iNDF對亞熱帶反芻動物采食量的調節作用及對iNDF評估的重要性
2017-10-13 09:21:41李蕾蕾
中國飼料 2017年18期
關鍵詞:模型
李蕾蕾 譯
(全國畜牧總站,北京朝陽 100125)
iNDF對亞熱帶反芻動物采食量的調節作用及對iNDF評估的重要性
李蕾蕾 譯
(全國畜牧總站,北京朝陽 100125)
纖維素特別是中性洗滌纖維(NDF)含量對飼料的消化吸收有很大的影響。目前,為提高反芻動物的生產性能和瘤胃健康及其對飼料的攝入量,商業飼料配方和動物模型生產性能方面的研究都在關注NDF。假設認為NDF的可消化性可在較小范圍內發揮作用,其飼料配方中通常配有特定水平的NDF含量。飼草的NDF,尤其是對于熱帶飼草(C4),在瘤胃中的潛在消化率存在極大的差異。潛在可消化性被定義為在長時間發酵后NDF中消失的的那一部分,其余的難消化成分(iNDF)不能被微生物利用。據推測,日糧中的iNDF能促進瘤胃食糜填充和日糧攝入。不通過參考iNDF直接制定日糧配方,使NDF達到特定水平會顯著影響飼料的消化、吸收和能量代謝。結論:營養模型需要修改,直接引入iNDF這一指標。
iNDF;NDF;C4 牧草;攝入
1 前言
飼草的可消化性和反芻動物對飼草的消化能力在很大程度上與中性洗滌纖維(NDF)的含量有關。目前,對商業飼料成分和動物模型生產性能的研究,都是在不過渡犧牲瘤胃健康的情況下,研究總NDF量如何改善飼料攝入以及提高動物生產性能。反芻動物飼料配方中NDF有個特定水平,基本假設認為NDF的可消化性范圍很窄。當飼喂溫帶飼草或者玉米青貯時,這個假設成立,但是當對熱帶飼草(C4)進行大范圍選擇時,NDF在反芻動物的可消化性上表現出差異。這種纖維的可消化性直接與NDF的潛在可消化部分有關 (pdNDF)(Poppi等 ,2000;Allen 等 ,1988)。潛在可消化性被定義為在長時間發酵后NDF中消失的那一部分,留下的是NDF的難消化成分 (iNDF),這部分難以被微生物利用(Wilkins,1972;Waldo 等,1972)。熱帶飼草也許含有相同的NDF量,但是在iNDF上差異顯著。Ellis等(1999)從營養模型預測,iNDF可以有效使瘤胃承載消化物,進而促進食物攝入。Lippke(1986)研究發現,在機體內,當iNDF在飼料干物質中的含量超過15%時,iNDF和采食量之間表現出很強的負相關。體內數據表明,iNDF能作為預測以飼草為基礎的飼料中有機物(OM)可消化性的指標 (Nousiainen等,2003)。如果不參考iNDF直接設計飼料配方,NDF將達不到一個特定的水平,這個配方會因為飼料攝入減少,飼料中可消化性及代謝能(ME)含量的減少而顯著影響能量供應。在飼草基本飼養系統中,所有的供能營養物質,NDF中可消化的比例對反芻動物的能量供應影響最大。在澳大利亞只有極少數營養學家把iNDF考慮進配方中。而且從飼料分析數據庫里并不容易獲取日糧中通用的飼草iNDF值。了解熱帶牧草品種通用的iNDF范圍的重要性,并對其進行評估能幫助營養學家預測NDF的可消化性,從而有效平衡飲食。
2 NDF的定義
NDF是代謝能(ME)的重要來源,可以監控瘤胃周轉率,幫助產生唾液,唾液可緩沖瘤胃內環境從而促進瘤胃的健康。NDF構成飼草的細胞壁,其成分是木質素復合基質,少量的蛋白質,各種多糖,特別是纖維素、半纖維素和果膠 (Moore和Hatfield,1994)。物種間、植物組織間及成熟植株機體內的細胞壁結構組成差異非常大。植物細胞壁的多糖能被瘤胃細菌、原生生物和真菌消化。Wilson(1995、1991、1990)的研究說明,植物根葉對瘤胃微生物的消化能力十分重要,木質素對這種消化能力有損害。一株植物在發育起始,細胞壁由一層初生壁構成,主要成分是纖維素。Weimer(1996)提出,當細胞完成生長和延伸,細胞壁會發生二次增厚,同時伴隨著大量的纖維素和半纖維素沉積。初生壁被木質化后,在細胞內腔緊接著進行次生壁的物質沉淀。Terashima等(1993)發現,初生壁的木質素濃度是最高的,由于次生壁的體積質量更大,所以次生壁所含的木質素更多。在成熟階段,根莖與葉子相比含有更多的次生壁組織(Wilson,1996),纖維素、木聚糖和木質素的濃度也更高(Jung,2012)。在瘤胃內,木質素是限制細胞壁消化的主要因子,由于木質素與多糖交聯,所以其能進一步限制多糖的消化(Jung,2012)。
