆粒被侵蝕且顆粒表面的孔道變深。發(fā)酵后淀粉顆粒表面被侵蝕,孔洞變深,且發(fā)酵后顆粒不在保持其完整表面主要是由于發(fā)酵過程中微生物產(chǎn)酸產(chǎn)酶,使淀粉顆粒無定形區(qū)遭到破壞,淀粉的顆粒特性發(fā)生改變。
2.3 發(fā)酵對小米淀粉基團(tuán)的影響
中紅外光譜分為官能團(tuán)區(qū)和指紋區(qū),其頻率范圍為4 000~1 300 cm-1及1 300~400 cm-1[26]。利用紅外光譜研究發(fā)酵前后小米淀粉分子鏈上基團(tuán)結(jié)構(gòu)及化學(xué)鍵的變化,分析發(fā)酵對小米淀粉分子結(jié)構(gòu)的影響,結(jié)果如圖3所示。小米淀粉在2 931、2 062 cm-1處有特征吸收峰,其對應(yīng)著C—H的伸縮振動峰,1 648 cm-1處為C=O振動吸收峰,且發(fā)酵后,小米淀粉官能團(tuán)區(qū)的特征吸收峰位未發(fā)生變化,但峰強(qiáng)減弱。指紋區(qū)在1 242、929、861、765 cm-1處出現(xiàn)吸收峰,分別對應(yīng)C—O、C—C伸縮振動峰和C—H面外彎曲振動吸收峰。屎腸球菌、戊糖片球菌及植物乳桿菌的樣品在1 368.38~1 154.00 cm-1及 527.54~438.01 cm-1處的吸收峰幾乎消失,而釀酒酵母及芽孢菌在該波長范圍內(nèi)其吸收峰減弱。其原因?yàn)槭耗c球菌、戊糖片球菌及植物乳桿菌發(fā)酵產(chǎn)的酸、酶等作用于小米淀粉顆粒的孔道,進(jìn)入淀粉內(nèi)部,淀粉顆粒結(jié)晶結(jié)構(gòu)遭到一定程度的損壞,使淀粉部分水解,化學(xué)鍵發(fā)生改變,指紋區(qū)的圖譜部分消失,而釀酒酵母及芽孢菌的作用較三者弱,故吸收峰減弱但并未消失。淀粉紅外光譜指紋區(qū)可以用來分析淀粉分子短程有序的結(jié)構(gòu),指紋區(qū)圖譜部分消失,說明發(fā)酵使淀粉的結(jié)構(gòu)遭到一定程度破壞。
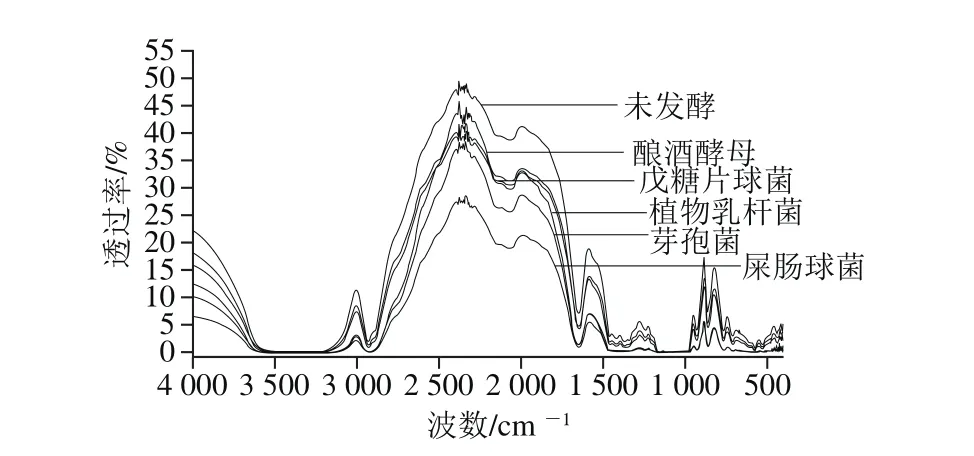
圖3 小米淀粉的紅外光譜圖Fig. 3 FTIR spectra of fermented millet starch
2.4 發(fā)酵對淀粉分子質(zhì)量的影響
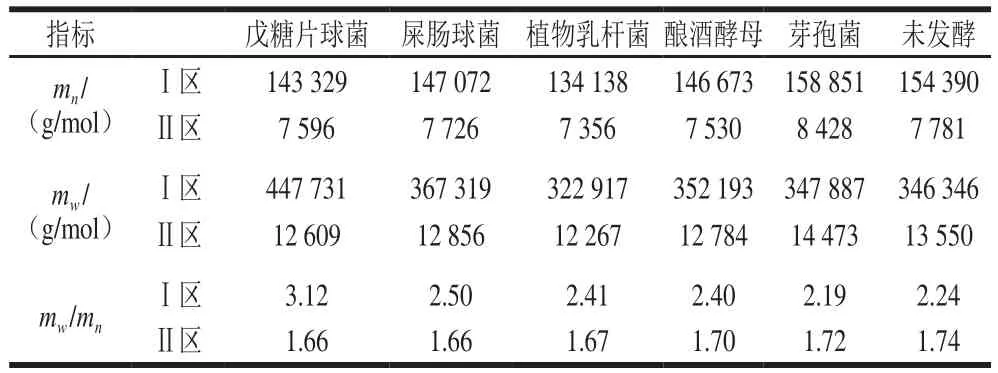
