導入OsVTE 基因提高普那菊苣的抗旱性
2017-09-13 05:18:30穆志新張麗君周建萍
草地學報 2017年4期
穆志新, 師 穎, 張麗君, 周建萍
(山西省農業科學院農作物品種資源研究所 農業部黃土高原作物基因資源與種質創制重點實驗室雜糧種質資源發掘與遺傳改良山西省重點實驗室, 山西 太原 030031)
生育酚是一類重要的脂溶性抗氧化劑,在高等植物及藍細菌光合器官中合成[1],是動物和人類維持生長、繁殖和保證健康所需要的一種重要的營養物質,具有重要的代謝功能和抗氧化作用,統稱為維生素E(VE)[2-3]。植物在生長過程中會遭遇干旱、高鹽、高溫等逆境脅迫,這些脅迫會使其體內蛋白質、葉綠素、脂類等許多重要成分發生氧化,并導致細胞死亡。生育酚作為一種抗氧化劑,可以清除單線態氧[4],使活性氧自由基失活,并通過與膜脂水解產物形成復合物,清除類囊體膜上的脂質過氧化產物,阻止膜脂過氧化的擴大,從而保護膜的完整性。提高植物生育酚含量可以增加植物對干旱、高鹽、高溫等逆境的抵抗性[5].已有研究表明,油菜(BrassicacampestrisL.)中過量表達生育酚環化酶基因OsVTE可以增加油菜籽中生育酚總量,并改變菜籽油中生育酚的組成[6]。煙草(NicotianatabacumL.)植株中過量表達OsVTE可以提高植株的耐鹽性[7]。
菊苣(CichoriumintybusL)為菊科(Asteraceae)菊苣屬(Cichorium)多年生宿根草本雙子葉植物。菊苣具有藥用和食用價值,其干物質中粗蛋白含量約占30%,粗脂肪約占5.3%,粗纖維約為9.9%,無氮浸出物約為30%,并富含17種氨基酸,鈣(12.3%)、磷(0.53%)、鉀、鈉、鋅、銅等微量元素含量也較高,還含有多種維生素,更重要的是含有一些生物化學成分:馬栗樹皮素、馬栗樹皮甙、野萵苣甙、山萵苣素和山萵苣苦素等特殊成分,有抗菌、提高食欲,改善消化功能,是其他牧草植物所不具有的[8-9]。在我國主要分布在西北、華中、華北、東北地區,這些地區都比較干旱、寒冷,不利于普那菊苣的生長,因此通過基因工程手段提高普那菊苣的耐鹽能力具有明顯的應用價值,在美國該作物已被批準為商業化種植的轉基因作物[10-11]。目前已報道AtNHX1[12]、AtNHX2[13]以及APX[14]基因可以提高普那菊苣的耐鹽性和抗逆性。