瘤胃細菌進入非木質化的細胞壁有三種途徑,一種是通過細胞壁暴露的外表面,另一種是通過細胞腔進入細胞內部,還有一種是通過相鄰細胞的細胞壁進入(Engels等,2005;Jung,2001)。 初生壁中高濃度的木質素會形成阻止微生物的屏障,從而防止植物細胞被消化(Wilson,1996;Engels,1989)。瘤胃細菌從內部消化細胞,先攻擊次生壁,然后是初生壁。因為次生壁表面難以被瘤胃微生物進入,所以咀嚼和反芻過程對微生物的作用十分重要,此過程會對木質化的植物細胞進行物理破壞(Wilson,1996),同時增加微生物可遷移的表面積(Jung,2012)。Wilson(1995)推測,反芻動物在攝入草料后,約有1/3的細胞由于沒有暴露于瘤胃微生物環境,從而未被消化就離開瘤胃。木質素是一種非碳水化合類聚合物,由酚類單元組成,其可進一步延伸交聯形成木質素-碳水化合物復合體。Jung(1989)報道,飼草木質素是鄰甲氧苯基和紫丁香基單體的組合,后者隨著機體的成熟在組成中的比例會增加。木質素-碳水化合物復合體的性質隨著細胞壁的類型和植物的品種而變化(Chesson,1988)。
Grabber(2005)和 Casler等(1999)研究發現,在牧草中,木質素通過阿魏酸交聯與半纖維素緊密聯系,不依賴木質素的濃度就能降低微生物對細胞壁的消化能力。Jung(2010)試驗發現,多種玉米突變體,如棕色葉子突變體中阿魏酸脂儲量減少會導致細胞壁的可消化性增加。目前并不清楚C4飼草是否由大量的阿魏酸脂交聯,是否有研究的價值。推測在豆科植物中木質素與細胞壁多糖交聯,但是還沒發現其間的化學聯系(Jung,2004)。為更準確地假設木質素與日糧能量供應間的關系,需要更多的了解飼草組織中木質素的結構和化學特性。同樣作為一個預測指標,對iNDF的直接測定可以避免使用木質素,這個指標的選擇更實際些。
3 iNDF對反芻動物自由采食的重要性
Mertens(1994)和 Minson(1990)發現,在某些情況下,NDF可作為自由采食干物質(DM)攝入量的可靠預測指標,但是NDF含量和采食量間的關系很復雜,并不是簡單的線性關系。日糧中的NDF數量和質量能夠促進或限制采食量。NDF濃度較低(7.5%~35.5%)時,隨著日糧中NDF濃度的增加,干物質攝入量也逐漸增加,但是在高產動物中,當NDF濃度為22.2%增加到45.8%時,干物質采食量則逐漸減少(Arelovich等,2008)。在高濃度NDF的情況下,低干物質采食量與瘤胃的填充限制有關(Poppi等,2000)。干物質采食量與NDF間的關系不只依賴日糧中NDF的含量,更依賴于 NDF的潛在消化性 (pdNDF)(Poppi等,2000)。pdNDF被認為是NDF和iNDF間的差異。在NDF高水平的飼草中,iNDF是其消化速度的限速因素。
Huhtanen等(2006)發現,在反芻動物中,即使纖維在消化道中的停留時間足夠長,通過微生物消化仍然無法獲取iNDF。飼草中iNDF難以消化是因為細胞壁木質素和半纖維素的交聯(Van Soest,1994)。iNDF的過多攝入會限制反芻動物攝取足量飼草,從而影響營養補給。Poppi(2011)試驗證明,反芻動物對飼草基礎日糧的攝入由瘤胃填充物和填充物消失的速率所控制。填充物消失的速率主要受消化的遺傳率和通過率影響,不能消化的部分只能通過流道離開瘤胃,相比于潛在可消化部分,其會在瘤胃積聚,因此停留時間更長,不可消化部分在瘤胃長時間停留會導致采食量減少(Poppi,1981)。 Ellis (1999)建議在基礎飼料分析中應該把iNDF考慮進去,因為iNDF完全不被消化,是一個理想的用于估計pdNDF的部分。因此有人認為,應該明確日糧中iNDF所占比例。Lippke(1986)建議,每天iNDF最大的消耗約為每千克代謝體重 20 g (20 g/kg BW0.75·day),還需要更多的研究來確定這個值對不同生產系統和飼草是否相關。
