無芒隱子草種子發(fā)育過程中不同位置葉片的光合速率及重量與種子產(chǎn)量的相關(guān)性
2017-09-13 07:18:34陶奇波白夢杰孫秋瑾韓云華王彥榮
草地學(xué)報 2017年3期
關(guān)鍵詞:產(chǎn)量
陶奇波, 白夢杰, 孫秋瑾, 韓云華, 王彥榮
(草地農(nóng)業(yè)生態(tài)系統(tǒng)國家重點實驗室 蘭州大學(xué)草地農(nóng)業(yè)科技學(xué)院, 甘肅 蘭州 730020)
無芒隱子草(Cleistogenessongorica)為禾本科隱子草屬植物,是我國西北地區(qū)重要的野生鄉(xiāng)土草種[1]。其具有抗旱、耐寒、適口性好等特性,不僅可以作為優(yōu)良的牧草,而且在城市綠化、荒漠化治理與恢復(fù)中具有廣闊的應(yīng)用前景[2]。當前對無芒隱子草的研究主要集中在建植技術(shù)[3]、種子萌發(fā)生態(tài)學(xué)[4-6]、圍欄封育[7-8]等方面。就其種子生產(chǎn)方面的研究則僅限于栽培密度以及施氮肥兩個方面[9-10]。
光合作用是植物完成整個生活史的原初動力,也是植物生長發(fā)育的基礎(chǔ)和生產(chǎn)力高低的決定性因素[11-12]。然而,目前關(guān)于無芒隱子草光合生理方面的研究尚未見報道。大量在農(nóng)作物上的研究表明,植株不同位葉的光合特性以及生物量存在顯著差異,對種子產(chǎn)量的貢獻也不同[13-14]。紅花(Carthamustinctorius)不同位葉的光合特性存在顯著差異,其上位葉凈光合速率與單株花產(chǎn)量呈極顯著正相關(guān)[15]。羅俊[16]等研究表明能源甘蔗(Saccharumofficinarum)分蘗期第3葉的葉寬和水分利用效率以及第5葉的葉寬對產(chǎn)量影響較大。本文通過在無芒隱子草種子發(fā)育各時期測定其生殖枝自上而下第1~7葉的凈光合速率以及重量,闡明種子發(fā)育各時期不同位置葉片的凈光合速率、葉片重量以及含水量、穗重量的變化趨勢,并探究以上指標與種子產(chǎn)量的相關(guān)性,以期為無芒隱子草種子生產(chǎn)提供更加具有針對性的技術(shù)指導(dǎo)。
1 材料與方法
1.1 研究區(qū)概況
試驗地位于蘭州大學(xué)草地農(nóng)業(yè)科技學(xué)院民勤試驗站(38°44′N, 103°1′E),海拔1 307 m,年均溫為8.3℃,年平均降水量127.7 mm,蒸發(fā)量2 623 mm,無霜期162 d,年日照時數(shù)約為3 073 h。土壤為沙壤土,0~30 cm土壤速效氮、磷、鉀含量分別為31.08, 17.48, 118.36 mg·kg-1,全氮含量為7.76 g·kg-1,有機質(zhì)含量為12.19 g·kg-1,土壤pH值為8.01。
1.2 試驗方法與測定指標