表1 發(fā)酵小米淀粉分子質(zhì)量Table 1 Molecular mass of fermented millet starch
淀粉是由分子質(zhì)量不同的高分子化合物組成的同系混合物,其凝膠特性、老化特性、糊化特性等不僅與平均分子質(zhì)量有密切的關(guān)聯(lián),還受分子質(zhì)量分布的影響[27]。淀粉的分子質(zhì)量分布是表征聚合物分子鏈長短的重要參數(shù),常用mw/mn(多分散指數(shù))表示。mw/mn越接近于1,說明樣品的組分越單一,越大則說明樣品的組分越復(fù)雜,分子分布越寬[28]。對于多分散、寬分布的聚合物其mw/mn范圍在1.5~3或者3~30之間,表1為不同處理小米淀粉分子質(zhì)量及其分布。小米淀粉分子質(zhì)量分為兩個區(qū),分別為Ⅰ區(qū)(主要為支鏈淀粉)和Ⅱ區(qū)(主要為直鏈淀粉及少部分支鏈淀粉)(圖4,20 min的峰為蛋白質(zhì)峰,本實(shí)驗(yàn)不予考慮)。植物乳桿菌發(fā)酵后重均分子質(zhì)量Ⅰ區(qū)降低23 429 g/mol、Ⅱ區(qū)降低1 283 g/mol,數(shù)均分子質(zhì)量Ⅰ區(qū)降低20 252 g/mol、Ⅱ區(qū)降低425 g/mol,說明發(fā)酵使Ⅰ區(qū)、Ⅱ區(qū)小米支鏈淀粉及直鏈淀粉發(fā)生水解,生成小分子物質(zhì)被微生物利用,分子質(zhì)量和回生值降低。而Ⅰ區(qū)戊糖片球菌、屎腸球菌、釀酒酵母的重均分子質(zhì)量分別升高101 385、20 973 g/mol和5 847 g/mol,Ⅱ區(qū)降低941、694、766 g/mol;數(shù)均分子質(zhì)量Ⅰ區(qū)分別降低11 061、7 318 g/mol和7 717 g/mol,Ⅱ區(qū)分別降低185、55、251 g/mol,說明發(fā)酵使Ⅰ區(qū)短支鏈淀粉水解,而較長支鏈淀粉的比例相對增加,起到純化支鏈淀粉的作用,同時發(fā)酵使Ⅱ區(qū)直鏈淀粉水解,淀粉的回生值降低。芽孢菌Ⅰ區(qū)的數(shù)均分子質(zhì)量略有升高,說明發(fā)酵使小米淀粉支鏈部分水解為短支鏈及部分直鏈淀粉,故發(fā)酵后Ⅱ區(qū)的直鏈淀粉重均、數(shù)均分子質(zhì)量升高,淀粉回生值略有升高。除芽孢菌外,發(fā)酵后Ⅰ區(qū)淀粉的分子質(zhì)量分布mw/mn均略高于小米淀粉,說明發(fā)酵使Ⅰ區(qū)小米淀粉分子鏈的長短發(fā)生了變化,淀粉的組分變復(fù)雜,而Ⅱ區(qū)小米淀粉的分子質(zhì)量分布變狹窄,說明發(fā)酵使Ⅱ區(qū)直鏈淀粉組分變得單一,起到純化淀粉的作用,同時,淀粉的回生值降低,起到抗老化的作用。
