不同葉位和離體失水時間對烤煙葉片保水能力的影響
2017-09-27 02:10:50張小全許志文闞洪贏張林張鋆鋆郭傳濱楊立均夏宗良
中國煙草學報 2017年4期
關鍵詞:煙草
張小全,許志文,闞洪贏,張林,張鋆鋆,郭傳濱,楊立均,夏宗良
1 河南農業大學煙草學院 鄭州 450002;
2 河南省煙草公司駐馬店市公司 駐馬店 463000;
3 河南農業大學生命科學學院 鄭州 450002
農藝與調制
不同葉位和離體失水時間對烤煙葉片保水能力的影響
張小全1,許志文1,闞洪贏1,張林1,張鋆鋆1,郭傳濱2,楊立均2,夏宗良3
1 河南農業大學煙草學院 鄭州 450002;
2 河南省煙草公司駐馬店市公司 駐馬店 463000;
3 河南農業大學生命科學學院 鄭州 450002
為闡明葉位和離體時間對烤煙葉片保水能力的影響。以2個田間抗旱性不同的烤煙品種豫煙6號和農大202的離體葉片為材料,采用離體葉片自然失水法,研究了不同葉位和離體時間對葉片失水率、抗氧化酶活性、激素和抗逆脅迫響應基因表達的影響。結果表明:5個不同葉位的失水率為離體0~24 h較高,24 h后失水速率逐漸變緩,葉位間差異不顯著,農大202失水率顯著高于豫煙6號;離體葉片內抗氧化酶(SOD、POD、CAT)活性、可溶性蛋白含量、脫落酸(ABA)含量均在離體24 h時達到最高值,丙二醛(MDA)含量也在24 h后增速變緩;4個脅迫響應基因(AREB、CDPK2、ERD10C、LEA5)相對表達量也在24 h達到最高,說明烤煙葉片離體24 h生理代謝反應最強,2個品種在離體24 h各指標差異最大。因此,烤煙葉片保水力與取樣葉位關系不大,離體葉片失水24 h為測定煙草保水力的合適時間。
烤煙;葉片保水力;抗氧化酶活性;脅迫響應基因
煙草起源于降雨充沛的熱帶,整個生育期對水分的需求都很高。我國多數優質煙區位于丘陵山區,季節性干旱嚴重影響煙葉質量的提高和產量的穩定[1]。通過改善生產條件和應用栽培技術來緩解雨養煙區的水分脅迫問題是有限的,因此選育優質抗旱品種對穩定和發展旱地煙葉生產有著重要意義。鑒定抗旱相關性狀及不同品種的抗旱機制,成為煙草抗旱育種的重要前提。抗旱性鑒定指標多種多樣,有干旱條件下的生物量[2]、細胞膜透性[3]、氣孔阻力[4]、抗氧化酶活性[5]、葉片保水力[6]等。葉片保水力是葉片在離體條件下具有保持原有水分的能力,其保水力的強弱與植物遺傳性、細胞特性和原生質膠體性質有關,離體葉片的保水力可以反應植物原生質的耐脫水能力和葉片角質層的保水能力,在一定時間內含水量越高,表明葉片保水力越強,抗旱性也越強。Clarke等[7-10]最先研究了不同發育時期的葉片、不同離體時間的小麥葉片的保水力差異,并將葉片保水力作為小麥的抗旱性鑒定指標。葉片保水力具有測定方法簡單、時間短、工作量小等優點,目前在小麥[11]、玉米[12]、油菜[13]、煙草[14]、紅樹[15]等抗旱性鑒定當中有大量應用。煙草葉片較大、水分含量較高,國內外大量研究在測定葉片保水力時,選用的葉位、離體時間不同,沒有統一的標準[8,14]。本試驗擬對不同葉位和離體時間對烤煙葉片保水能力的影響進行研究,以期探明不同葉位和離體時間煙草葉片保水力的變化規律,為應用葉片保水力作為煙草抗旱性鑒定和抗旱育種選擇指標提供依據。
1 材料與方法
1.1 試驗材料與處理