于2016年7月,在行距30 cm、播種后第3年且長勢均衡的無芒隱子草種子生產(chǎn)田中隨機選取保留了完整7枚葉片的生殖枝為試驗材料,并自上而下將7枚葉片分別編為1~7號。
1.2.1不同位葉凈光合速率及光合貢獻率 在盛花期、灌漿期、乳熟期、蠟熟期以及完熟期隨機選取10個生殖枝,于晴朗無風的上午,用LI-6400便攜式光合儀,在自然光照下測定每個生殖枝自上而下第1~7葉的凈光合速率Pn(net photosynthetic rate),每個葉片測定3個數(shù)值。并按照如下公式計算不同位葉的光合貢獻率:
光合貢獻率(%)=(該葉片凈光合速率/第1~7葉總光合速率)×100%
1.2.2不同位葉重量、重量比例以及含水量 在盛花期、灌漿期、乳熟期、蠟熟期以及完熟期隨機選取10個生殖枝,按照葉片位置順序剪取各個生殖枝的葉片,稱鮮重后分別裝袋,于80oC烘至恒重,用萬分之一天平稱干重,并按照如下公式計算葉重量比例及葉片含水量:
葉重量比例=(該葉重量/第1~7葉總重量)×100%
葉片含水量=[ (鮮重-干重)/鮮重]×100%
1.2.3種子發(fā)育過程中小穗重量 在盛花期、灌漿期、乳熟期、蠟熟期以及完熟期隨機選取10個生殖枝,手工摘除其小穗,于80℃烘至恒重,用萬分之一天平稱干重。
1.2.4種子產(chǎn)量 在完熟期隨機選取10個生殖枝,手工采種,風干至種子含水量為8%左右后,用萬分之一天平稱重,即為單個生殖枝的種子產(chǎn)量。
1.3 數(shù)據(jù)分析
用SPSS 19.0統(tǒng)計軟件對各指標進行方差分析,并對各指標與種子產(chǎn)量進行相關(guān)分析。
2 結(jié)果與分析
2.1 無芒隱子草種子發(fā)育過程中各位葉凈光合速率
相同位葉在各時期的凈光合速率有顯著差異(P<0.05)(表1)。各位葉的凈光合速率隨種子發(fā)育總體呈下降趨勢。其中,灌漿期第1~6葉顯著高于其他時期,而第7葉的最大凈光合速率出現(xiàn)在盛花期。除第7葉的凈光合速率隨種子發(fā)育一直呈顯著下降外,其余各位葉在盛花期至乳熟期變異不大,而自乳熟期以后則隨種子發(fā)育顯著降低。同一時期不同位葉的凈光合速率同樣存在顯著差異(P<0.05)(表1)。在相同的種子發(fā)育期內(nèi),同一生殖枝上的各位葉自上而下總體呈先增加后降低的趨勢。在盛花期,第2~5葉顯著高于其他位置葉片;灌漿期第3~4葉顯著高于其他葉片,第1,6葉顯著高于第7葉;乳熟期2~5葉相對較高,隨后隨生殖枝部位降低呈現(xiàn)顯著下降;蠟熟期與完熟期第2~3葉凈光合速率顯著高于其他位葉,其后亦隨生殖枝部位降低而下降。在各時期,第1,6,7葉凈光合速率均顯著小于其他位葉。
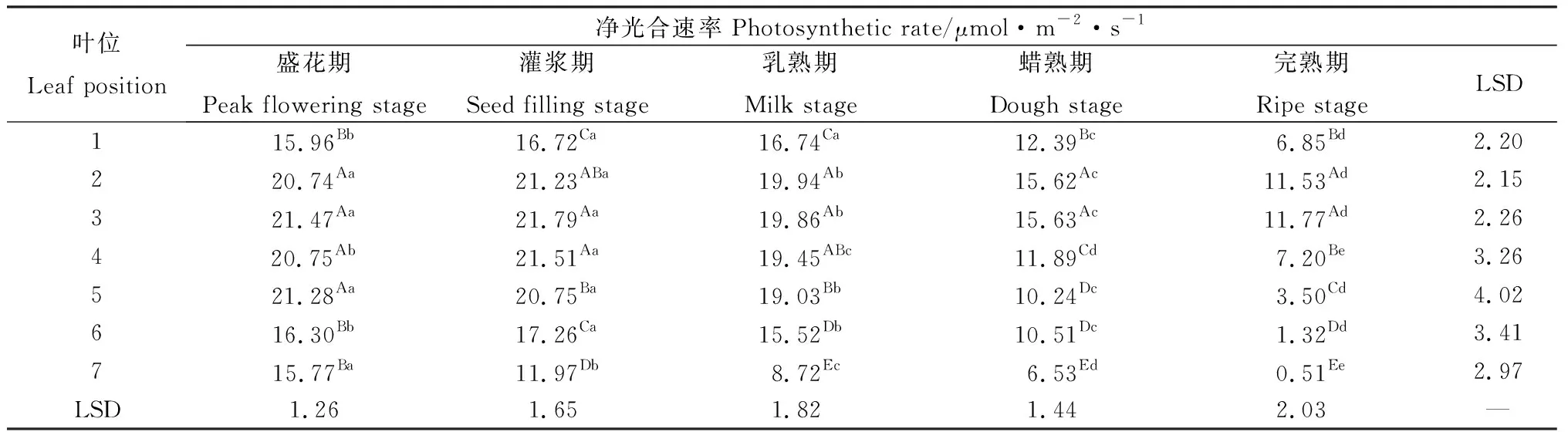