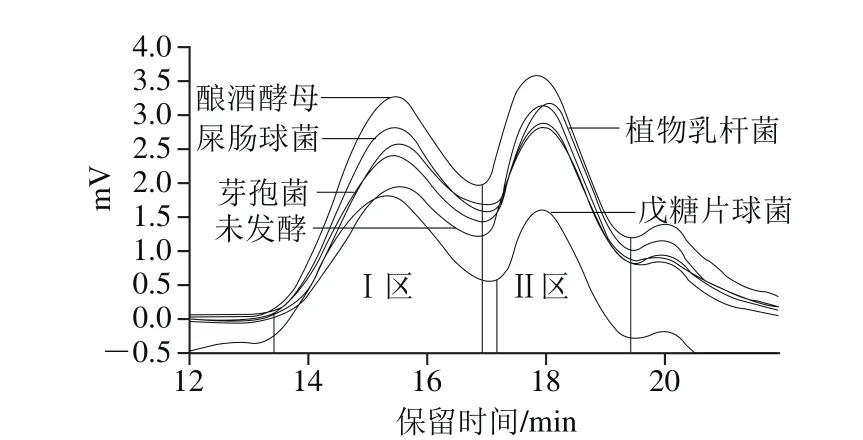
圖4 小米淀粉分子質(zhì)量Fig. 4 Molecular mass of millet starch
2.5 發(fā)酵對淀粉糊化特性的影響
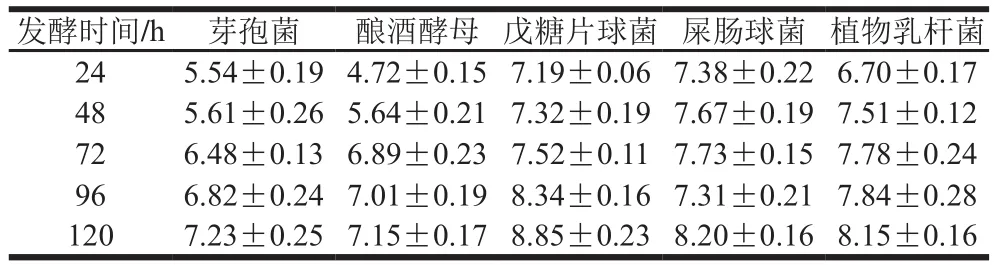
表2 不同發(fā)酵處理小米淀粉熱焓值Table 2 Enthalpy value of fermented millet starch J/g

表3 不同發(fā)酵處理小米淀粉糊化溫度Table 3 Gelatinization temperature of millet starch fermented for different times℃
如表2所示,乳酸菌發(fā)酵小米淀粉的熱焓值高于釀酒酵母及芽孢菌,并均隨發(fā)酵時間的延長熱焓值逐漸升高,且發(fā)酵所得淀粉的熱焓值高于小米淀粉。戊糖片球菌的熱焓值高于其他兩株乳酸菌,而釀酒酵母和芽孢菌的熱焓值相差不大。如表3所示,乳酸菌、釀酒酵母和芽孢菌發(fā)酵的糊化溫度均隨發(fā)酵時間延長逐漸降低并略低于未發(fā)酵小米淀粉。糊化溫度的降低是由于發(fā)酵過程中微生物代謝所產(chǎn)生的酸、酶作用于淀粉顆粒的無定形區(qū),使淀粉無定形區(qū)的結(jié)構(gòu)遭到了破壞,淀粉顆粒結(jié)合水的能力增強(qiáng),使淀粉更易糊化。另外,發(fā)酵也降低了小米淀粉蛋白和脂肪的含量,使其與淀粉的絡(luò)合能力減弱[29-30],故淀粉的糊化溫度降低。熱焓值上升是由于其發(fā)酵產(chǎn)生大量的有機(jī)酸及酶使較長支鏈淀粉水解為大量的中間級及較短鏈的支鏈淀粉,使結(jié)晶區(qū)的比例相對增大的緣故。
2.6 發(fā)酵對小米淀粉老化特性的影響
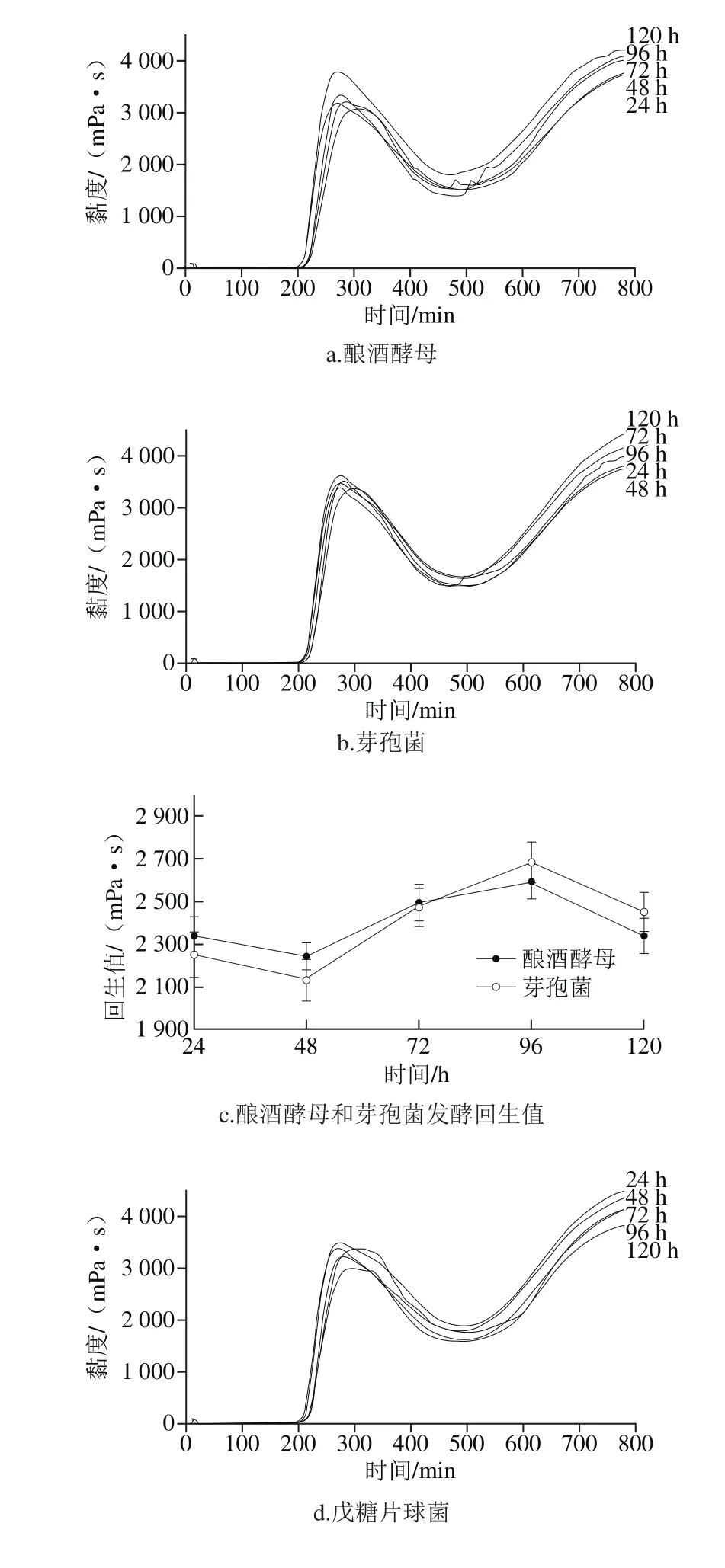
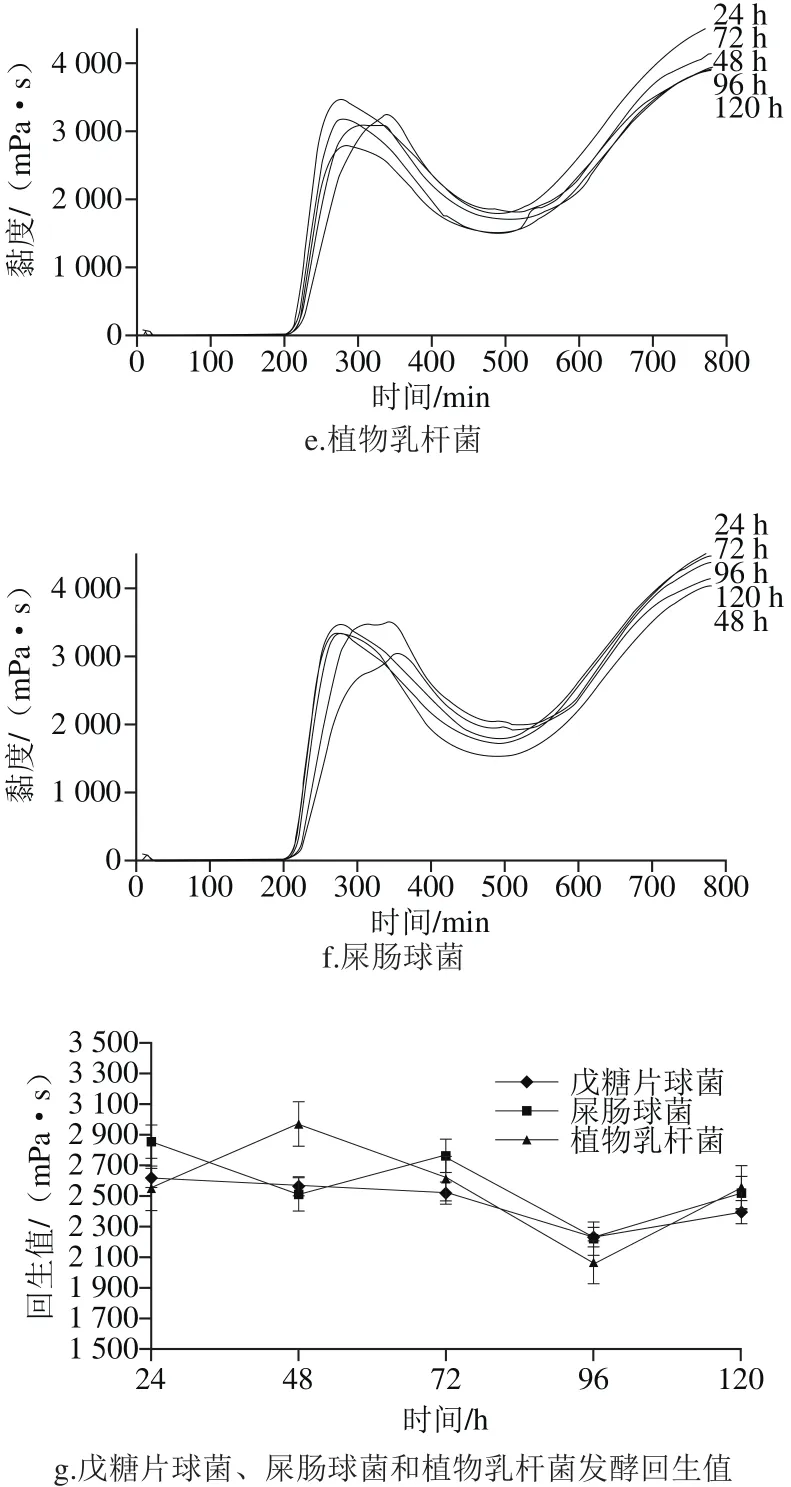
圖5 老化特性的測定結(jié)果Fig. 5 Retrogradation characteristics of fermented millet starch
