農桿菌介導小麥成熟胚遺傳轉化的幾個影響因素研究
2017-09-16 00:25:28李朝煒劉穎王丹娜陽秦婧魏景芳
江蘇農業科學 2017年12期
李朝煒+劉穎+王丹娜+陽秦婧+魏景芳


摘要:為研究農桿菌介導的小麥成熟胚轉化的影響因素,以2個小麥品種石4185、科麥一號的成熟胚為轉化受體材料,分別以GV3101、EHA105、LBA4404等3種農桿菌菌株(含有pCAMBIA1380雙元載體)為供體材料進行轉化,主要檢測成熟胚愈傷組織β-葡萄糖苷酸酶(GUS)報告基因的表達情況。結果表明,不同小麥品種對轉化體系的反應各不相同,菌株、受體的基因型以及外植體的繼代時間和生理狀態等均對轉化效率有很大的影響。GV3101的侵染轉化效果優于另外2個菌株,石4185成熟胚愈傷對農桿菌侵染的敏感性比科麥一號要高。繼代培養時間的延長會不同程度地影響小麥成熟胚愈傷的轉化效果。通過對轉化體系幾個影響因素的研究,有助于提高農桿菌介導成熟胚轉化體系的效率。
關鍵詞:小麥;成熟胚;農桿菌;GUS表達率
中圖分類號: S512.101文獻標志碼: A文章編號:1002-1302(2017)12-0045-03
lizhaowei79@163.com。小麥是世界三大糧食作物之一,種植較為廣泛,一直占據著世界糧食產量的重要地位。近年來,通過分子改良的方法對其進行研究以緩解目前糧食短缺問題已逐漸成為研究重點[1]。在針對小麥的轉基因研究中,基因槍法是應用最多的轉化方法,但同時也存在試驗成本高、轉化效率較低以及外源基因容易存在多個拷貝等缺點[2-3],相比較而言,利用農桿菌介導法進行小麥遺傳轉化則具有操作簡單、成本較低、所轉移基因拷貝數少、可轉移較大DNA片段等優點,近年來已成為研究者關注的熱點,成功獲得了眾多轉化植株[4-8]。
農桿菌介導法轉化小麥大多以幼胚作為外植體,可獲得相對較高的誘導率和再生率,但是取材會面臨諸多困難和限制;而以成熟胚作為外植體,具有取材便利、無氣候與季節性影響、可保證不同個體間生理狀況一致等優點,具有較高的研究價值[9-10]。小麥的基因組非常復雜,不同基因型之間遺傳轉化條件差異較大,轉化體系的有效性及轉化機制等諸多問題尚待解決。目前用農桿菌介導法進行小麥遺傳轉化的體系與玉米、水稻等單子葉植物相比仍然存在明顯差距,而這種普遍較低的轉化效率也制約了小麥基因工程改良的進一步發展[11-12]。本研究以小麥成熟胚為轉化受體,針對菌株、基因型、愈傷培養時間等幾個影響農桿菌轉化小麥的重要因素進行比較和優化,為進一步提高侵染效果、改善農桿菌介導的小麥遺傳轉化體系提供參考。
1材料與方法
1.1植物材料
供試小麥品種為普通冬小麥品種石4185和科麥一號,均由筆者所在實驗室保存。
1.2菌株及植物表達載體
農桿菌菌株為EHA105、GV3101和LBA4404;植物表達載體為pCAMBIA1380,該質粒T-DNA區帶有由CaMV35S啟動子驅動的β-葡萄糖苷酸酶(GUS)基因、潮霉素(HYG)基因。均由筆者所在實驗室保存。
1.3方法