表1 無芒隱子草種子發(fā)育過程中各位葉凈光合速率Table 1 Photosynthetic rate of C. songorica leaves in different position during seed development
注:同列不同大寫字母表示不同位葉間差異顯著(P<0.05),同行不同小寫字母表示不同生育期間差異顯著(P<0.05),下同
Note:Different capital letters in the same column indicate significant difference between positions at the 0.05 level, different lowercase letters in the same row indicate significant difference between stages at the 0.05 level. The same below
2.2 無芒隱子草種子發(fā)育過程中各位葉光合貢獻率
相同位置葉片隨著種子發(fā)育其光合貢獻率表現(xiàn)出較大的波動(表2)。第1~2葉隨種子發(fā)育始終呈現(xiàn)顯著的上升趨勢(P<0.05),而第3~4葉在乳熟期之前光合貢獻率變化不大,之后則顯著上升。在完熟期,第1~4葉的凈光合速率占據(jù)了整個生殖枝葉片凈光合速率的87.55%。第5~6葉的光合貢獻率在乳熟期前差異不明顯,隨后則表現(xiàn)出顯著的降低趨勢。第7葉除在蠟熟期有所增加外,均隨種子發(fā)育而顯著下降。在同一時期,各位置葉片的凈光合速率亦有顯著差異(P<0.05)(表2)。同一生殖枝上的各位葉自上而下表現(xiàn)出先增加后降低的規(guī)律。盛花期第3葉與第5葉顯著高于其他葉片;在灌漿期與乳熟期,第2~5葉顯著高于其他位葉;蠟熟期與完熟期第2~3葉的光合貢獻率占有絕對優(yōu)勢,其余葉片則隨位置下降而顯著降低。
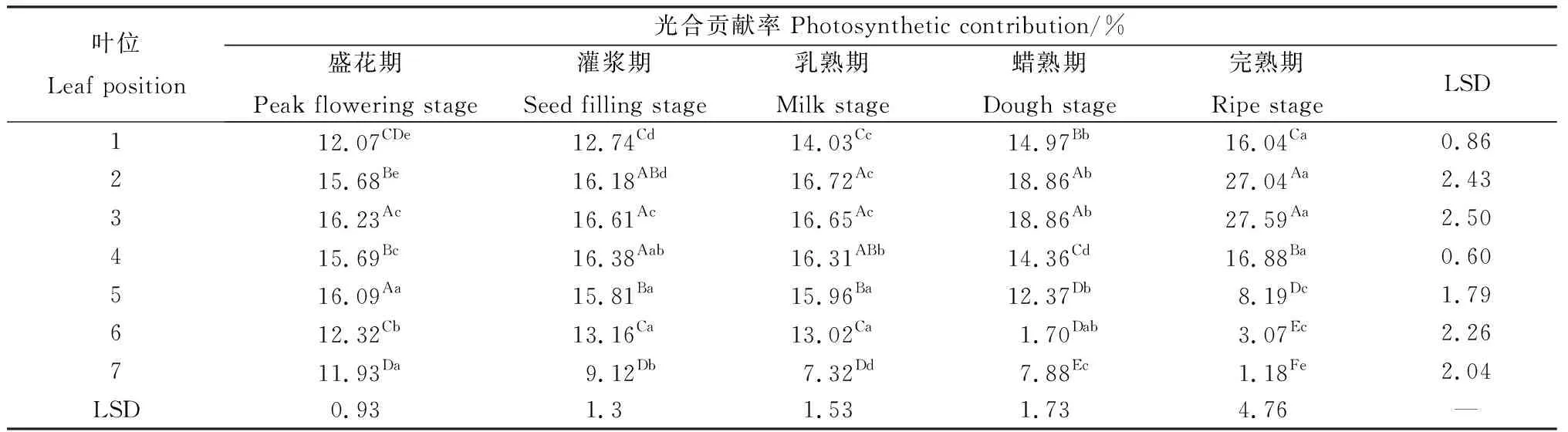
表2 無芒隱子草種子發(fā)育過程中各位葉光合貢獻率Table 2 Photosynthetic contribution rate of C. songorica leaves in different position during seed development
2.3 無芒隱子草種子發(fā)育過程中各位葉重量
無芒隱子草生殖枝上相同位葉的重量在各發(fā)育期差異顯著(P<0.05)(表3)。其中,第1~2葉在種子發(fā)育過程中表現(xiàn)出先增加后降低的趨勢,自盛花期至灌漿期顯著增加,而后則顯著降低。第3~7葉始終隨著種子發(fā)育呈現(xiàn)出降低的態(tài)勢,其中,第3~5葉從盛花期至蠟熟期顯著降低。第6~7葉則從盛花期至乳熟期顯著降低,此后無顯著變化。在同一時期內(nèi),不同位葉重量同樣有顯著差異(P<0.05)(表3)。在各時期,各位葉自上而下表現(xiàn)出先增加后降低的趨勢,第4葉的重量在各時期均顯著高于其他位葉,各個時期第5至第7葉均連續(xù)顯著降低。多重比較表明,盛花期、灌漿期、乳熟期及完熟期1,2葉之間的重量無顯著差異,蠟熟期2,3葉間無顯著差異。