如圖5a、b所示,釀酒酵母發(fā)酵所得小米淀粉的最終黏度隨發(fā)酵時間逐漸升高,發(fā)酵120 h時其最終黏度達(dá)到最大值。而芽孢菌的最終黏度則出現(xiàn)不規(guī)律的變化。圖5c所示為釀酒酵母、芽孢菌發(fā)酵后小米淀粉回生值的變化,可見發(fā)酵進(jìn)行48 h時小米淀粉的回生值最低,96 h后回生值略有下降,回生值隨發(fā)酵時間的變化可以解釋為發(fā)酵初期,發(fā)酵菌株代謝產(chǎn)生淀粉酶,使淀粉顆粒無定形區(qū)的顆粒結(jié)構(gòu)遭到破壞,直鏈淀粉水解為小分子物質(zhì)被微生物利用,回生值降低。隨發(fā)酵時間的延長,淀粉酶等通過淀粉顆粒表面的孔道及無定形區(qū)破壞后的孔洞進(jìn)入淀粉顆粒結(jié)晶區(qū),使結(jié)晶區(qū)外側(cè)部分結(jié)構(gòu)較為疏松的支鏈淀粉水解為短支鏈及大部分直鏈淀粉,直鏈淀粉的溶出加速了淀粉分子的締合,導(dǎo)致48 h后淀粉的回生值升高。由于結(jié)晶區(qū)內(nèi)部支鏈淀粉的結(jié)構(gòu)致密,不易被酸、酶水解,淀粉酶無法繼續(xù)水解支鏈淀粉生成更多的直鏈淀粉,而溶出的直鏈淀粉在淀粉酶的作用下繼續(xù)被水解為小分子物質(zhì),故96 h后淀粉回生值略有降低。
如圖5d、e、f所示,乳酸菌發(fā)酵后小米淀粉的最終黏度隨發(fā)酵時間延長逐漸降低,戊糖片球菌和植物乳桿菌在發(fā)酵120 h達(dá)到最低。由圖5g可以看出,植物乳桿菌在發(fā)酵48 h回生值升高,屎腸球菌在72 h淀粉的回生值升高,而戊糖片球菌隨發(fā)酵的進(jìn)行回生值逐漸降低,且植物乳桿菌在發(fā)酵后期淀粉的回生值顯著低于其他兩株菌,這可能與發(fā)酵菌株特性有關(guān)。結(jié)合掃描電子顯微鏡和凝膠滲透色譜可知,乳酸菌發(fā)酵后小米淀粉回生值降低一方面是由于發(fā)酵過程中乳酸菌代謝產(chǎn)生大量的有機(jī)酸作用于淀粉顆粒表面,使淀粉顆粒不在保持完整的顆粒,同時淀粉顆粒無定形區(qū)的直鏈淀粉發(fā)生水解,另一方面,有機(jī)酸通過淀粉顆粒表面的孔道及無定形區(qū)破壞后的孔洞進(jìn)入淀粉顆粒結(jié)晶區(qū),使結(jié)晶區(qū)部分結(jié)構(gòu)較為疏松的支鏈淀粉水解為更短鏈及小分子物質(zhì)被微生物利用,導(dǎo)致發(fā)酵后回生值顯著低于小米淀粉,淀粉顆粒短期抗老化性能提高。