選用抗旱性能不同的豫煙6號和農大202為試驗品種[6]。試驗于2016年在河南農業大學許昌校區進行,選取2個品種整齊一致的煙苗于4月30日移栽至大田,株距55 cm,行距120 cm。每個品種種植4行,每行100株。供試土壤為輕壤土,土壤中堿解氮含量為48.8 mg.kg-1,速效磷含量為51.7 mg.kg-1,速效鉀含量為103.9 mg.kg-1,pH為7.7。田間管理措施按當地優質煙葉栽培技術規程進行。
在移栽后60 d,旺長后期,現蕾前,選取2品種自下而上第1、4、7、10、13葉位的葉片各6片用于不同葉位對葉片保水力的影響研究;選取2個品種第10葉位的葉片各36片用于不同失水時間對葉片相關生理指標的影響研究。選取得各葉位的葉片均為正常生長,朝向一致的葉片。
1.2 測定指標與方法
1.2.1 葉片失水率和葉片保水力測定
采用離體葉片自然失水法測定,具體步驟如下:在早上8:00~9:00,在田間摘取葉片后,迅速裝入塑料封口袋,并密封袋口,置于冰盒內迅速帶回實驗室。實驗室保持恒溫、避風、無陽光直射,將葉樣從塑料袋內取出,迅速擦干葉片上沾有的水分,用電子天平稱取葉樣鮮重(FW0),而后懸掛于室溫25±1°C,相對濕度40%的室內,每隔12 h再次稱重,獲得每個樣品的FW12、FW24、FW36、FW48。24 h和48 h稱重后,每個處理選取3片葉105℃殺青30 min,95℃烘干至恒重,稱取干重獲得DW24和DW48。分別在0、12、24、36、48 h節點各選取6片葉,用于生理指標、激素和基因表達分析,每項指標3次生物學重復。葉片失水率和葉片保水力采用下面公式計算:

1.2.2 生理指標測定
抗氧化酶活性測定參考Li Q的方法[16],取0.5 g葉片于預冷的研缽中,加入1 mL磷酸緩沖液在冰浴下研磨成漿,加緩沖液使最終體積為5 mL。取2 mL于12000 r/min下離心10 min,上清液即為SOD、POD、CAT粗提液,比色并計算酶活。丙二醛(MDA)含量和相對電導率參照李合生等[17]方法測定。可溶性蛋白含量測定參照考馬斯亮藍(Bradford)法測定[18]。激素含量的測定采用酶聯免疫吸附分析法[19]。
1.2.3 脅迫響應基因的相對表達量分析
樣品總RNA的提取按照美國Invitrogen公司TRIZOL試劑說明書操作,反轉錄取2 μgRNA為模版,參照立陶宛Fermentas公司的RevertAidTM First Strand cDNA Synthesis Kit產品說明書進行反轉錄,合成第一鏈cDNA。PCR擴增采用BioRad iQ5實時定量PCR儀進行。反應結束后分析熒光值變化,每項指標3次生物學重復。目的基因的相對表達量在將CT值導入Excel2010后按照公式2-△△CT計算[20],并繪制基因表達差異圖。內參基因為煙草ACTIN2基因,表1為4個脅迫響應基因的PCR擴增引物信息。

表1 熒光定量PCR分析用的引物Tab.1 Primers used for quantitative real-time PCR
1.3 數據處理
試驗數據的統計分析采用DPS 7.05[21]以及Excel 2010進行。
2 結果與分析
2.1 不同葉位葉片含水率和失水率比較
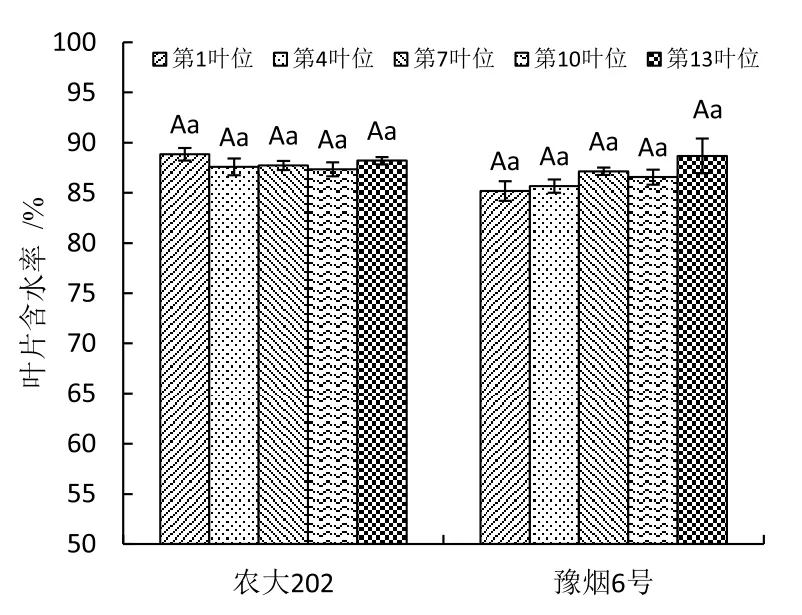