4 NDF在日糧組成中的使用
目前對日糧組成和高性能反芻動物模型的商業關注聚焦在總NDF上,以便提高含高能日糧的攝入。國家研究委員會(NRC,2001)建議奶牛的日糧干物質中應該含有30%~35%的NDF。Arelovich等(2008)也證明,這個范圍可以達到提高干物質自由采食量和牛奶生產間的平衡,同時不會使牛奶中的脂肪減產。在亞熱帶/熱帶地區要達到這么低水平的NDF十分困難,因為這些地區以高水平NDF飼草為主,在亞熱帶放牧體系中40%的水平才是實際可行的。特別是在生產牛肉的飼育場,日糧中由于淀粉含量高,對NDF含量的水平要求更低(16%)。日糧中低水平的NDF能夠造成瘤胃的功能紊亂,因此為了動物的健康及符合市場規范,NDF需要達到最低水平。
以NDF含量為基礎,評估高產的反芻動物日糧仍然存在著問題,原因是飼草中NDF的變化因素太多。通過對200份夏日奶牛草料(溫帶和亞熱帶)進行NDF的分析發現,其含量為20%~80%。這與Jung和Allen (1995)的研究結果 (30%~80%)相似。隨著植物的生長,飼草中的NDF含量會增加(Jung,2012),這也與其生長環境,包括溫度、光照密度、水資源可利用度和地理緯度有關。即使把飼料中NDF含量的高變異性考慮上,NDF水平和動物性能之間的關系仍然不能得以確定。即使日糧中NDF的含量相似,飼草構成中NDF的消化程度仍然差異較大。這種多變性在亞熱帶地區尤為明顯。對200份夏日奶牛的飼草分析發現,NDF中的pdNDF含量為26%~90%。pdNDF的顯著差異對干物質采食量影響較大。當給反芻動物飼喂飼草料時,纖維可消化性的提高通常會改善動物生產性能。對pdNDF的精確估計是對改善日糧組成、飼草評估和生產反應的先決條件。
Waldo等 (1972)對潛在可消化性的概念建立的模型進行了優化,而這個模型對日糧攝入的調節和消化的研究仍然有參考作用。Poppi(1996)指出,在多種模型中不被消化的NDF是決定飼草利用的主要因素。營養模型如康奈爾凈碳水化合物和蛋白體系 (CNCPS) (Tylutki等,2008;Fox等,2004)以及北歐奶牛的新陳代謝模型“Karoline”(Danfaer等,2006)是通過間接計算iNDF得出OM和NDF的可消化性。瘤胃的NDF池與微生物N流最終會影響能量和微生物蛋白質的供應。
iNDF含量對采食量的影響在亞熱帶和熱帶地區的反芻動物放牧體系中特別明顯。不需減少日糧中NDF含量而又確保日糧中iNDF含量處于低水平,生產就能得到提高。為達到這個目的,除了通過降低牧草和青貯飼料的iNDF含量改善飼草質量外,還需要對iNDF含量的評估進行優化,而不僅僅依賴于NDF含量。
盡管iNDF在反芻動物生產體系中非常重要,但在澳大利亞的飼料分析數據庫中并不容易獲得,特別是對亞熱帶和熱帶地區生產體系的iNDF數據。在數據庫如Feedipedia或與流行的營養模型有關的飼料表中并不存在實際指標。信息的缺漏可能是因為評估iNDF具有難度。
5 iNDF的評估
對iNDF的測定方法有原位和體外技術。原位技術是通過在瘺管動物瘤胃放置的尼龍袋中培養樣本使飼草降解,袋子在第10~12天取出。Damiran 等(2002)和 Nocek(1988)都證明這種方法能使飼料在實際瘤胃環境中發酵,所以能進行有效的預測。但是,因為沒有一個標準的程序來執行,所以這個技術的可重復性很差 (Lund等,2004;Vanzant,1998)。
體外技術更適合于對大型飼養的評估。商業性飼料檢驗實驗室能夠為NDF的消化性提供體外測定。但是并沒有一個標準的分析程序,因此實驗室之間的分析結果差異較大。另外這些方法普遍是短時間發酵(24 h或48 h),因此不能測定出實際iNDF的成分,因為要完整測定出iNDF成分需要10 d的發酵時間。