1.3.1種子滅菌與愈傷組織誘導選取無霉變的飽滿種子,經75%乙醇消毒2 min,無菌水沖洗干凈,用0.1%氯化汞消毒8~10 min,再用無菌水沖洗3次,置于適量無菌蒸餾水中,25 ℃避光浸泡16~18 h。在超凈臺中取出種子,小心剝離出胚,盾片向上置于誘導培養基(MS+2.0 mg/L 2,4-D+30 g/L 蔗糖+8 g/L瓊脂,pH值5.8)上。置于培養箱中 25 ℃ 暗培養。愈傷誘導15 d后切去長出的芽,挑選長勢良好的愈傷組織,去掉胚后轉接入繼代培養基(MS+2.0 mg/L 2,4-D+0.4 g/L 2-(N-嗎啡啉)乙磺酸(MES)+150 μmol/L 乙酰丁香酮+40 g/L麥芽糖+8 g/L瓊脂,pH值5.8)。如有需要每20 d在相同培養基上繼代1次。
1.3.2小麥愈傷的農桿菌侵染及共培養侵染前2~3 d挑取含重組質粒的農桿菌單菌落,接種于5 mL含有50 mg/L利福平和50 mg/L卡那霉素的YEB液體培養基中,28 ℃、200 r/min 振蕩培養過夜。再按1 ∶50的比例轉接到含有相同抗生素的液體YEB培養基中,28 ℃、200 r/min振蕩培養至D600 nm為0.6~0.8。離心收集菌體,重懸于侵染培養基(1/10MS+2.0 mg/L 2,4-D+400 mg/L MES+200 μmol/L乙酰丁香酮+40 g/L麥芽糖,pH值5.8)中,調整D600 nm為0.6。在超凈臺中選取淡黃色致密的愈傷組織,轉入鋪有滅菌紗布的培養皿中,倒入含有菌體的侵染培養基,將愈傷組織完全浸泡,室溫侵染40 min。隨后用滅菌紗布將愈傷組織托起并瀝干液體,再將愈傷組織轉移到鋪有3層滅菌濾紙的培養皿中,置于24 ℃黑暗條件下共培養2~3 d。
1.3.3脫菌與抗性愈傷篩選將共培養后的愈傷組織移入滅菌培養瓶中,用脫菌液(MS+2.0 mg/L 2,4-D+40 g/L麥芽糖+500 mg/L特美汀)清洗3次,每次浸泡30 min,最后棄去脫菌液,將愈傷組織移至滅菌濾紙上,在超凈臺中干燥1 h。將愈傷組織轉移到含有300 mg/L特美汀的繼代培養基中,24 ℃ 黑暗恢復培養7 d。再將其轉入篩選培養基(2.0 mg/L 2,4-D+50 mg/L潮霉素+300 mg/L特美汀+40 g/L麥芽糖+8 g/L瓊脂)中,26 ℃黑暗條件下進行篩選培養。
1.3.4GUS染色GUS活性的組織化學染色檢測,按 Jefferson 的方法[13]進行。待經過農桿菌轉化的愈傷組織在含有潮霉素抗性的篩選培養基上篩選一段時間之后,每個小麥品種各挑取一部分抗性愈傷組織進行GUS表達檢測。
1.3.5試驗結果分析統計本研究中相應公式如下:
GUS表達率=(GUS表達受體組織塊數/感染總受體組織塊數)×100%;endprint
抗性愈傷率=(抗性愈傷數/供試愈傷數)×100%。
2結果與分析
2.1不同農桿菌菌株對小麥成熟胚愈傷轉化的影響