圖1 不同葉位葉片含水率比較Fig.1 Comparison of leaves water content in different leaf positions
在旺長后期對豫煙6號和農大202的第1、4、7、10和13葉位離體葉片取樣測定葉片含水率與葉片失水率。圖1顯示2個品種不同葉位葉片含水率在85%~90%之間,不同葉位之間無差異,品種間差異不顯著。葉片失水曲線如圖2所示,2個品種5個不同葉位的失水率為0~24 h較高,24 h后失水速率逐漸變緩,葉位間差異不明顯,農大202失水率顯著高于豫煙6號。
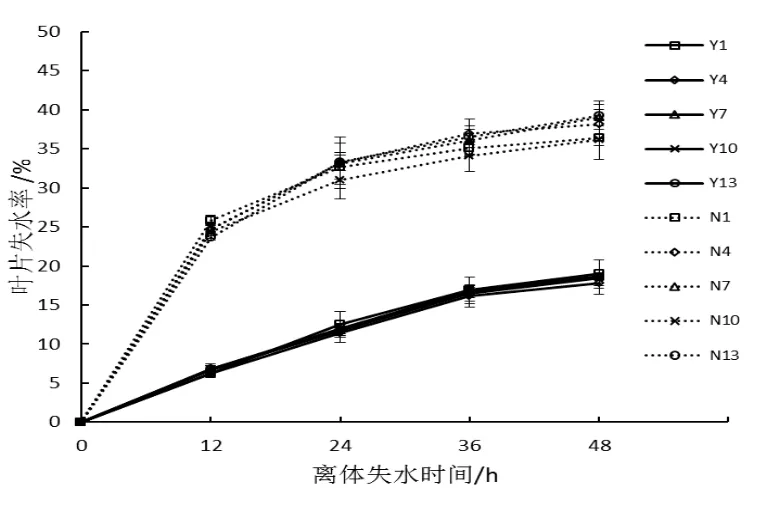
圖2 不同葉位葉片失水率比較Fig.2 Comparison of leaf water loss in different leaf positions
2.2 不同離體失水時間烤煙葉片生理變化
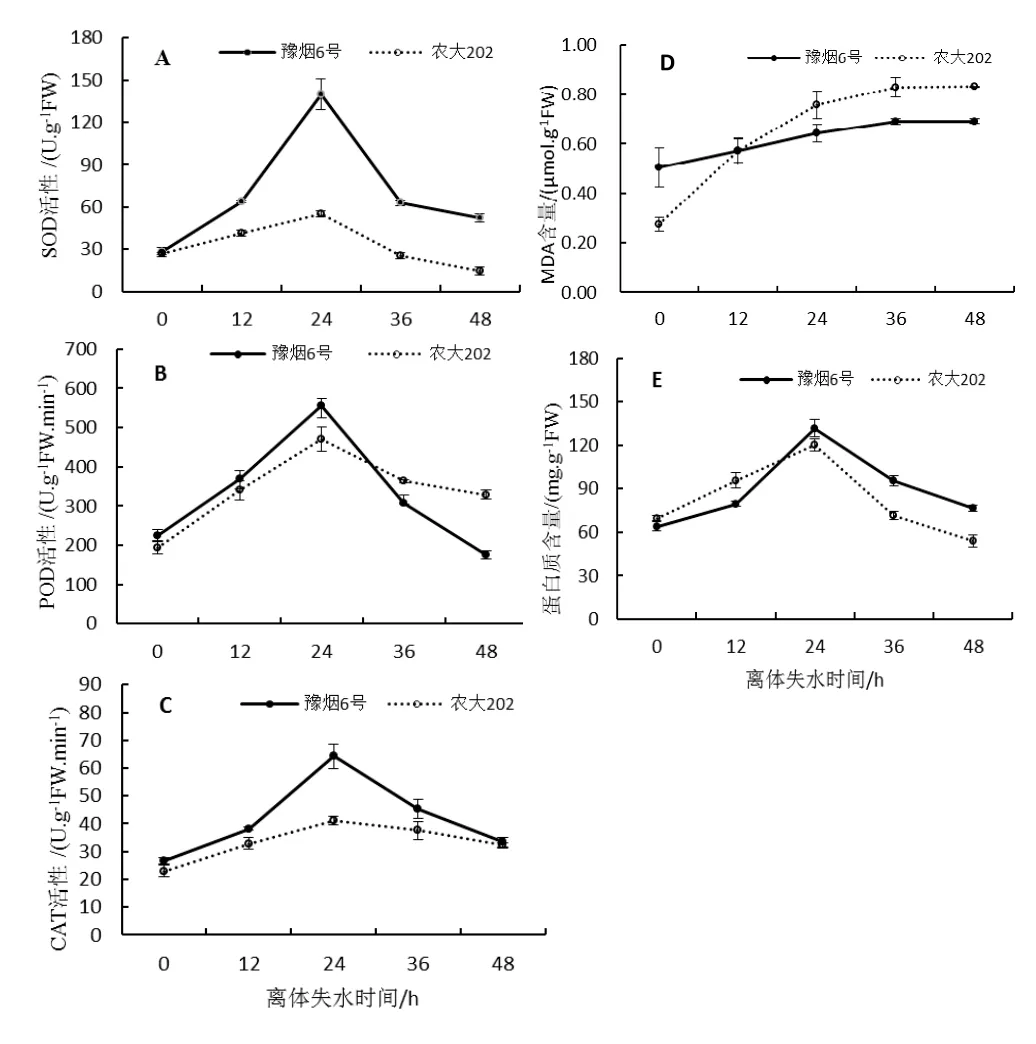
圖3 不同離體失水時間烤煙葉片生理變化Fig.3 Physiological changes of flue - cured tobacco leaves under different dehydration time
對豫煙6號和農大202不同離體失水時間的第10葉位葉片分別進行了SOD、POD、CAT活性,MDA含量以及可溶性蛋白含量測定。結果表明3種抗氧化酶活性均呈現出先升后降趨勢,且在24 h時達到最高值,整體上看豫煙6號抗氧化酶活性均高于農大202(圖3A、B、C),說明豫煙6號能對細胞膜起到更好的保護作用。MDA作為膜脂過氧化的產物,其含量呈不斷增加趨勢,但24 h后增速減緩,這說明膜脂過氧化在加劇但葉片失水24 h后過氧化程度在變緩,12 h后農大202的MDA含量超過了豫煙6號,說明隨失水時間農大202膜系統受傷害程度大于豫煙6號(圖3D)。可溶性蛋白含量也呈先升后降趨勢,失水24 h其含量達到最高值(圖3E)。這些表明烤煙葉片失水24 h后其生理活性減弱,葉片生命力開始降低。
2.3 不同離體失水時間烤煙葉片激素含量變化
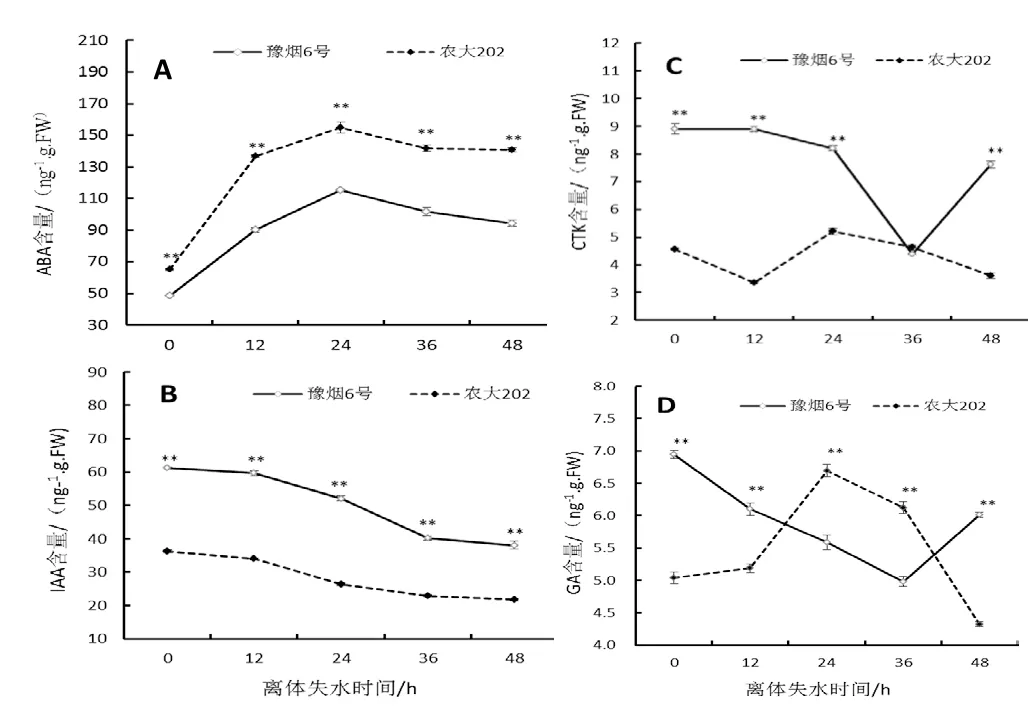
圖4 不同離體失水時間葉片激素含量變化Fig.4 Changes of hormone contents in tobacco leaves at different dehydration time
激素作為植物體內重要的微量調節和信號轉導物質,對植物抵御非生物脅迫具有重要作用。對2個品種不同離體失水時間的第10葉位葉片內的ABA、IAA、CTK和GA含量進行了測定,結果如圖4所示,ABA含量在葉片離體后迅速升高,在24 h達到最高值,而后開始下降(圖4A),并且農大202體內ABA含量顯著高于豫煙6號,這可能由于農大202遭受水分脅迫時其依賴ABA信號途徑的調控網絡反應較為敏感。IAA含量表現出逐漸降低趨勢,隨時間增加其降幅減緩,并且豫煙6號葉片內IAA含量高于農大202(圖4B)。CTK含量2個品種離體葉片均先下降后上升,但農大202在離體12 h前后下降后開始升高,而豫煙6號在36 h前后下降后開始升高(圖4C),CTK有延緩植物衰老的效應,2個品種離體葉片CTK變化的不同可能跟其葉片衰老進程不同有關。GA有提高生長素水平,抑制側芽休眠,衰老,加速細胞分裂,誘導α-淀粉酶的形成等作用,豫煙6號離體葉片GA從較高的水平先下降后上升,農大202離體葉片GA則從較低的含量水平先上升后下降,表現出不同的特征,表明2個品種離體葉片在失水衰老的過程中生理過程有著很大的不同。