表4 不同發(fā)酵淀粉回生值Table 4 Retrogradation values of fermented millet starch
由表4可知,除戊糖片球菌與屎腸球菌發(fā)酵后小米淀粉回生值差異不顯著,其余均差異顯著。乳酸菌、釀酒酵母發(fā)酵后小米淀粉的回生值顯著降低,而芽孢菌發(fā)酵后小米淀粉的回生值略高,說明乳酸菌和酵母菌發(fā)酵可能是自然發(fā)酵對小米淀粉回生值起改性作用的主要菌種,且釀酒酵母的存在使發(fā)酵液有柔和的酒香味,賦予小米制品特有的風(fēng)味。
3 結(jié) 論
利用從自然發(fā)酵液中篩選出的乳酸菌(戊糖片球菌、屎腸球菌、植物乳桿菌)、酵母菌(釀酒酵母)及芽孢菌發(fā)酵小米后,其淀粉的顆粒形態(tài)被破壞,分子結(jié)構(gòu)發(fā)生變化,淀粉短期抗老化性能提高。戊糖片球菌、屎腸球菌、植物乳桿菌發(fā)酵后小米淀粉顆粒表面侵蝕較重,指紋區(qū)的化學(xué)鍵發(fā)生變化,圖譜部分消失,導(dǎo)致重均、數(shù)均分子質(zhì)量發(fā)生改變,此外小米淀粉分子結(jié)構(gòu)的變化使其回生值、最終黏度、糊化溫度降低,熱焓值升高,適用于焙烤類等混合粉的添加。而釀酒酵母發(fā)酵后淀粉顆粒表面孔道數(shù)增多,指紋區(qū)圖譜強(qiáng)度減弱,結(jié)晶區(qū)分子質(zhì)量略有升高,無定形區(qū)分子質(zhì)量均降低,說明發(fā)酵后部分短支鏈淀粉、直鏈淀粉水解為小分子物質(zhì),回生值、糊化溫度降低,熱焓值升高。芽孢菌發(fā)酵后淀粉顆粒表面形成較多深孔,指紋區(qū)圖譜峰強(qiáng)減弱,結(jié)晶區(qū)分子質(zhì)量變化較小,無定形區(qū)直鏈淀粉含量增加,淀粉老化性能較上述菌種略有增加,抗老化能力弱。此研究從發(fā)酵后淀粉顆粒特性及分子結(jié)構(gòu)的角度闡明了乳酸菌(戊糖片球菌、屎腸球菌、植物乳桿菌)、酵母菌(釀酒酵母)及芽孢菌對小米淀粉老化特性的改性機(jī)理,為分析小米自然發(fā)酵抗老化的改性機(jī)理奠定了基礎(chǔ)及數(shù)據(jù)支持,為開發(fā)發(fā)酵小米新途徑、生產(chǎn)小米發(fā)酵劑提供依據(jù)。
參考文獻(xiàn):
[1] 張超, 張暉, 李冀新. 小米的營養(yǎng)以及應(yīng)用研究進(jìn)展[J]. 中國糧油學(xué)報, 2007(1): 51-55; 78.