通過培育適合于我國西北部地區栽培的抗干旱、耐鹽堿的牧草新品種,解決牧草品種較少,品種更新慢的問題,進一步改善生態環境。本研究應用農桿菌介導法將OsVTE基因轉入普那菊苣中,獲得了轉基因植株,并對該基因對普那菊苣抗逆性的影響進行了研究,以便為提高普那菊苣的抗逆性,進而為其在生態環境較惡劣地區的推廣奠定基礎。
1 材料與方法
1.1 植物材料及供體質粒
供試植物材料為普那菊苣2 n=2 x=18,種子由山西省農業科學院畜牧獸醫研究所提供。來自水稻的OsVTE基因載體由中國科學院遺傳發育研究所803組陳受宜研究員提供,質粒名稱為pBin438- VTE;啟動子:CaMV35S,終止子:Nos。農桿菌菌株是GV3101,其菌落篩選標記基因為卡那霉素(Km)和利福平( rif )抗性基因,植物篩選標記基因為Km 抗性基因。
1.2 農桿菌介導法轉化普那菊苣及轉化再生植株的獲得
將普那菊苣的無菌苗葉片在MS 固體培養基上預培養2~3 d。用培養至對數生長期(OD600約為0.4~0.6)的上述農桿菌菌株侵染普那菊苣葉片8 min,并在生長素(IAA) 2.0 μM + 激動素(KT)5.0 μM + 水解酪蛋白 (CH) 100 mg·L-1的MS固體培養基上共培養2~3 d,然后轉入含 IAA 2.0 μM + KT 5.0 μM + CH 100 mg·L-1及60 mg·L-1Km 和1 000 mg·L-1頭孢霉素(cef)的MS 固體篩選培養基上培養,待不定芽長至1 cm時,轉移到含80 mg·L-1Km 和1 000 mg·L-1cef的MS + 2.0 mg·L-1IBA固體培養基上生根。
1.3 卡那霉素抗性植株的PCR分析和Southern雜交分析
對Km抗性植株總DNA進行PCR擴增,檢測目的基因OsVTE是否存在。PCR擴增時以未轉化普那菊苣植株為陰性對照,以質粒DNA為陽性對照。將擴增產物在1%的瓊脂糖凝膠中電泳,通過觀察特異性條帶的有無來初步確定目的基因是否已經轉入。PCR擴增的引物分別為:引物1:5’-CGCACAGCGGGTATCACTAT-3’;引物2:5’-AGCAAGCCAACCAGCAGTT-3’(由上海生工生物工程公司合成)。擴增片段大小為700 bp。PCR反應體系為20 μL。PCR擴增程序為:94℃預變性2 min;94℃變性30 s;53℃退火45 s;72℃延伸50 s;進行35次循環;然后72℃終延伸10 min。