2.4 不同離體失水時間烤煙葉片脅迫響應基因表達量變化
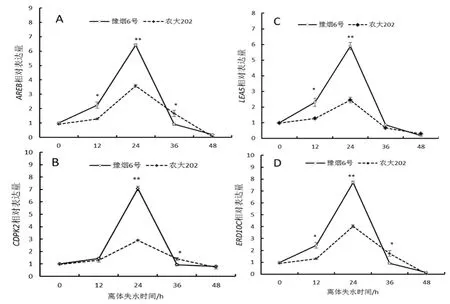
圖5 脅迫響應基因相對表達量Fig.5 Stress response gene relative expression
對4個脅迫響應基因(AREB、CDPK2、ERD10C、LEA5)的熒光定量PCR結果如圖5所示:隨葉片離體失水時間增加,4個基因的相對表達量均表現出先升后降的趨勢,并在24 h時達到最高值,表明葉片在24 h時應脅反應達到最強。在葉片離體失水12 h~24 h基因表達量迅速增加,說明葉片的應脅反應在此時間段較為劇烈,也反映了此時葉片已經受到了巨大的影響,而24 h~48 h表達量迅速下降說明了葉片因過度失水生命力開始漸失,生理活性減弱。整體上看豫煙6號脅迫響應基因的相對表達量顯著高于農大202,說明豫煙6號受到水分脅迫后應脅反應強于農大202,抗逆性相對較強。
2.5 不同葉位離體葉片保水力比較

圖6 不同葉位保水力比較Fig.6 The water retention capacity of excised leaves among different leaf positions
采用葉片離體失水24 h對農大202和豫煙6號進行了葉片保水力測定(圖6)。2個品種不同葉位間的離體葉片保水力均無顯著差異,說明離體葉片保水力與葉位無關。豫煙6號葉片保水力達到了85%以上,而農大202的葉片保水力在65%左右,差異達極顯著水平。表明豫煙6號離體葉片有相對較強的保水能力。同一品種不同葉位葉片具有不同的葉齡,其含水率和離體失水速率差異不大(圖1和圖2),葉片保水力沒有顯著性差異,可能跟這些葉片均為移栽后60天同時取樣,處在旺長后期,測定的第1、4、7、10、13葉位葉片均已定長,與葉片水分散失相關的角質層厚度、氣孔及細胞壁發育等已經基本定型,以及取樣前一段時間這些葉片的生長環境相同有關,而品種間差異顯著則可能主要跟品種本身的遺傳性、細胞原生質膠體性質等有關。
3 結論與討論
本研究表明同一品種烤煙的不同葉位葉片保水力及含水率無顯著差異,說明葉片保水力與葉位關系不大。在烤煙葉片失水24 h后,其抗氧化酶活性、可溶性蛋白含量、ABA含量以及脅迫響應基因相對表達量均達到最高值,而后開始下降。該結果說明離體葉片失水24 h后其生理活動由強變弱,葉片開始失去活力。這一現象可能跟植物生長發育的生物節律有關,離體條件下,烤煙葉片在自然環境中仍然要像正常生長條件下一樣努力維持晝夜生長節律。因此葉片失水24 h左右為煙草離體葉片保水力的合適研究時間。