本研究分別采用攜帶表達載體的3種農桿菌菌株(EHA105、GV3101、LBA4404)對石4185和科麥一號的成熟胚愈傷組織進行侵染轉化,經恢復培養7 d后轉入含有潮霉素抗性的篩選培養基中,篩選一段時間之后,挑選抗性愈傷組織進行GUS基因檢測。結果顯示,3種農桿菌菌株對小麥成熟胚愈傷的侵染轉化效果不同,EHA105和GV3101對愈傷的侵染效果要明顯優于LBA4404(圖1)。經EHA105和GV3101侵染轉化的愈傷組織,在抗性篩選5 d之后,2個小麥品種的絕大部分愈傷組織呈現明顯的藍色。篩選20 d之后,愈傷組織GUS染色的面積明顯縮小、顏色變淡,并且僅有部分愈傷組織表面可見GUS基因表達。篩選5 d時,GUS基因仍屬于瞬時表達,此時呈現較深的藍色,說明以35S啟動的GUS基因在小麥愈傷組織細胞中得到了表達;篩選20 d時,不同處理中顯示藍色的部分均明顯減弱且面積變小,此時GUS基因已經基本整合到植物基因組中。經LBA4404侵染轉化的愈傷組織,抗性篩選5 d時GUS表達情況不強,整體染色較淺;而篩選至20 d時,大部分愈傷組織塊已無明顯GUS基因表達,說明此菌株對這2種小麥愈傷的侵染效果較差。
2.2不同基因型小麥成熟胚愈傷組織的轉化效率比較
石4185和科麥一號這2個小麥品種經過不同農桿菌轉化并經抗生素篩選20 d后,分別統計其愈傷組織的GUS表達率。圖2結果表明,GV3101菌株對石4185的轉化效果最好,GUS表達率達到64.3%;LBA4404菌株對這2種小麥成熟胚愈傷的侵染效果均較差,GUS表達率均不足30.0%,尤其是對科麥一號的轉化,GUS表達率僅為16.0%;EHA105菌株的轉化效果居中,并且對2種小麥的轉化效果差異不大。
從圖2還可以看出,石4185和科麥一號這2個小麥品種在相同農桿菌菌株及轉化條件處理時的轉化效率有所差異。石4185對3種農桿菌侵染的反應效果均不同,經GV3101菌株的侵染效果最佳,篩選20 d后的GUS表達率最高;經EHA105菌株侵染的效果居中,GUS表達率為42.8%;經LBA4404菌株侵染的效果最不敏感,GUS表達率為29.3%。3種農桿菌侵染石4185的表達率差異明顯。科麥一號經EHA105、GV3101菌株的侵染轉化效果差異不明顯,GUS表達率分別為32.7%、31.0%;經LBA4404菌株侵染的效果最差,GUS表達率僅有16.0%。
2.3不同繼代培養時間對小麥成熟胚愈傷轉化的影響
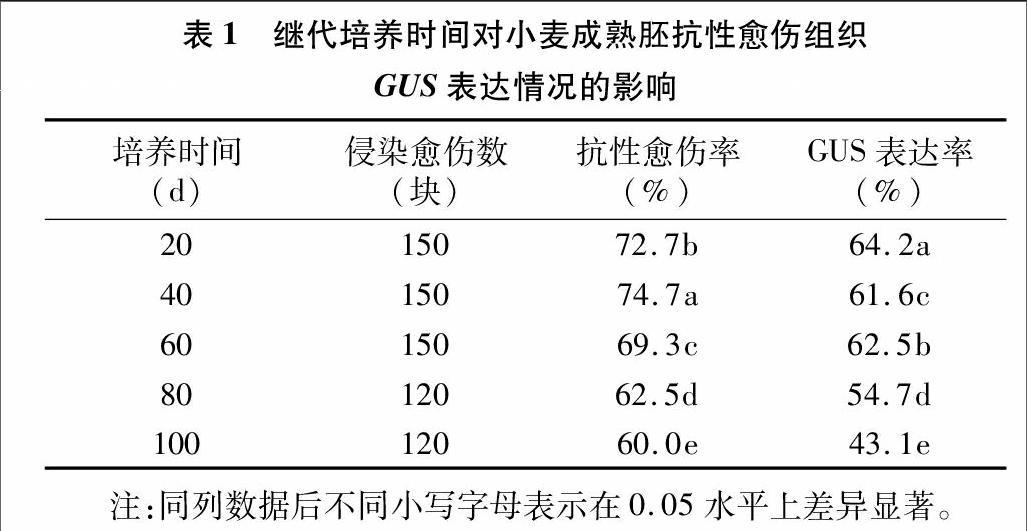
在探索不同農桿菌菌株及不同基因型小麥在轉化效率方面的差異之后,選取石4185作為受體材料,以GV3101為侵染菌株,進一步分析繼代培養時間對成熟胚愈傷轉化效率的影響。在試驗中分別選取誘導后繼代培養不同時間的愈傷組織,每個處理條件接種120~150個愈傷塊進行試驗,分別經農桿菌介導轉化及潮霉素抗性篩選20 d后計算抗性愈傷的獲得率,隨后將抗性愈傷進行GUS染色分析并統計表達率。
表1結果顯示,隨著繼代培養時間的延長,石4185愈傷的轉化效果會受到一定的影響,培養時間不同,石4185的抗性愈傷率和GUS表達率在0.05水平存在著顯著差異。在愈傷培養時間超過60 d后,抗性愈傷率和GUS表達率均隨培養時間的延長而下降。隨著培養時間的延長,GUS表達率比抗性愈傷率的下降程度更為明顯。因此,針對本試驗所采用的轉化體系,在進行成熟胚愈傷的農桿菌侵染轉化時,愈傷的繼代培養時間不可過長,控制在2個月之內為好。
3結論與討論