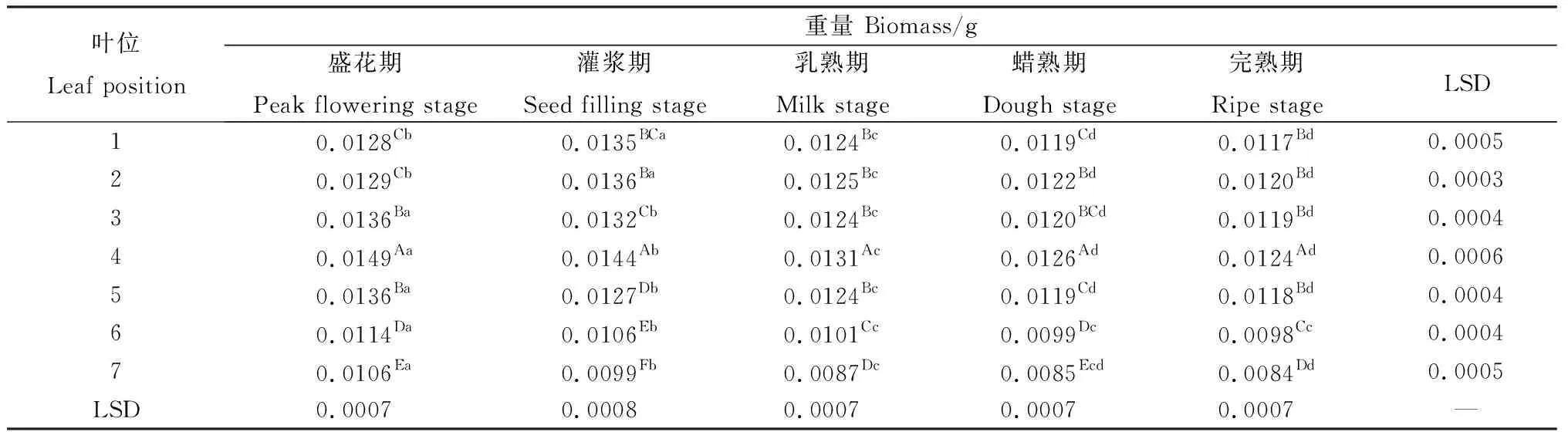
表3 無芒隱子草種子發(fā)育過程中各位葉重量Table 3 Biomass of C. songorica leaves in different position during seed development
2.4 無芒隱子草種子發(fā)育過程中各位葉重量比例
相同位葉在各個生育期的葉重量比例存在顯著差異(P<0.05)(表4)。前3枚葉片隨種子發(fā)育有上升趨勢,第1~2葉盛花期葉重量比例顯著小于其他時期;第3葉在整個種子發(fā)育期略有上升,但是差異不明顯。第4,7葉在盛花期至乳熟期顯著降低,之后無顯著變化;第5~6葉除盛花期至灌漿期外,在各個時期差異均不顯著。在同一生育期內(nèi),各位葉的葉重量比例變化顯著(P<0.05)(表4)。各時期第4位葉的葉重量比例顯著高于其他位葉,同時第6~7葉顯著低于其他位葉,表明中部葉片構(gòu)成了無芒隱子草生殖枝葉片重量的主要部分。
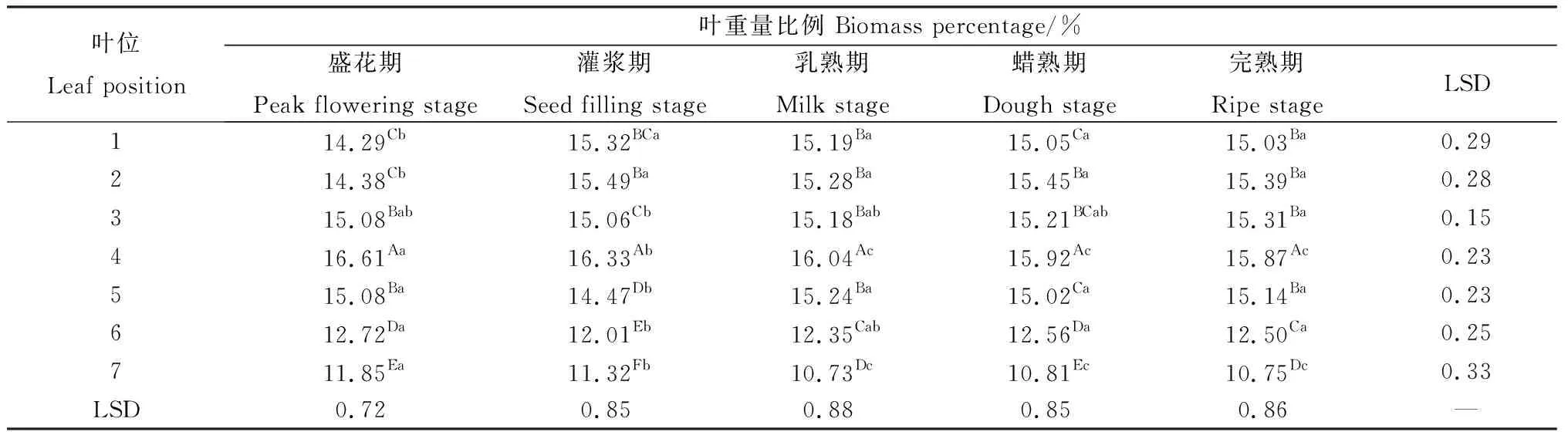
表4 無芒隱子草種子發(fā)育過程中各位葉重量比例Table 4 Biomass percentage of C. songorica leaves in different position during seed development
2.5 無芒隱子草種子發(fā)育過程中各位葉含水量
同一位葉在各個生育期的葉片含水量呈現(xiàn)出類似規(guī)律(表5)。第1~4葉以及第7葉在盛花期與灌漿期葉片含水量差異不顯著,在灌漿期以后則持續(xù)顯著降低(P<0.05)。第5葉在整個種子發(fā)育期內(nèi)含水量均顯著降低,第6葉的最大含水量出現(xiàn)在灌漿期,自灌漿期以后則顯著降低。同一時期不同位葉的葉片含水量表現(xiàn)出自上而下逐漸降低的趨勢(表5),各時期第1葉含水量均顯著高于其他位葉,而第6~7葉含水量均顯著低于其他位置葉片,其他葉片之間普遍表現(xiàn)出隨位置降低而顯著下降的趨勢,其中盛花期與灌漿期2~3葉間含水量無顯著差異,蠟熟期3~4葉間含水量無顯著差異。
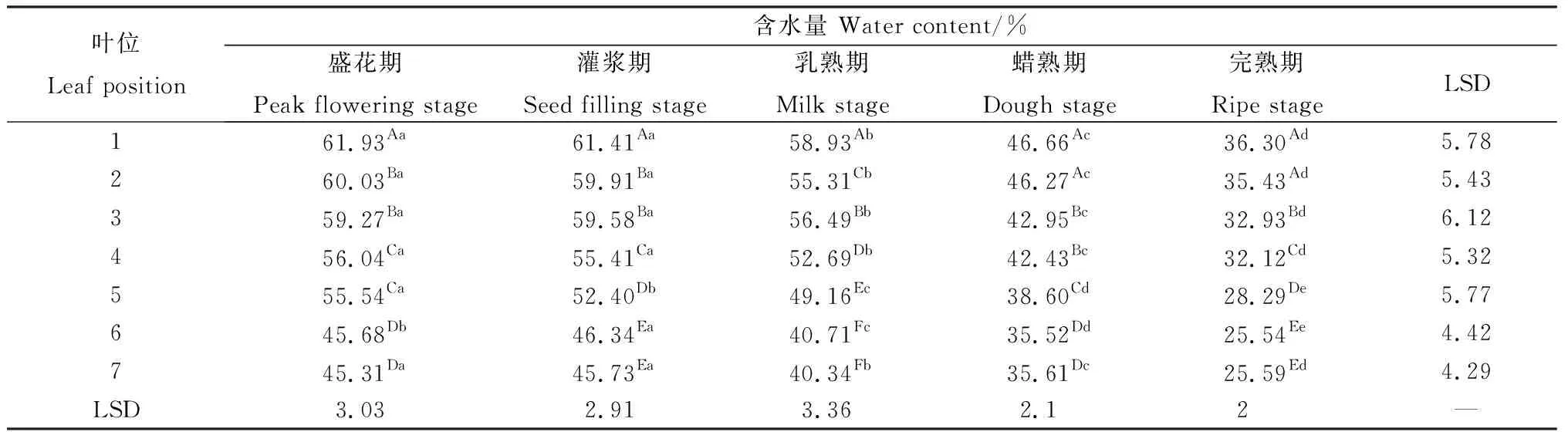
表5 無芒隱子草種子發(fā)育過程中各位葉含水量Table 5 Water content of C. songorica leaves in different position during seed development
2.6 葉片位置與時期對光合特性及生物量影響的雙因素方差分析