體外方法是Goering和Van Soest (1970)體外程序的改進,草料在瘤胃的液體、緩沖液和礦物質中培養,接著進行中性洗滌劑的沖洗,然后干燥和稱重。體外技術是Tilley和 Terry(1963)體外DM兩步消化技術的改進,這種方法是飼料在反芻液和緩沖液中進行培養,固體殘留物在胃蛋白酶中繼續培養,以模仿皺胃的消化,把最終的殘留物進行干燥和稱重。瘤胃液為分析體系提供大量的酶類,可以有效地消化飼料樣品,但是這會給試驗間帶來巨大的誤差,因為瘤胃液中的種菌活性不同,這與供體動物及攝入的日糧質量有關。為使瘤胃液的有效性達到最大化,應該給供體動物飼喂相似的日糧(Vanzant等,1998;Nocek,1988),瘤胃的氨水平必須在 50 mg/L以上 (Satter等,1974)。 Rymer等(2005)發現,在一個固定時間收集瘤胃液,并且避免氧氣和唾液的污染,收集好的瘤胃液需要在39°C保存,并快速轉移到體外分析系統。ANKOMDaisy系統的發展是基于對Goering和Van Soest(1970)技術的改進形成,這個技術使用F57 Ankom過濾袋進行流線型NDF分析,這個過程不需要過濾消化樣品。經過長時間發酵,5 d后補充培養胃液。
假如對瘤胃的降解能進行有效的評估,那么試驗間在消化率和iNDF測定方面的差異就會縮小。由于瘤胃液的不同,一些差異的來源是無法避免的,但是應用統一的方法可以降低差異性,例如對飼草的磨削長度、袋子的孔隙率、開放表面區和樣本重與袋子大小的比值等進行標準化(Vanzant等,1998;Nocek,1988)。
Traxler等(1998)、Weiss等(1992)和 Chandler等(1980)試驗均發現,為測定iNDF而進行的長時間發酵過程比較復雜,且費用較高,因此許多營養模型嘗試從酸性洗滌劑木質素(ADL)的濃度預測出iNDF。根據Chandler (1980)結果推測,iNDF為測量的ADL含量的2.4倍,兩者間的關系廣泛用于多個實驗室和營養模型中。Van Soest(2005)等通過幾種飼草品種(玉米、苜蓿、草和小麥秸稈)也已經證實了這個結果,而且觀測和預期iNDF間的回歸(R2=0.94)相似。
但是,Harper等(2015)發現,這種簡單的數學關系對C4飼草并不適用,因為這種飼草的iNDF/ADL間關聯變化很大。本試驗使用長期發酵的方法對各種飼草的iNDF和ADL進行分析。iNDF/ADL比率為1.5~6.2,平均為3.82,這明顯高于CNCPS模型預測牧草iNDF時所適用的2.4。本研究與 Huhtanen 等(2006)、Nousiainen(2004)結果相似,對來自ADL含量的飼草iNDF進行預測,還需要進一步研究。
Raffrenato 等(2010)和 Vanzant(1998)研究發現,基于形態的相似性,針對牧草品種的特定方程式能提高ADL含量預測iNDF的準確性。Harper(2014)研究指出,在亞熱帶大多數飼草中,iNDF/ADL的比率差異很大。Kikuyu草場的iNDF/ADL的比率為 1.4~ 4.8。 Huhtanen(2006)發現,同種飼草品種內存在差異,個體品種的日糧為2.8~5.5。因此,使用同種飼草簡單的ADL模型對iNDF評估仍然不精確。在大部分亞熱帶和熱帶地區,iNDF的計算和模型中與ADL的關聯仍是個待解決問題。使用對C4飼草的方程式來預測飼草NDF的消化性并不精準,對預測ME含量也會造成誤差。
Van Soest(1994)發現,木質素并非細胞壁中均勻的物質,ADL與iNDF之間關系的多變可能是因為不同C4飼草品種成熟前后,木質素和半纖維素間交聯的差異造成。另外,ADL容易受環境條件的影響,例如溫度和光照密度等(Huhtanen等,2006)。比較偏冷環境中生長的溫帶飼草和惡劣環境中生長的C4飼草的ADL/iNDF存在差異。
6 對生產預測的影響