植物通過保持吸水率與自身水分損失率間的平衡來防止其組織水分含量下降[22],當葉片離體以后吸水停止,只能通過保持水分來維持組織水分平衡。在此過程中細胞的代謝會因失水而改變,耐脫水的細胞必須做到能夠調節失水情況下的代謝和能修復失水代謝造成的損傷[22-25]。細胞開始脫水后,調節失水代謝的主要策略為關閉氣孔和調節滲透。因此水分脅迫后葉片內ABA和可溶性蛋白含量往往會大幅上升,ABA可以誘導氣孔關閉,并且是植物抵御脅迫的重要信號轉導物質。因此二者遇脅迫后含量上升幅度越大意味著葉片保水能力越強,抗逆能力越強。另外,葉片失水后IAA含量出現下降,推測是由于葉片為適應水分虧缺,抑制自身生長以減少水分需求,這也是細胞調節失水代謝的手段之一。細胞失水造成的損傷主要在細胞膜系統上,表現為膜脂過氧化和膜透性增加,故失水后葉片MDA含量會呈增加態勢,與此同時抗氧化酶活性會明顯升高以清除膜脂過氧化產生的大量活性氧,因此保水能力較強的葉片失水后MDA含量增幅較小,抗氧化酶活性較高。綜上可知,保水能力強的葉片失水后表現為可溶性蛋白含量及抗氧化酶活性相對較高,MDA含量增幅相對較低,這與本試驗結果相一致。
AREB轉錄因子基因是ABA信號途徑下游的關鍵調控因子,可啟動下游基因的表達,而AREB基因是植物應對滲透脅迫關鍵基因之一[26]。CDPKs即鈣離子依賴蛋白激酶基因,是植物細胞應對各種生物和非生物脅迫中承接Ca2+流變化的重要因素。研究表明CDPK2基因參與了植物對干旱的抵御反應[27]。LEA蛋白能在水分虧缺時束縛水分子并穩定蛋白質和細胞膜結構,其過表達可以提高轉基因煙草對滲透脅迫的抵抗力[28]。CBF類轉錄因子在植物應對低溫及水分脅迫中發揮著重要作用,ERD10C基因作為其下游基因,同樣對植物的抗逆反應起著關鍵作用。而Liu等的研究也證實了煙草在受到水分脅迫時ERD10C會表現出上調表達[29],本研究表明,失水后葉片的脅迫響應基因相對表達量顯著增加,且2個品種基因表達量高低與保水力趨勢相一致,說明煙草的葉片保水力以及各生理指標變化是受脅迫響應基因調控的。當細胞受到水分脅迫后首先會引發滲透勢的改變,繼而脅迫響應基因編碼大量蛋白以應對脅迫,主要包括起到抗性作用的蛋白(包括水通道蛋白、合成滲透保護物質的關鍵酶、起保護作用的蛋白酶以及解毒酶類)和對信號轉導起關鍵作用的蛋白因子(主要包括與第二信使生成有關酶類以及表達調控逆酶基因的有關轉錄因子)。通過這些脫水脅迫誘導基因產物實現了對失水情況下代謝的調節以及失水代謝造成損傷的修復。
植物的抗旱性通常是非常復雜的數量性狀,受多種因素的影響,選擇不同的指標研究得到的結論也不一樣。植物角質層是植物在4億5000萬年前向陸地過渡過程中進化的一項重要創新[30-31]。葉片是烤煙的主要蒸騰部位,水分經葉表面的蒸騰作用有兩條途徑:一是通過角質層蒸騰,另一種是通過氣孔蒸騰。烤煙幼葉因角質層發育不完全,角質層蒸騰幾乎占蒸騰量的40%~70%,而隨著葉片成年其角質層蒸騰減少,僅占總蒸騰量的5%~10%[32]。趙陽等研究表明水分脅迫情況下抗旱能力強的農大202的一級側根數目和二級側根密度顯著增加,說明農大202的抗旱機制可能主要在根系而不在葉片,因此其保水力相對豫煙6號較弱[33]。這也從側面反映了植物對于耐旱有著不同的機制,即根系發達具有較強的吸水能力或者葉片細胞組織彈性較高,細胞壁以及角質層較厚,具有較強的抗脫水能力。因此可對不同抗旱烤煙品種角質層及根系再做進一步深入研究。
[1]云南省煙草科學研究所.云南煙草栽培學[M].北京:科學出版社,2005: 15-16.Institute of tobacco science of Yunnan.Yunnan tobacco cultivation[M].Beijing: Science Press, 2005: 15-16.