通過雙因子方差分析,可以看出葉位、種子發(fā)育期及其交互作用對凈光合速率、葉生物量以及含水量均有極顯著影響。種子發(fā)育期對光合貢獻率及生物量比例無顯著影響。葉位及種子發(fā)育期的交互作用對各個指標均有極顯著影響 (表6)。
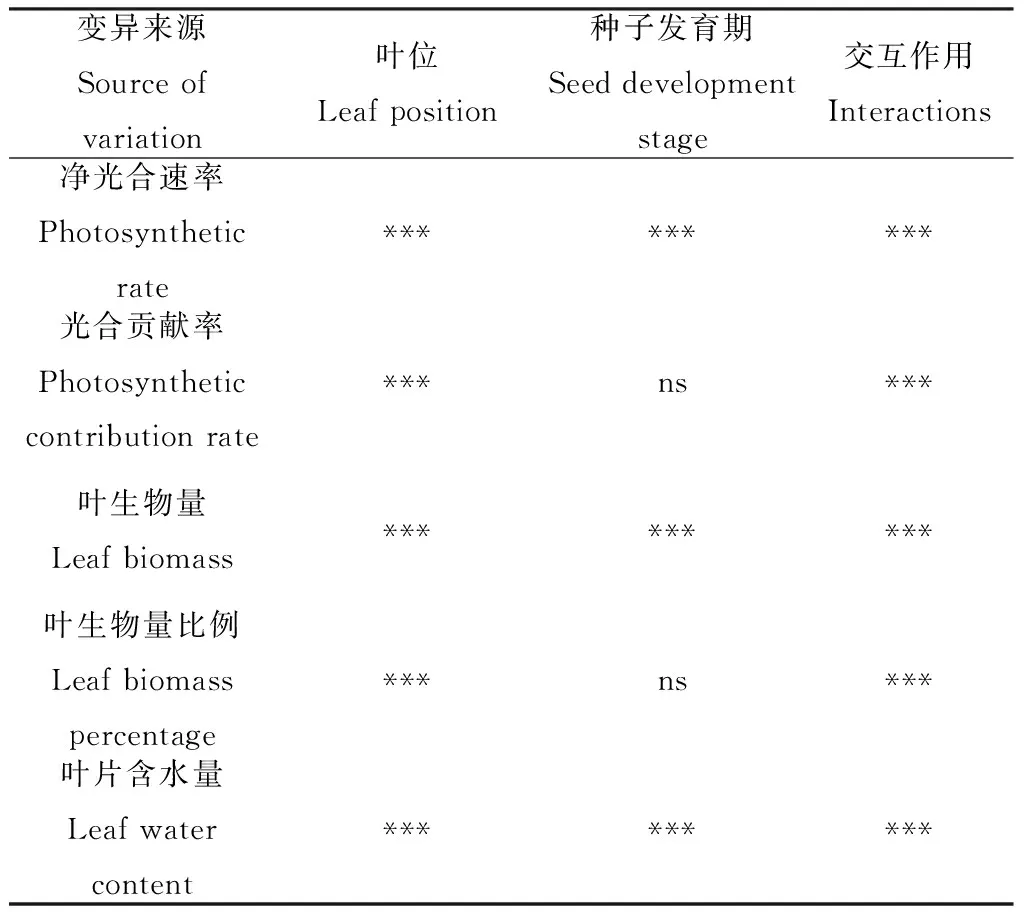
表6 葉位與種子發(fā)育期的雙因素方差分析Table 6 Two-way ANOVA of leaf position, seed development stage and their interactions
注:***表示在0.001水平顯著;ns:不顯著
Note:***indicate significant at the 0.001 level;ns:Not significant at the 0.05 level
2.7 無芒隱子草種子發(fā)育過程中葉總生物量與穗生物量變化
隨著無芒隱子草種子的發(fā)育,生殖枝上的葉總重量逐漸降低,其中盛花期與灌漿期間無顯著差異,灌漿期與乳熟期間無顯著差異,其余時期間則差異顯著(P<0.05)。穗重量則隨著種子發(fā)育持續(xù)顯著增加(P<0.05)(表7),完熟期穗重量較盛花期增加了161%。
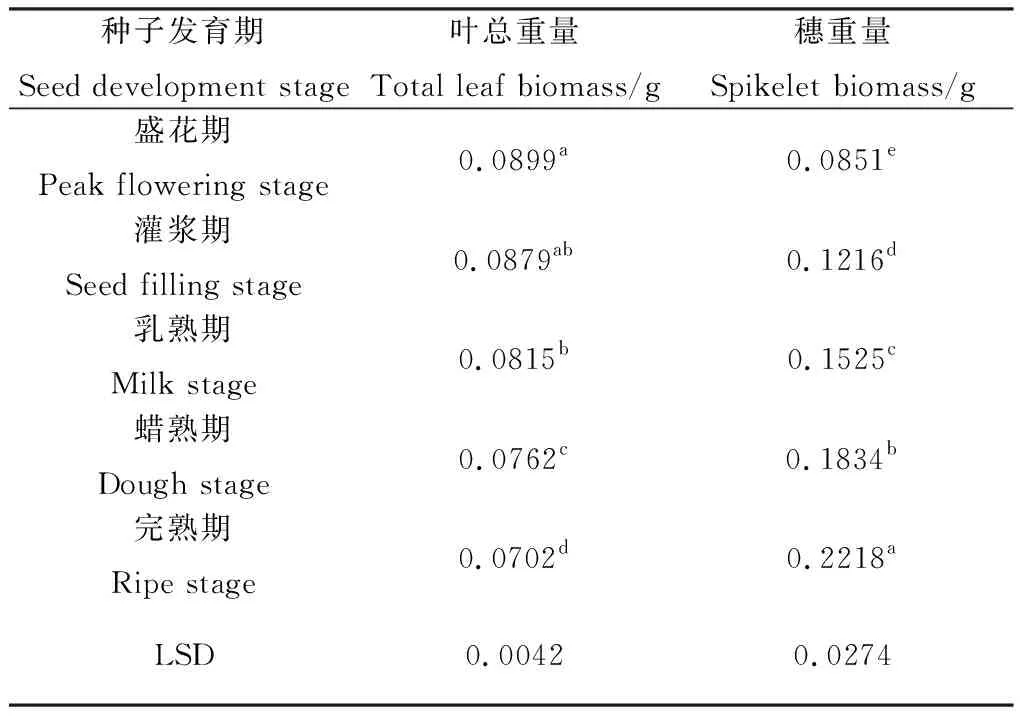
表7 無芒隱子草種子發(fā)育過程中葉總重量與穗重量Table 7 Total leaf biomass and spikelet biomass of C. songorica during seed development
2.8 無芒隱子草生殖枝種子產(chǎn)量與不同時期各位葉凈光合速率、重量、穗生物量相關(guān)分析
各個位葉光合、形態(tài)參數(shù)與種子產(chǎn)量的相關(guān)性如表8所示。盛花期第1~6葉的凈光合速率與重量均與種子產(chǎn)量呈正相關(guān),其中第1,2,4葉的凈光合速率,第1,3葉的重量與種子產(chǎn)量存在極顯著正相關(guān)(P<0.01),第3,5葉凈光合速率與第2,4,5,6葉重量與種子產(chǎn)量存在顯著正相關(guān)(P<0.05),盛花期葉總重量與種子產(chǎn)量亦表現(xiàn)出極顯著正相關(guān)關(guān)系,穗總重量則與種子產(chǎn)量呈現(xiàn)負相關(guān)。灌漿期第1~5葉的凈光合速率、重量、葉總重量以及穗重量與種子產(chǎn)量之間表現(xiàn)出正相關(guān),然而第6~7葉的凈光合速率及重量與種子產(chǎn)量存在負相關(guān)關(guān)系,尤其是第7葉的凈光合速率及重量分別與種子產(chǎn)量存在顯著負相關(guān)與極顯著負相關(guān)。乳熟期第1~6葉凈光合速率及第1~5葉重量與種子產(chǎn)量呈正相關(guān),其他位葉的凈光合速率與重量則與種子產(chǎn)量呈負相關(guān),乳熟期葉總重量與種子產(chǎn)量存在負相關(guān)關(guān)系,而穗重量則與種子產(chǎn)量呈顯著正相關(guān)。