CNCPS營養模型中的iNDF是對動物生產性能預測的重要參數。飼喂C4牧草時,通過iNDF(ADL×2.4)預測并不可靠。因為在C4飼草方面,iNDF和ADL間的關系并不密切。總體來說,低估iNDF會造成對大多C4飼草的部分消化率(Kd)評估過高。Harper等(2014)研究結果如表1所示,說明真實的iNDF值能改變泌乳奶牛的產奶量和ME供給的預測。
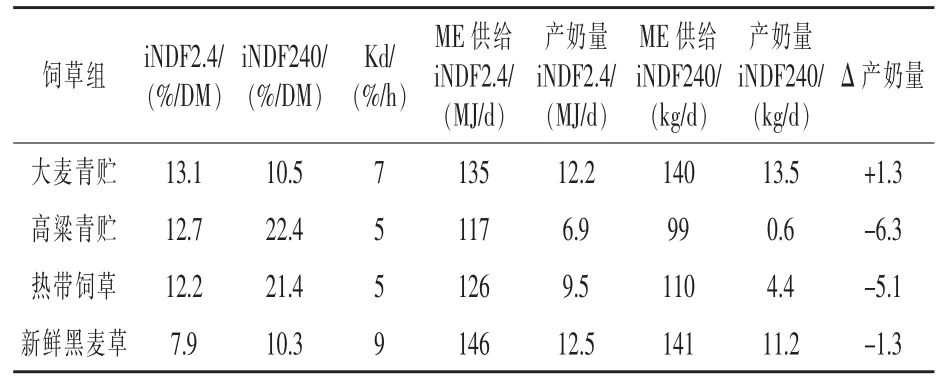
表1 不同飼草組預測的iNDF 2.4或體外iNDF 240
在大部分C4飼草組中,營養模型常會低估iNDF含量,因此會高估潛在的產奶量。假設直接測量iNDF能提高模型的精確度,這個設想需要利用C3和C4飼草進行全面的測試。
從ADL中計算iNDF,潛在ME是從NDF部分進行估計的,導致在CNCPS模型中對ME供給和產奶的預估過高,這與實際的iNDF估計相反。潛在消化的NDF和可利用的ME,是從NDF和iNDF含量間的差異進行估計的。反過來,iNDF含量是通過ADL含量乘以2.4估算出來(Traxler等,1998)。基于ADL×2.4模型估算iNDF在其他換算也有作用,在模型中這些換算可用來評估重要的代謝活動,包括對可利用的NDF部分(CB3),瘤胃降解的 CB3(RDCB3),瘤胃逸出的CB3 (RECB3)以及瘤胃逸出的 iNDF(RECC)進行預測,這些都受 iNDF 的影響(Tylutki等,2008)。這些部分由可利用碳水化合物組成,而碳水化合物可以在公式中計算ME供給。在CNCPS模型中,低估飼草的iNDF含量,將會導致高估奶牛的產奶量。
7 iNDF的替代性評估
Huhtanen等(2006)研究表明,對預測OM可消化性來說,難消化NDF參數在評估模型中使用可以提高精確度。但是,難消化NDF的使用因為需要長期發酵而受限制,而且其從ADL含量中預測并不準確。因此有一個短期的程序來預測iNDF是十分重要的。Deboever等(1995)發現,近紅外反射光譜(NIRS)是一個快速、有效且精準的飼草分析技術。
Kramer等(2015)對青貯牧草 NIRS和 iNDF間的緊密關系進行了報道。飼草樣本的NIR頻譜數據與OM成分有很強的關聯性,例如NDF、ADF、ADL 和粗蛋白質(Norris等,1976),這些物質直接與有機碳鍵關聯。這種關系可以在大樣本(>100)的校準公式中使用,預測多種OM成分。Mentink等(2006)通過試驗證明,預測的精準度依賴于頻譜數據和濕化學的顯著性和強相關性(R2> 0.85) 。
Harper等(2015)發現,使用大量C4飼草預測iNDF使NIRS校準更加全面。FT-NIR的使用可提供快捷和可重復的試驗結果。對飼草大樣本實際評估中,這些校準預測的iNDF結果比較準確。與目前使用的ADL估測iNDF值相比,由NIRS校準改進的iNDF值能提高飼喂和評估模型的精確度和實用性。
8 建議模型
目前營養模型建立的失敗歸結于這些模型不允許直接輸入iNDF值。以CNCPS為例,這個模型只允許輸入ADL值,然后根據iNDF=ADL×2.4公式計算iNDF。在這類模型中,只能通過調整ADL值來輸入飼草的實際iNDF值。例如,假如預期的iNDF值是12,那么輸入的ADL值就是12/2.4=5。模型需要進行調整以使能直接輸入iNDF值,跟ADL值的估計一樣簡單。
9 結論
iNDF含量在日糧可消化性和反芻動物飼料攝入的調節中起著重要的作用。這在C3與C4飼草的對比中尤為明顯,C4飼草有定向木質化的特性,可以使植物適應惡劣的環境。這些形態和解剖特性,可以為C4植物提供支持,但同時對微生物降解產生抗性,這影響了消化時其中能量的利用。C4飼草的NDF含量,依賴于植物的成熟階段和生長的環境條件,總NDF中擁有含量較高但多變的iNDF。相比于NDF,在模型中對iNDF的評估,能夠準確估計反芻動物對C4飼草的總能量攝入。飼草的長期發酵可以對iNDF含量精確估計,也可以使用NIRS進行精確和快速的iNDF評估。飼草樣本的iNDF可以應用到日糧配方的模型中,特別是C4牧草日糧。營養模型需要調整來接受直接測量的輸入,而不是通過數學衍化的iNDF值。這可提高對NDF可消化性的預測能力,從而可以更有效地平衡飲食,特別是對亞熱帶和熱帶的反芻動物生產體系中尤為適用。