為進一步證實PCR擴增結果的可靠性,提取未轉化植株和擴增結果為陽性的植株中隨機選取幾個擴增條帶亮度不同的陽性植株葉片總DNA進行Southern 雜交,以含有外源基因的質粒DNA為陽性對照。以限制性內切酶Hind Ⅲ 酶切20 μg 總DNA,37℃溫浴過夜。以地高辛(Digoxigenin (DIG) -dUTP)標記的OsVTE基因為探針,雜交溫度為68℃,過夜。以CSPD (4-methoxyspiro {1,2-dioxetane-3,2’- (5’- chloro) tricyclo [3.3.1.1] decan) -4-yl } phenyl phosphate (CSPD) 為發光底物。具體操作根據Southern 試劑盒(The Dig DNA Labeling and Detection試劑盒)操作手冊進行,CSPD 購于Boehringer Mannheim Co. Ltd, Germany。Southern 雜交中以地高辛標記的OsVTE擴增產物為探針。探針制備方法如下,
擴增體系:buffer: 2 μL; dNTP (地高辛標記):4 μL; p1:0.8 μL;p2: 0.8 μL; Taq (2 μ·μL-1):1 μL; ddH2O:10.4 μL;template(質粒DNA):1 μL 。
擴增程序:94℃ 5 min; 94℃ 30 s; 56℃ 50 s; 72℃ 50 s; 72℃ 10 min;循環35次。
1.4 普那菊苣干旱脅迫處理方法
選取未轉化植株和Southern雜交分析檢測含單拷貝外源基因轉基因植株,做以下處理:
在普那菊苣培養基中分別添加不同濃度的蔗糖、瓊脂、甘露醇,造成不同干旱脅迫的培養基上培養30 d,后在含2.0 mg·L-1IBA 的MS 固體生根培養基上恢復生長30 d (25 ℃,12 h 光照,12 h 黑暗)。蔗糖濃度設為:20(CK),30,40,50,60 g·L-1;瓊脂濃度設為:6(CK),8,10,12,14 g·L-1;甘露醇濃度設為:0(CK),10,20,30,40 g·L-1。每組處理3個,設3組重復,觀察植株的生長情況。