[2] 王海濱, 夏建新. 小米的營養(yǎng)成分及產(chǎn)品研究開發(fā)進(jìn)展[J]. 糧食科技與經(jīng)濟(jì), 2010(4): 36-38; 46.
[3] 王勇. 小米的營養(yǎng)價值及內(nèi)蒙古小米生產(chǎn)加工現(xiàn)狀[D]. 呼和浩特:內(nèi)蒙古大學(xué), 2010.
[4] 袁美蘭, 魯戰(zhàn)會, 程永強(qiáng), 等. 自然發(fā)酵對米粉RVA黏度性質(zhì)及米粉拉伸性質(zhì)的影響[J]. 中國糧油學(xué)報, 2008, 23(1): 6-9.
[5] 張秀媛, 何擴(kuò), 石后, 等. 響應(yīng)面法優(yōu)化小米紅曲制備工藝及其抗腫瘤活性研究[J]. 中國糧油學(xué)報, 2015, 30(3): 19-22; 34.
[6] ISSOUFOU A. 發(fā)酵小米多肽的抗氧化與抗菌活性的研究[D]. 無錫:江南大學(xué), 2014.
[7] FAROOQ U. 小米膳食纖維作為主要碳源對益生菌生長和發(fā)酵過程中短鏈脂肪酸產(chǎn)量的影響研究[D]. 無錫: 江南大學(xué), 2013.
[8] 李喜仙, 劉璽. 小米液態(tài)發(fā)酵生產(chǎn)紅曲色素技術(shù)研究[J]. 食品科學(xué), 2003, 24(1): 83-86. DOI:10.3321/j.issn:1002-6630.2003.01.020.
[9] SERRAINO M R, THOMPSON L U, SOVOIE L, et al. Effect of phytic acid on the in vitro rate of digestibility of rapeseed protein and amino acids[J]. Journal of Food Science, 1985, 50(5): 1689-1692.
[10] yOON J H, THOMPSON L U, JENKINS D J A. The effect of phytic acid on in vitro rate of starch digestibility and dlood glucose response[J]. The American Journal of Clinical Nutrition, 1983, 38(6): 835-842.