[2]梁太波, 王建偉, 尹啟生,等.水分脅迫對烤煙氨同化關鍵酶活性和生物量的影響[J].中國煙草學報, 2012, 18(3): 50-54.LIANG Taibo, WANG Jianwei, YIN Qisheng, et al.Effects of water stress on ammonium assimilation and biomass of fl ue-cured tobacco[J].Acta Tabacaria Sinica, 2012, 18(3): 50-54.
[3]馬新蕾, 房燕, 王玉軍,等.十個烤煙品種的抗旱性鑒定[J].中國煙草學報, 2005, 11(5): 26-30.MA Xinlei, FANG Yan, WANG Yujun, et al.The evaluation of drought resistance of ten tobacco cultivars.Acta Tabacaria Sinica,2005, 11(5): 26-30.
[4]劉貞琦, 伍賢進, 劉振業.土壤水分對煙草光合生理特性影響的研究[J].中國煙草學報, 1995, 2(1): 44-49.LIU Zhenqi, WU Xianjin, LIU Zhenye.The effects of soil water content on photosynthetic characteristics of tobacco[J].Acta Tabacaria Sinica, 1995, 2(1): 44-49.
[5]裴斌,張光燦,張淑勇,等.土壤干旱脅迫對沙棘葉片光合作用和抗氧化酶活性的影響[J].生態學報,2013,(05):1386-1396.PEI Bin, ZHANG Guangcan, ZHANG Shuyong, et al.Effects of soil drought stress on photosynthetic characteristics and antioxidant enzyme activities inHippophae Rhamnoides Linn.Seeding[J].Acta Ecologica Sinica,2013,33(5):1386-1396.
[6]任慶成,陳秀華,張生杰,等.不同烤煙品種抗旱生理特征比較研究[J].西北植物學報,2009,29(10):2019-2025.REN Qingcheng, CHEN Xiuhua, ZHANG Shengjie, et al.Comparison of drought resistance characteristics of different fl uecured tobacoo varieties[J].Acta Botanica Boreali Occidentalia Sinica, 2009, 29(10): 2019-2025.
[7]Clarke J M, Mccaig T N.Excised-leaf water retention capability as an indicator of drought resistance ofTriticumgenotypes.[J].Canadian Journal of Plant Science, 1982, 62(3): 571-578.