選取5個在試驗室抗性試驗中表現較好的T0代轉基因株系的植株幼苗擴繁,將在MS培養基中生長20 d的幼苗移栽在直徑10 cm、高10 cm 的塑料營養缽中,以蛭石作基質,培養30 d 左右,然后移栽至花盆中,14 d后進行旱脅迫處理。試驗設3個處理,甘露醇濃度為20,30,40 g·L-1。每組處理20 株,設3組重復,光照培養30 d,觀察植株的生長情況。
1.5 生理指標的測定
分別對蔗糖濃度為50 g·L-1、瓊脂濃度為10 g·L-1和甘露醇濃度為30 g·L-1脅迫條件件下生長30 d,以及在含2.0 mg·L-1IBA的MS固體生根培養基上恢復生長30 d后的轉基因植株的SOD活性、POD活性以及MDA含量進行測定[15]。
1.6 數據分析
試驗數據采用Excel 2003及用 DPS 7.05 軟件進行方差分析,顯著水平為0.05。
2 結果與分析
2.1 轉OsVTE 基因植株的獲得與鑒定
普那菊苣外植體在經過農桿菌感染轉化后,在篩選培養基上分化培養30 d左右,外植體分化出現明顯差異,經60 mg·L-1Km 篩選后,部分外植體無法形成再生芽,或產生的再生芽在培養中白化死亡。將篩選后長至0.5~1.0 cm的綠色抗性再生芽轉移到MS + 2.0 mg·L-1IBA+ 80 mg·L-1Km + 1000 mg·L-1cef 的固體生根培養基上繼續進行篩選培養,部分再生芽無法形成根狀物。將得到的轉化植株在 MS + 2.0 mg·L-1IBA固體生根培養基上繼續培養30 d后,進行PCR 檢測。普那菊苣共處理外植體640 塊,509 塊形成愈傷組織,得到109 株再生植株。

圖1 普那菊苣轉化植株再生Fig.1 Regeneration of transgenic plants of Puna Chicory
用目的基因OsVTE的特異引物對轉化處理后再生的普那菊苣進行PCR 檢測,從109 株轉化植株中,有35 株中擴增出預期大小(700 bp)的片段,而未轉化植株中沒有擴增出該片段,初步證明OsVTE基因已成功地轉入普那菊苣植株。
從PCR 檢測呈陽性的材料中隨機選取幾株擴增片段亮度不同的材料,提取DNA,以HindⅢ限制性內切酶消化后,以OsVTE基因為探針進行Southern blot 雜交分析,雜交結果如圖2所示。雜交分析結果表明有3株為單拷貝插入,其余2株為多拷貝插入,未轉化植株則無雜交信號。