[11] OJOKOH A O, FAyEMI O E, OCLOON F C K, et al. Effect of fermentation on proximate composition, physicochemical and microbial characteristics of pearl millet (Pennisetum glaucum (L.) R. Br.) and acha (Digitaria exilis (Kippist) Stapf) flour blends[J]. Academic Journals, 2015, 7(1): 1-8. DOI:10.5897/JABSD2014.0236.
[12] KOUAKOU B, ALEXIS K K S, ADJéHI D, et al. Biochemical changes occurring during germination and fermentation of millet and effect of technological processes on starch hydrolysis by the crude enzymatic extract of millet[J]. Journal of Applied Sciences Research, 2008(11): 1502-1510.
[13] OUATTARA L T S, RIVIER C M, VERNIERE C, et al. Enzyme activities of lactic acid bacteria from a pearl millet fermented grauel (ben-saalga) of functional interest in nutrition[J]. International Journal of Food Microbiology, 2008, 128(2): 395-400.
[14] ILANGO S, ANTONy U. Assessment of the microbiological quality of koozh, a fermented millet beverage[J]. African Journal of Microbiology Research, 2014, 8(15): 308-312.
[15] OUATTARA C A T. Isolation and identification of lactic acid and non-acid lactic bacteria from “dèguè” of western africa traditional fermented millet-based food[J]. African Journal of Microbiology Research, 2015, 9(36): 2001-2005. DOI:10.5897/AJMR2015.7548.
[16] LEI V, FRIIS H, MICHARLSEN K F, et al. Spontaneously fermented millet product as a natural probiotic treatment for diarrhoea in young children: an intervention study in Northern Ghana[J]. International Journal of Food Microbiology, 2006, 110(3): 246-253. DOI:10.1016/ j.ijfoodmicro.2006.04.022.
[17] OH y J, JUNG D S. Evaluation of probiotic properties of lactobacillus and pediococcus strains isolated from omegisool, a traditionally fermented millet alcoholic beverage in Korea[J]. LWT-Food Science and Technology, 2015, 63: 437-444.
[18] OLUWAJOBA S O, AKINyOSOyE F A, OyETAyO O V. Comparative sensory and proximate evaluation of spontaneously fermenting kunu-zaki made from germinated and ungerminated composite cereal grains[J]. Food Science & Nutrition, 2013, 1(4): 336-349.
[19] ZAKARI U M, HASSAN A, ABBO E S. Physico-chemical and sensory properties of “Agidi” from pearl-millet (Pennisetum glaucum) and bambara groundnut (Vigna subterranean) flour blends[J]. African Journal of Food Science, 2010, 4(10): 662-667.
[20] KHETARPAUL N, CHAUHAN B M. Fermentation of pearl millet flour with yeasts and lactobacilli: in vitro digestibility and utilisation of fermented flour for weaning mixtures[J]. Plant Foods for Human Nutrition, 1990, 40(3): 167-173.
[21] REDDy C K, SURIyA M, VIDyA P V, et al. Effect of γ-irradiation on structure and physico-chemical properties of Amorphophallus paeoniifolius starch[J]. International Journal of Biological Macromolecules, 2015, 79(2): 309-315. DOI:10.1016/ j.ijbiomac.2015.05.003.
[22] 冷雪, 曹龍奎. 利用差示掃描量熱儀研究小米淀粉及小米粉的糊化特性[J]. 食品科學(xué), 2015, 36(19): 60-66. DOI:10.7506/spkx1002-6630-201519011.
[23] 李堅(jiān)斌, 溫雪馨, 李琳, 等. 超聲場對馬鈴薯淀粉顆粒形貌與結(jié)晶結(jié)構(gòu)的影響[J]. 農(nóng)業(yè)工程學(xué)報, 2008, 24(4): 284-286.
[24] 劉東莉. 不同鏈/支比玉米淀粉顆粒結(jié)構(gòu)原位表征與分析[D]. 杭州:浙江大學(xué), 2014.
[25] 李玥, 鐘芳. 大米淀粉糊化過程的光譜分析[J]. 高分子學(xué)報, 2008(7): 720-725.
[26] 蒲華寅. 等離子體作用對淀粉結(jié)構(gòu)及性質(zhì)影響的研究[D]. 廣州: 華南理工大學(xué), 2013.
[27] 許永亮, 程科, 趙思明, 等. 大米淀粉的分子質(zhì)量分布及其與粘性的相關(guān)性研究[J]. 中國農(nóng)業(yè)科學(xué), 2007(3): 566-572.
[28] 張攀峰, 陳玲, 李曉璽, 等. 不同直鏈/支鏈比的玉米淀粉分子質(zhì)量及其構(gòu)象[J]. 食品科學(xué), 2010, 31(19): 157-160.