[8]Clarke J M, Mccaig T N.Evaluation of techniques for screening for drought resistance in wheat.[J].Crop Science, 1980, 22(3): 503-506.
[9]Clarke J M, Romagosa I, Jana S, et al.Relationship of excised-leaf water loss rate and yield of durum wheat in diverse environments.[J].Canadian Journal of Plant Science, 1989, 69(4): 1075-1081.
[10]Clarke J M, Townley smith T F.Heritability and relationship to yield of excised-leaf water retention in durum wheat[J].Crop Science, 1986, 26(2): 289-292.
[11]Winter S R, Musik K B Porter.Evaluation of screening technique for breeding drought-resistant winter wheat[J].Crop Science, 1988,28: 512–516.
[12]武斌,李新海,肖木輯,等.53份玉米自交系的苗期耐旱性分析[J].中國農業科學, 2007, 40(4): 665-676.WU Bin, LI Xinhai, XIAO Muji, et al.Genetic variation in fi ftythree maize inbred lines in relation to drought tolerance at seeding stage[J].Scientia Agricultura Sinica, 2007, 40(4): 665-676.
[13]王丹丹,唐章林,荊蓉蓉,等.甘藍型油菜遺傳圖譜構建及苗期耐旱相關性狀的 QTL 定位[J].西南大學學報(自然科學版),2014, (7): 8-16.WANG Dandan, TANG Zhanglin, JING Rongrong.et al.Construction of a molecular genetic map of rapeseed (Brassica napus L.)and QTL mapping of its drought tolerance-related traits[J].Journal of Southwest University(Natural Science Edition),2014, (7): 8-16.
[14]Han Y, Wang W, Sun J, et al.Populus euphratica XTH overexpression enhances salinity tolerance by the development of leaf succulence in transgenic tobacco plants[J].Journal of Experimental Botany, 2013, 64(14): 4225.
[15]Lechthaler S, Robert E M R, Tonné N, et al.Rhizophoraceae mangrove saplings use hypocotyl and leaf water storage capacity to cope with soil water salinity changes[J].Frontiers in Plant Science,2016, 7: 1-13.
[16]Li Q, Yu B, Gao Y, et al.Cinnamic acid pretreatment mitigates chilling stress of cucumber leaves through altering antioxidant enzyme activity.[J].Journal of Plant Physiology, 2011, 168(9): 927-934.
[17]李合生.植物生理生化實驗原理和技術[M].北京: 高等教育出版社, 2000: 280-283.LI Hesheng.Principles and techniques of plant physiological biochemical experiment[M].Beijing: Higher education press, 2000:280-283.
[18]Bradford M M.A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of proteindye binding[J].Analytical Biochemistry, 1976, 72: 248–254.
[19]Zhao J, Li G, Yi G X, et al.Comparison between conventional indirect competitive enzyme-linked immunosorbent assay(icELISA) and simplified icELISA for small molecules[J].Analytica Chimica Acta, 2006, 571(1): 79-85.
[20]Livak K J, Schmittgen T D.Analysis of relative gene expression data using real-time quantitative PCR and the 2-△△CTmethod[J].Methods, 2001, 25: 402-408.
[21]Tang Q Y, Zhang C X.Data Processing System (DPS) software with experimental design, statistical analysis and data mining developed for use in entomological research.Insect Science.2013,20(2) : 254-260.
[22]Verslues P E, Agarwal M, Katiyaragarwal S, et al.Methods and concepts in quantifying resistance to drought, salt and freezing,abiotic stresses that affect plant water status.[J].Plant Journal,2006, 45(4): 523-539.
[23]Leprince O, Deltour R, Thorpe P C, et al.The role of free radicals and radical processing systems in loss of desiccation tolerance in germinating maize (Zea mays L.)[J].New Phytologist, 2010,116(4): 573-580.
[24]Leprince O, Harren F J M, Buitink J, et al.Metabolic dysfunction and unabated respiration precede the loss of membrane integrity during dehydration of germinating radicles[J].Plant Physiology,2000, 122(2): 597-608.
[25]Walters C, Pammenter N W, Berjak P, et al.Desiccation damage,accelerated ageing and respiration in desiccation tolerant and sensitive seeds[J].Seed Science Research, 2001, 11(2):135-148.
[26]Fujita Y, Yoshida T, Yamaguchishinozaki K.Pivotal role of theAREB/ABF-SnRK2pathway inABRE-mediated transcription in response to osmotic stress in plants[J].Physiologia Plantarum,2013, 147(1): 15.
[27]Urao T, Katagiri T, Mizoguchi T, et al.Two genes that encodeCa-dependent protein kinases are induced by drought and highsalt stresses inArabidopsis thaliana[J].Molecular Genetics and Genomics, 1994, 244(4): 331-340.
[28]Yang L, Li W, Jiang S, et al.Group 5 LEA protein,ZmLEA5C,enhance tolerance to osmotic and low temperature stresses in transgenic tobacco and yeast[J].Plant Physiology & Biochemistry,2014, 84:22.