蠟熟期2~7葉的凈光合速率與種子產(chǎn)量存在負相關(guān)關(guān)系,第2,5,7葉重量亦與種子產(chǎn)量呈負相關(guān),葉總重量與種子產(chǎn)量呈現(xiàn)顯著負相關(guān),穗重量則與種子產(chǎn)量呈現(xiàn)極顯著正相關(guān)。完熟期第1~7葉凈光合速率、第2~4與第6~7葉重量與種子產(chǎn)量均呈負相關(guān),尤其以第3,7葉凈光合速率與種子產(chǎn)量表現(xiàn)出極顯著負相關(guān),葉總重量與種子產(chǎn)量表現(xiàn)出顯著負相關(guān),穗重量則與種子產(chǎn)量呈極顯著正相關(guān)。
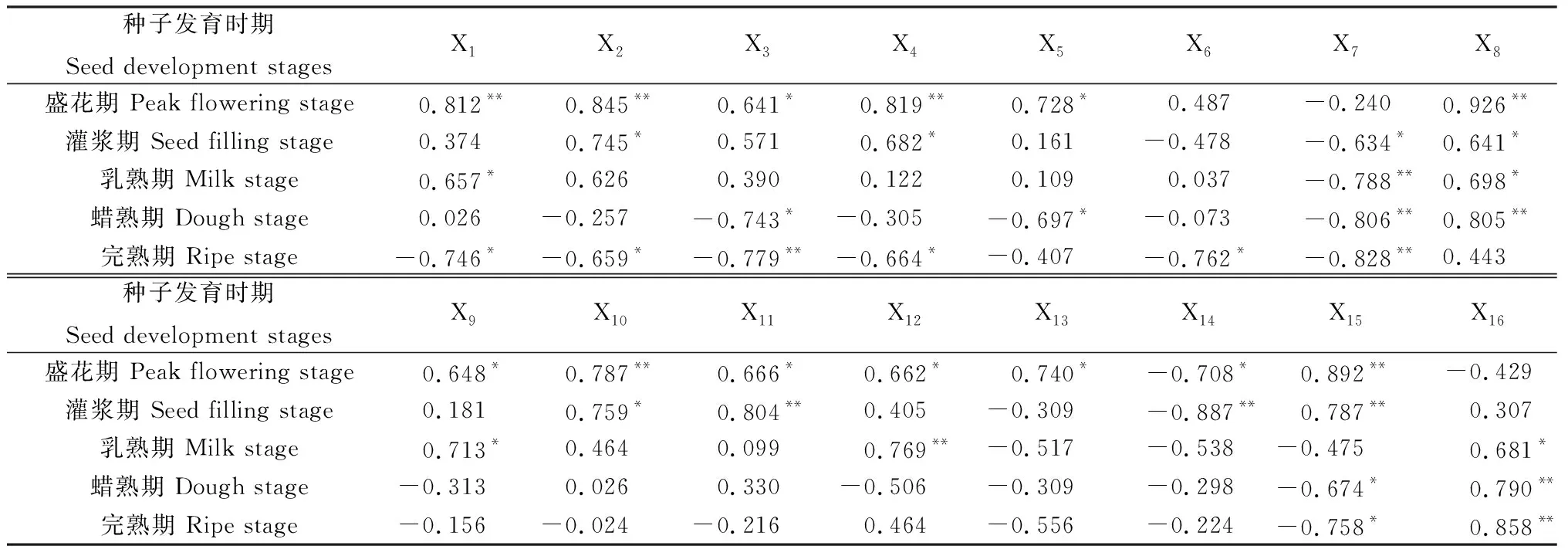
表8 無芒隱子草種子發(fā)育過程中各位葉光合速率及重量與種子產(chǎn)量相關(guān)分析Table 8 Correlation analysis between seed yield and photosynthetic rate,biomass of C. songorica leaves in different position during seed development
注:X1~X7為第1~7葉凈光合速率,X8~X14為第1~7葉重量,X15為葉總重量,X16為穗重量,*表示顯著相關(guān)(P<0.05),**表示極顯著相關(guān)(P<0.01)
Note:X1~X7indicates photosynthetic rate from the first leaf to the seventh leaf, X8~X14indicates biomass from the first leaf to the seventh leaf, X15indicates total leaf biomass, X16indicates spikelet biomass, * indicates significant correlation at the 0.05 level, ** indicates extremely significant correlation at the 0.01 level
3 討論與結(jié)論
3.1 無芒隱子草種子發(fā)育過程中各位葉凈光合速率與光合貢獻率變化
光合作用是產(chǎn)量形成的原動力,本研究說明在不同生育期,各位葉凈光合速率與光合貢獻率均有顯著差異,并且同一位葉在各個時期的凈光合速率與光合貢獻率也有明顯變化。在各時期,第2~5葉的光合貢獻率始終占據(jù)絕對優(yōu)勢。李偉[17]等研究認為黃瓜(Cucumissativus)幼苗第3~4凈光合速率最大,而第1,5葉較低。