農桿菌介導法是目前植物上應用最為廣泛的基因轉化方法之一,自20世紀80年代出現以來,已經廣泛應用于煙草、棉花、馬鈴薯等多種雙子葉植物和玉米、水稻等一些單子葉植物中。自Cheng等首次利用農桿菌侵染獲得轉基因小麥以來[5],近年來對其轉化體系的研究已取得不少成果。為進一步提高轉化效率,研究者在添加酚類物質、調整菌體濃度和侵染時間、選用高滲處理或表面活性劑、采用不同共培養方式等方面做過很多有效的探索和嘗試。此外,菌株、受體基因型、外植體的類型、繼代時間和生理狀態等也被證實對轉化效率具有很大的影響[2-4,11-15]。
研究者普遍認為,不同的農桿菌菌株對小麥外植體的侵染能力存在差別,即使對同一基因型的小麥侵染能力也不同[2-3,6,11]。本研究中3種菌株GV3101、EHA105和LBA4404介導的對小麥成熟胚愈傷的轉化效果也體現出這一點。此外,從本研究可見,經EHA105侵染的愈傷在抗性篩選5 d時GUS染色的效果較GV3101侵染時要略深一些,但篩選20 d后卻比GV3101表現要差一些。其原因是在篩選5 d時,GUS基因尚屬瞬時表達,EHA105的瞬時轉化效果更好,很可能是由于該菌株侵染毒力強;而在篩選20 d時,絕大多數顯示GUS染色陽性的愈傷組織已經是穩定表達,而GV3101的穩定轉化效果要強于EHA105。同時,本研究發現,LBA4404的轉化效果不佳,可能是由于與EHA105、GV3101的侵染毒性相比,LBA4404更為溫和。但也有研究顯示,LBA4404溫和的侵染毒性更適于進行轉化[2]。而多數研究者的研究結果顯示,侵染毒性較強的農桿菌菌株對小麥的轉化效果較好。
在農桿菌介導小麥的遺傳轉化中,不同基因型小麥之間也存在對農桿菌敏感性的明顯差異。從本研究可見,利用3種菌株進行侵染時,石4185的敏感性均優于科麥一號,尤其對GV3101的敏感性最強,GUS表達率最高。因此,小麥基因型之間的差別,不僅是愈傷再生體系方面的主要影響因素,同時也對遺傳轉化效率具有重要影響,這也是小麥復雜性的又一體現[2-4,11,14]。本研究將不同小麥基因型與不同農桿菌菌株對轉化效率的影響結合起來進行對比研究,對于小麥轉基因育種水平的提高具有一定的現實意義。endprint
雖然GUS表達率并不能完全代表小麥農桿菌侵染的最終轉化效率,但可以作為一項重要的參考指標。以往研究顯示,能夠檢測出GUS基因表達的受體,分化再生之后對其進行PCR檢測,一般也能夠檢測出目的基因;而即使GUS基因無表達時,目的基因也并非一定沒有轉化成功,可能是GUS基因表達很弱,肉眼觀察不到所致[16]。因此在本研究中選用這樣一項簡單直觀的報告基因來進行轉化體系效率的分析,可以直觀地了解到轉化體系的效果。
綜上所述,不同的農桿菌菌株對小麥的侵染轉化效果不同;同一菌株對不同基因型小麥的侵染能力也不同。因此利用農桿菌介導法進行小麥轉基因研究時,應當針對具體所用的外植體狀態,采用適宜菌株,探索最佳轉化條件,才能獲得較高的轉化效率。
參考文獻:
[1]奚亞軍,路明. 小麥轉基因技術的研究現狀及在育種上的應用[J]. 中國農學通報,2002,18(3):55-57,108.