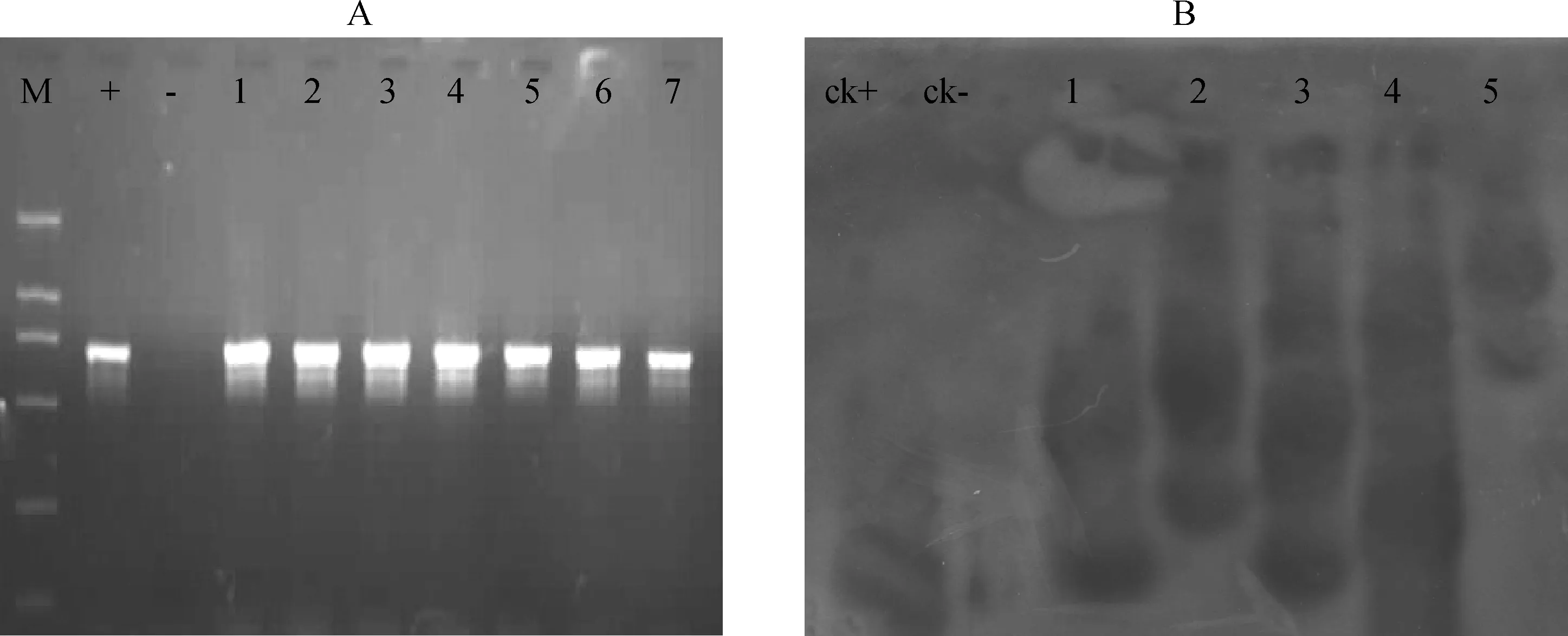
圖2 部分轉化植株檢測結果Fig.2 Tests for some transgenic plants
2.2 蔗糖濃度對普那菊苣組培苗生理指標的影響
對普那菊苣組培苗培養基添加蔗糖改變滲透壓脅迫培養30 d 后,再轉移到正常蔗糖濃度培養基上恢復生長30 d,從組培苗的株高變化上看,低濃度(CK)蔗糖對轉基因植株幼苗和未轉化植株幼苗的生長沒有顯著的抑制作用。隨著蔗糖濃度的升高,抑制作用逐漸加強。蔗糖濃度達到50 g·L-1時未轉化植株幼苗生長受到明顯抑制,株高較小,植株生長緩慢;轉基因植株幼苗仍然保持良好的生長態勢。50 g·L-1蔗糖是未轉化植株幼苗生長可以承受的最大濃度。在蔗糖濃度為50 g·L-1下(稱為延緩生長),對未轉化植株幼苗和轉基因植株幼苗的SOD,POD活性以及MDA含量進行測定。轉基因植株幼苗與未轉化植株幼苗相比SOD、POD活性增強、MDA含量下降。停止脅迫后組培苗恢復生長30 d后(稱為恢復生長)的SOD活性、POD活性以及MDA含量各處理間差異均不顯著。
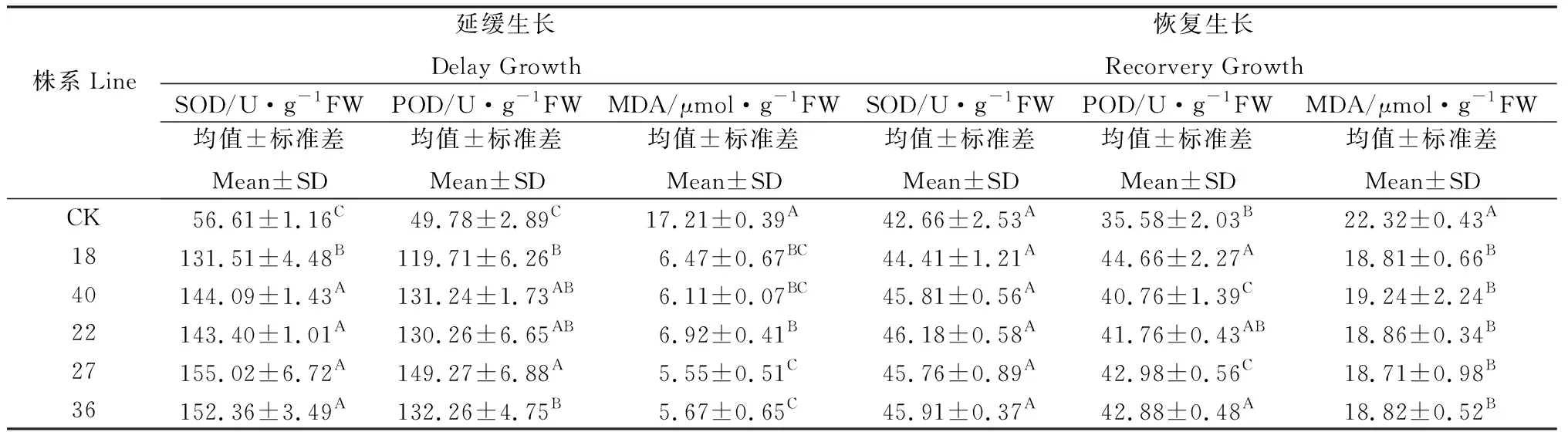
表1 蔗糖濃度對普那菊苣組培苗SOD、POD 活性和 MDA 含量的影響Table 1 Effects of sucrose content on the SOD,POD activities and MDA content in leaves of chicory plants
2.4 瓊脂濃度對普那菊苣組培苗生理指標的影響
對普那菊苣組培苗培養基添加瓊脂改變滲透壓脅迫培養30 d 后,再轉移到正常瓊脂濃度培養基上恢復生長30 d。瓊脂濃度低于10 g·L-1時,未轉化植株幼苗不如轉基因植株幼苗生長旺盛,但尚未明顯表現出受抑制;瓊脂濃度高于10 g·L-1時,未轉化植株幼苗的生長受到明顯得抑制,生長緩慢,植株矮小、根系生長弱。瓊脂濃度為10 g·L-1時(稱為延緩生長),對其生理指標進行測試,普那菊苣轉基因植株幼苗SOD 、POD 活性顯著增加,MDA 含量下降。停止脅迫后,組培苗恢復生長30 d后(稱為恢復生長)的SOD活性、POD活性以及MDA含量各處理間差異均不顯著(表2)。