玉米(Zeamays)抽雄后不同位葉光合強度由高到低依次為中部>上部>下部[18],與本試驗結(jié)果一致。類似的結(jié)果亦可見前人在向日葵(Helianthusannuus)[19]、生姜(Zingiberofficinale)[20]以及砂仁(Amomumvillosum)[21]等作物上的研究。韓清芳[22]等研究了紫花苜蓿(Medicagosativa)不同位葉凈光合速率日變化,指出其日平均值依次為上位葉>中位葉>下位葉。水稻(Oryzasativa)抽穗期自上而下的4枚葉片凈光合速率亦呈遞減趨勢[23]。各位葉的凈光合速率主要受到葉片自身因素以及環(huán)境因子兩方面影響。玉米上位葉葉綠體基質(zhì)含量顯著高于下位葉[24],且葉綠素含量亦隨著葉位上升而增加[25],這被認為是造成不同位葉間光合特性差異的主要內(nèi)部因素。從外部環(huán)境因子考慮,凈光合速率主要受光合有效輻射(PAR)的影響[26],上位葉處在較好的光照與通風環(huán)境中,因此凈光合速率普遍大于下位葉。本文著重探討無芒隱子草不同位葉光合特性與種子產(chǎn)量的相關(guān)性,因此僅測定了種子發(fā)育過程中各位葉凈光合速率,關(guān)于不同位葉的葉綠體超微結(jié)構(gòu)、葉綠素含量以及生理生態(tài)因子的影響等則有待于進一步研究。
3.2 無芒隱子草種子發(fā)育過程中各位葉重量、重量比例、含水量以及生殖枝穗重量變化
任何作物的生長都可分為營養(yǎng)生長與生殖生長兩個階段,兩階段是對立統(tǒng)一的。營養(yǎng)生長是生殖生長的基礎(chǔ),其積累和儲藏的物質(zhì),在生殖生長階段流向植物生殖器官,促進產(chǎn)量形成;另一方面,過度的營養(yǎng)生長則會抑制生殖生長,造成減產(chǎn)。小麥(Triticumaestivum)、水稻等作物上的研究表明,種子產(chǎn)量最終取決于源、庫容和物質(zhì)運輸能力[27]。在本試驗中,無芒隱子草各位葉隨著種子發(fā)育,重量均顯著降低,其中第1~2葉最大重量出現(xiàn)于灌漿期,其他葉片最大重量出現(xiàn)在盛花期,同時,葉總重量的最大值出現(xiàn)在盛花期及灌漿期,這表明在盛花期以及灌漿期之后,葉片作為源器官,將其儲存物質(zhì)輸送給了種子以及穗等庫器官,在本試驗中,生殖枝上穗的重量隨著種子發(fā)育不斷增加,亦印證了該觀點。在蠟熟期至完熟期這一個階段,各位葉重量雖略微下降,但沒有顯著差異,表明葉片對種子產(chǎn)量的貢獻主要發(fā)生在盛花期至乳熟期這一階段,自乳熟期以后,種子獲得的儲存物質(zhì)或許主要由小穗提供,分析發(fā)現(xiàn)自乳熟期后小穗重量與種子產(chǎn)量始終呈顯著正相關(guān),也在一定程度上證明了該結(jié)論。
3.3 無芒隱子草種子發(fā)育過程中各位葉凈光合速率、重量變化與種子產(chǎn)量相關(guān)性