[2]王永勤,肖興國,張愛民. 農桿菌介導的小麥遺傳轉化幾個影響因素的研究[J]. 遺傳學報,2002,29(3):260-265.
[3]宋成麗,王翾,徐虹,等. 農桿菌介導的小麥成熟胚轉化的影響因素[J]. 麥類作物學報,2012,32(2):209-214.
[4]張月琴,陳耀鋒,王麗,等. 小麥成熟胚培養條件的優化及高效基因型篩選[J]. 干旱地區農業研究,2014,32(5):100-105.
[5]Cheng M,Fry J E,Pang S,et al. Genetic transformation of wheat mediated by Agrobacterium tumefaciens[J]. Plant Physio1,1997,115:971-980.
[6]Xia G M,Li Z Y,He C X,et al. Transgenic plant regeneration from wheat(Triticum aestivum L.) mediated by Agrobacterium tumefaciens[J]. Acta Phytophysiol Sin,1999,25(1):22-28.
[7]葉興國,Shirley S,徐慧君,等. 小麥農桿菌介導轉基因植株的穩定獲得和檢測[J]. 中國農業科學,2001,34(5):465-468.
[8]Wu H,Sparks C,Amoah B,et al. Factors influencing successful Agrobacterium-mediated genetic transformation of wheat[J]. Plant Cell Rep,2003,21(7):659-668.
[9]劉香利,劉縉,郭藹光,等. 小麥不同外植體離體培養與再生研究[J]. 麥類作物學報,2008,28(4):568-572.
[10]安海龍,衛志明,黃健秋. 小麥幼胚培養高效成株系統的建立[J]. 植物生理學報,2000,26(6):532-538.
[11]趙芳方,田保明,位芳. 基于農桿菌介導的小麥遺傳轉化體系的影響因素的分析[J]. 安徽農業科學,2013,41(19):8443-8445.
[12]李朝煒,姚彬,苗苗,等. 不同品種小麥成熟胚愈傷組織的誘導與分化[J]. 湖北農業科學,2015,54(13):3279-3282.
[13]Jefferson R A. Assaying chimeric genes in plants:the GUS gene fusion system[J]. Plant Molecular Biology Reporter,1987,5(4):387-405.
[14]王艷麗,葉興國,劉艷鵬,等. 農桿菌敏感小麥基因型的篩選研究[J]. 麥類作物學報,2005,25(6):6-10.
[15]武海娜,張永升,王小龍,等. 農桿菌介導的小麥幼胚遺傳轉化體系的優化[J]. 麥類作物學報,2012,32(3):421-426.
[16]郭志江,丁在松,王金明,等. 農桿菌介導遺傳轉化敏感基因型小麥的篩選鑒定[J]. 華北農學報,2008,23(4):81-84.胡云平,張靜,劉丹. 水肥耦合對春小麥葉片生態特性及產量的影響[J]. 江蘇農業科學,2017,45(12):48-52.endprint