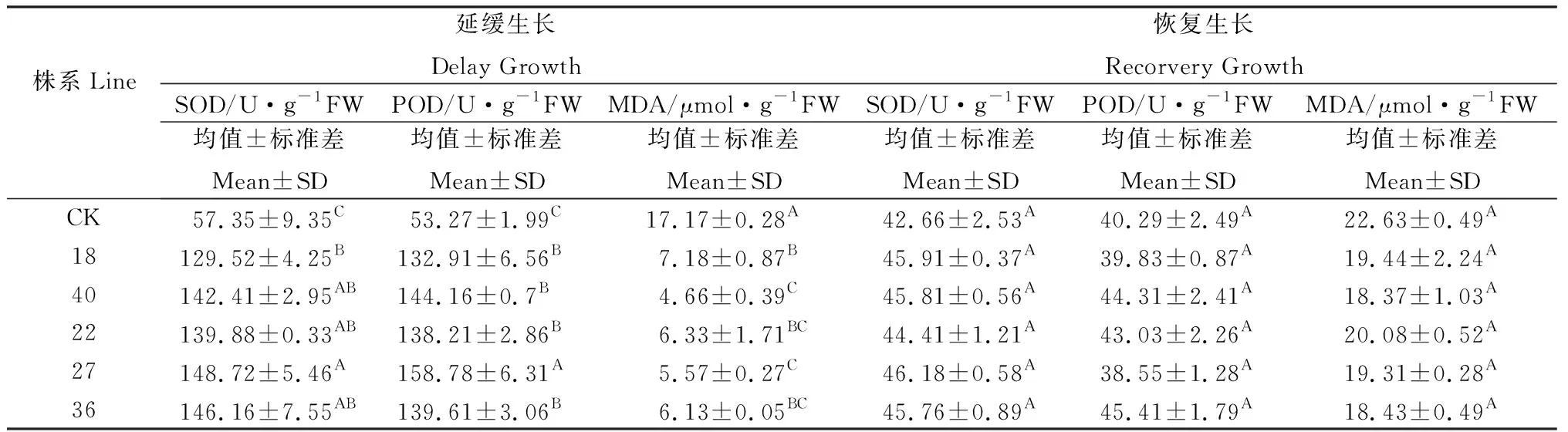
表2 瓊脂濃度對普那菊苣幼苗SOD、POD 活性和MDA 含量的影響Table 2 Effects of agarose concentrations on the SOD 、POD activities and MDA content in leaves of chicory plants
2.5 甘露醇濃度對普那菊苣苗生理指標的影響
對普那菊苣組培苗培養基添加甘露醇改變滲透壓脅迫培養30 d 后,再轉移到正常培養基上恢復生長30 d。當甘露醇濃度為20 g· L-1時,未轉化植株幼苗生長緩慢,出現玻璃化現象,葉片呈淡綠色、萎蔫狀,可以生根生長,但生根緩慢,并在基部開始形成少量的保護性的致密細胞團。甘露醇濃度為30 g·L-1時,未轉化植株幼苗葉片卷曲變形,嚴重失綠,幼葉完全不能夠正常伸展,根部基本褐化壞死;而轉基因植株幼苗在甘露醇的脅迫下葉片變得窄而細,但是仍然能夠正常生長,葉片也依然呈現綠色,能夠正常生根。甘露醇濃度高于30 g·L-1時,轉基因幼苗葉子卷曲、變形,葉片顏色逐漸變為黃綠色,根系不能正常生長。在對不同株系的轉基因普那菊苣株系幼苗的觀察中發現,不同株系之間對甘露醇的耐受力存在著一些微小的差異,可能與外源基因的拷貝數、插入點和表達量有一定的關系。綜上所述,30 g·L-1的甘露醇是未轉化植株幼苗可以承受的最大濃度,40 g·L-1的甘露醇是轉基因植株幼苗可以承受的最大濃度。甘露醇濃度為30 g·L-1時(稱為延緩生長),對其生理指標進行測試,普那菊苣轉基因植株幼苗SOD、POD活性顯著增加,MDA含量下降。停止脅迫后組培苗恢復生長30 d (稱為恢復生長)后的SOD 活性、POD 活性以及MDA 含量各處理間差異均不顯著(表3)。
將含有OsVTE基因的普那菊苣T0代轉基因株系進行盆栽,14 d后進行甘露醇脅迫處理,甘露醇濃度為20,30,40 g·L-1,每組處理20 株,3 組重復。脅迫30 d,甘露醇濃度為30 g·L-1時,未轉化植株幼苗葉片枯黃死亡,而轉基因植株幼苗葉片呈萎蔫狀、黃色,脅迫解除后可正常生長。