不同位葉在種子形成過程中起到的作用是不同的。盛花期前6位葉的凈光合速率和重量均與種子產(chǎn)量呈正相關(guān),在灌漿期與乳熟期,前5葉的凈光合速率和重量亦與種子產(chǎn)量正相關(guān),而在蠟熟期,僅有第1葉的凈光合速率、第1, 3, 4葉重量與種子產(chǎn)量呈正相關(guān),至完熟期,則僅有第1葉重量與種子產(chǎn)量呈正相關(guān)。在圍繞光合特性與作物產(chǎn)量的研究中,不同學(xué)者在不同作物上的研究結(jié)果并不一致,玉米上位葉光合速率和貢獻率顯著高于下位葉,從而對成粒率、籽粒充實度、千粒重和產(chǎn)量起決定性作用[25]。然而亦有研究表明光合速率與產(chǎn)量相關(guān)性很小[28],收獲指數(shù)增加與光合周期增長才是產(chǎn)量提高的主因。在本試驗中,前5枚葉片的凈光合速率與重量在乳熟期以前始終對種子產(chǎn)量起到積極作用,而第6位葉凈光合速率與重量僅僅在盛花期與種子產(chǎn)量表現(xiàn)出正相關(guān),第7位葉在種子發(fā)育過程中則始終與種子產(chǎn)量負相關(guān)。因此,上位葉即1~5位的葉片是與種子產(chǎn)量密切相關(guān)的,尤其是在盛花期至乳熟期這一階段,此結(jié)論與于洪蘭[29]等人在水稻上的研究結(jié)果一致。在河西走廊地區(qū),無芒隱子草盛花期至乳熟期,對應(yīng)7月下旬至8月下旬這一時間段[9],正是該地區(qū)褐斑病(Rhizoctoniasolani)、白粉病(Blumeriagraminis)等田間病害最為盛行的時期[30],在該時段,采取適宜的田間管理措施控制病蟲害,保證葉片健康完整是種子高產(chǎn)的基礎(chǔ)。同時,采取密度調(diào)控、施肥、灌溉等管理措施,設(shè)法增大葉片光合與形態(tài)參數(shù),也會對種子產(chǎn)量產(chǎn)生積極影響。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(chǎn)(2021年10期)2021-12-05 16:31:48
今日農(nóng)業(yè)(2021年14期)2021-11-25 23:57:29
今日農(nóng)業(yè)(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(zhì)(2021年1期)2021-02-22 14:14:44
今日農(nóng)業(yè)(2020年20期)2020-11-26 06:09:10
中國果業(yè)信息(2019年10期)2019-11-13 01:21:34
中國化肥信息(2019年2期)2019-01-18 15:24:35
中國化肥信息(2019年1期)2019-01-17 21:31:12
中國化肥信息(2019年4期)2019-01-17 18:47:06