[29] 閔偉紅, 李里特, 王朝輝. 乳酸菌發(fā)酵對大米淀粉物理化學(xué)性質(zhì)的影響[J]. 食品科學(xué), 2004, 25(10): 73-76. DOI:10.3321/ j.issn:1002-6630.2004.10.013.
[30] 魯戰(zhàn)會. 生物發(fā)酵米粉的淀粉改性及凝膠機(jī)理研究[D]. 北京: 中國農(nóng)業(yè)大學(xué), 2002.
Effects of Pure Culture Fermentation on Molecular Structure and Retrogradation Characteristics of Millet Starch
KOU Fang1, GE Yunfei1, SHEN Meng1, NING Dongxue1, XIA Tiantian1, WANG Weihao1,2, CAO Longkui1,2,*
(1. College of Food Science, Heilongjiang Bayi Agricultural University, Daqing 163319, China; 2. National Cereals Engineering Technology Research Center, Daqing 163319, China)
In this study, we examined the effects of fermentation by pure cultures of Lactobacillus plantarum, Pediococcus pentosaceus, Enterococcus faecium, Saccharomyces cerevisiae and Bacillus, which play an important role in affecting the retrogradation properties of naturally fermented starch, on the molecular structure and retrogradation characteristics of millet starch with the aim of providing a theoretical basis and data support for elucidating the mechanism by which natural fermentation and pure culture fermentation improve the retrogradation properties of millet starch, and new ways of developing fermented millet products. Millet starch was extracted from fermented millet with 0.2 g/100 mL NaOH and evaluated for granular characteristics, functional groups, molecular mass and, pasting and retrogradation properties. The results obtained were as follows. Fermentation did not changed the cross polarization characteristics of starch. The surface of millet starch granules fermented by Lactobacillus plantarum, Pediococcus pentosaceus and Enterococcus faecium was eroded, but the surface of starch granules after fermentation by Saccharomyces cerevisiae and Bacillus was eroded more seriously, with more deeper channels. The fermentation by Saccharomyces cerevisiae and Bacillus did not change the peak positions in the functional region, but reduced the intensity of the characteristic peaks. The fingerprint region of millet starch fermented by Lactobacillus plantarum, Pediococcus pentosaceus and Enterococcus faecium partially disappeared, The weight average and number average molecular masses in regions Ⅰ and Ⅱ were decreased after fermentation by Lactobacillus plantarum. In region Ⅰ, the weight average molecular mass after fermentation by Pediococcus pentosaceus, Enterococcus faecium and Saccharomyces cerevisiae was increased, and the average molecular weight was decreased, while in region Ⅱ, the weight average and number average molecular masses were both decreased. After fermentation by Bacillus, the number average molecular mass in region Ⅰ was slightly increased, but the weight average molecular weight was slightly decreased; both the weight average and number average molecular weight in region Ⅱ were decreased.The gelatinization temperature, final viscosity and retrogradation value were decreased, but the enthalpy was increased after fermentation by Lactobacillus plantarum, Pediococcus pentosaceus and Enterococcus faecium. The gelatinization temperature and retrogradation value were decreased, the final viscosity and enthalpy were increased after fermentation by Saccharomyces cerevisiae. In conclusion, fermentation could change the molecular structure of starch, branched chain starch and amylose, and improve the short-term anti-retrogradation performance.
pure culture fermentation; starch; molecular structure; retrogradation
10.7506/spkx1002-6630-201716014
TS231
A
1002-6630(2017)16-0092-07
2016-11-23
黑龍江八一農(nóng)墾大學(xué)研究生創(chuàng)新科研項(xiàng)目(YJSCX2017-Y54)
寇芳(1993—),女,碩士研究生,研究方向?yàn)殡s糧發(fā)酵。E-mail:18249556388@163.com
*通信作者:曹龍奎(1965—),男,教授,博士,研究方向?yàn)檗r(nóng)產(chǎn)品加工。E-mail:caolongkui2013@163.com
寇芳, 葛云飛, 沈蒙, 等. 純種發(fā)酵對小米淀粉分子結(jié)構(gòu)及老化特性的影響[J]. 食品科學(xué), 2017, 38(16): 92-98. DOI:10.7506/spkx1002-6630-201716014. http://www.spkx.net.cn
KOU Fang, GE Yunfei, SHEN Meng, et al. Effects of pure culture fermentation on molecular structure and retrogradation characteristics of millet starch[J]. Food Science, 2017, 38(16): 92-98. (in Chinese with English abstract)
10.7506/ spkx1002-6630-201716014. http://www.spkx.net.cn
猜你喜歡
中學(xué)生數(shù)理化·中考版(2022年10期)2022-11-10 09:37:42
中學(xué)生數(shù)理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學(xué)生數(shù)理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學(xué)生數(shù)理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(shè)(2018年6期)2018-04-22 03:16:54
產(chǎn)品可靠性報告(2017年7期)2017-09-05 09:49:12
中學(xué)生數(shù)理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