[29]Liu X, Liu S, Wu J, et al.Overexpression of Arachis hypogaeaNAC3in tobacco enhances dehydration and drought tolerance by increasing superoxide scavenging[J].Plant Physiology &Biochemistry, 2013, 70(1):354-359.
[30]Chen Guoxiong, Takao Komatsud, Ma Jianfeng et al.An ATP-binding cassette subfamily G full transporter is essential for the retention of leaf water in both wild barley and rice[J].Proceedings of the National Academy of Sciences, 2011, 108(30): 12354–12359.
[31]Karolina M.Jarzyniak, Micha Jasin ski.Membrane transporters and drought resistance – a complex issue[J].Frontiers in Plant Science, 2014, 5: e687.
[32]劉國順.煙草栽培學[M].北京:中國農業出版社, 2003: 221-222.LIU Guoshun.Tobacco Cultivation[M].Beijing: China Agriculture Press, 2003: 221-222.
[33]趙陽,王樹聲,張亞麗,等.增加煙草一級和二級側根是抵御干旱的生理機制[J].植物營養與肥料學報,2017,23(2): 548-555.ZHAO Yang, WANG Shusheng, ZHANG Yali, et al.Role of increasing first-and second-order lateral roots of tobacco for drought tolerance[J].Journal of Plant Nutrition and Fertilizer, 2017,32(2): 548-555.
Effects of stalk positions and natural dehydration after priming on water retention capacity of fl ue-cured tobacco leaves
ZHANG Xiaoquan1, XU Zhiwen1, KAN Hongying1, ZHANG Lin1, ZHANG Yunyun1, GUO Chuanbin2, YANG Lijun2, XIA Zongliang3*
1 College of Tobacco Sciences, Henan Agricultural University, Zhengzhou 450002, China;
2 Henan Zhumadian Municipal Tobacco Company, Zhumadian 463000, Henan, China;
3 College of Life Sciences, Henan Agricultural University, Zhengzhou 450002, China
In order to elucidate the effect of stalk positions and natural dehydration after priming on water retention capacity of fl ue-cured tobacco leaves, evaluation index of drought resistance of tobacco leaves was investigated.Effects of different stalk positions and natural dehydration after priming on leaf water loss rate, antioxidant enzyme activity, hormone and stress resistance gene expression were studied through natural dehydration method with two fl ue-cured tobacco varieties, Yuyan 6 and Nongda 202 of different drought resistance.Results showed that the water loss rate of leaves of 5 different stalk positions was higher during 0-24h, and gradually slowed down after 24h, and there was no signi fi cant difference among different stalk positions, but water loss rate of Nongda 202 was signi fi cantly higher than that of Yuyan 6.Antioxidant enzyme (SOD, POD, CAT) activity, soluble protein content, abscisic acid (ABA) content in excised leaves reached the highest value 24 hours in vitro, and Malondialdehyde (MDA) content also slowed down after 24 hours.The relative expression of four stress responsive genes (AREB, CDPK2, ERD10C, LEA5) reached the highest level after 24 hours.Physiological metabolism of excised fl ue-cured tobacco leaves was the strongest 24 hours in vitro, and the difference between the indexes of the two varieties was the greatest 24 hours in vitro.There was little relationship between water retention capacity and leaf position in excised fl ue-cured tobacco leaves, and leaf water loss 24 hours in vitro was the appropriate time to determine water retention capacity of excised tobacco leaves.
fl ue-cured tobacco;water retention capacity of leaves; antioxidant enzyme activity; stress responsive genes
張小全,許志文,闞洪贏,等.不同葉位和離體失水時間對烤煙葉片保水能力的影響[J].中國煙草學報,2017, 23(4)
河南省煙草公司駐馬店市公司項目和河南省煙草公司重點項目(HYKJ201401)
張小全(1980—),博士,副教授,主要從事煙草品種改良和推廣研究,Email: zxq013415@163.com
夏宗良(1973—),博士,教授,Email:xiazl08@163.com
2017-04-06;< class="emphasis_bold">網絡出版日期:
日期:2017-07-18
:ZHANG Xiaoquan, XU Zhiwen, KAN Hongying, et al.Effects of stalk positions and natural dehydration after priming on water retention capacity of fl ue-cured tobacco leaves [J].Acta Tabacaria Sinica, 2017, 23(4)
*Corresponding author.Email:xiazl08@163.com
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
中國煙草學報(2019年5期)2019-11-14 07:54:12
首都公共衛生(2019年5期)2019-05-21 01:08:34
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
新聞傳播(2016年3期)2016-07-12 12:55:34
當代化工研究(2016年9期)2016-03-20 16:22:08
自動化博覽(2014年6期)2014-02-28 22:32:15
聲屏世界(2014年6期)2014-02-28 15:18:09
西南學林(2013年2期)2013-11-12 12:58:54
中國煙草學報(2012年5期)2012-04-12 06:21:18