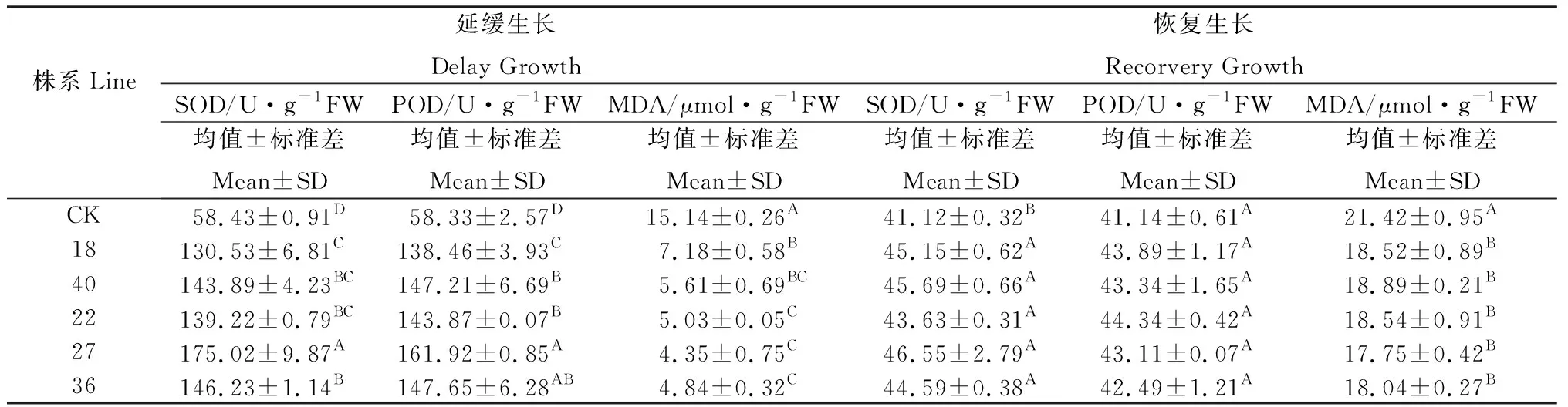
表3 甘露醇濃度對普那菊苣組培苗SOD、POD和MDA含量的影響Table 3 Effects of mannitol stress on the SOD, POD activities and MDA content in leaves of chicory plants

圖3 普那菊苣在甘露醇脅迫下的轉基因與非轉基因外植體的表現Fig.3 Performance of Puna chicory explants and seedlings under the mannitol stress: transgenic vs non-transgenic.
3 討論與結論
普那菊苣轉化試驗中共處理外植體640 塊,509 塊形成愈傷組織,得到109 株再生植株,其中35 株中擴增出預期大小(700 bp)的片段,初步證明OsVTE基因已成功地轉入普那菊苣植株。
在培養基中添加適當濃度的蔗糖能夠增強組培苗干物質含量,促進多糖物質的合成和貯藏,增強組培苗的抗逆能力,有利于組培苗的生長,但蔗糖濃度過低或過高則會增加培養基的滲透壓,使多糖物質的合成和貯藏受阻,影響營養物質的吸收及植株的正常生長[16]。在培養基中添加甘露醇降低了細胞膨壓和滲透勢,提高了培養基的滲透勢負值,造成水分逆境,使水分和養分吸收受阻,減少營養消耗。同時抑制細胞壁酶活性,減弱新陳代謝活動,延緩細胞生長[17]。對高濃度滲透壓處理下以及恢復生長后SOD、POD活性和MDA含量的變化研究表明,增加滲透壓處理后的組培苗SOD、POD活性較對照高,MDA含量較對照低。說明轉基因幼苗在增加滲透壓后發生生理代謝的協調變化以降低逆境對普那菊苣組培苗細胞膜系統的傷害,增強保護酶的活性,從而增強植株對環境的抗逆性,延緩衰老,達到提高逆境生存能力的目的。
猜你喜歡
中國民間療法(2021年5期)2021-06-09 09:21:26
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
食藥用菌(2016年6期)2016-03-01 03:24:27
護士進修雜志(2015年5期)2015-03-18 19:22:53
中國藥業(2014年24期)2014-05-26 09:00:09