[1]Allen M S,Mertens D R.Valuating constraints on fiber digestion by rumen microbes[J].J.Nutr,1988,118:261 ~ 270.
[2]Arelovich H M,Abney C S,Vizcarra J A,et al.Effects of dietary neutral detergent fiber on intakes of dry matter and net energy by dairy and beef cattle:Analysis of published data[J].Prof.Anim.Sci,2008,24:375 ~ 383.
[3]Chesson A.Lignin polysaccharide complexes of the plant-cell wall and their effect on microbial-degradation in the rumen[J].Anim.Feed Sci.Technol,1988,21:219 ~ 228.
[4]Danfaer A,Huhtanen P,Uden P,et al.The Nordic Dairy Cow Model,Karoline-Description [M].CABI Publishing:Wallingford,UK,2006.383 ~406.
[5]Damiran D,Del Curto T,Bohnert D W,et al.Comparison of techniques and grinding size to estimate digestibility of forage base ruminant diets[J].Proc.West.Sect.Am.Soc.Anim.Sci,2002,53:341 ~ 344.
[6]Ellis W C,Poppi D P,Matis J H,et al.Dietary-Digestive-Metabolic Interactions Determining the Nutritive Potential of Ruminant Diets[A].American Society of Animal Science[C].Champaign.IL,USA,1999;pp.423 ~ 481.
[7]Engels F M,Jung H J G.Alfalfa stem tissues:Impact of lignification and cell length on ruminal degradation of large particles[J].Anim.Feed Sci.Technol.2005,120:309 ~ 321.
[8]Fox D G,Tedeschi L O,Tylutki T P,et al.The Cornell Net Carbohydrate and Protein System model for evaluating herd nutrition and nutrient excretion[J].Anim.Feed Sci.Technol,2004,112:29 ~ 78.
[9]Grabber J H.How do lignin composition,structure,and cross-linking affect degradability[J].Crop Sci,2005,45:820 ~ 831.
[10]Huhtanen P,Nousiainen J,Rinne M.Recent developments in forage evaluation with special reference to practical applications[J].Agr.Food Sci,2006,15:293 ~ 323.
[11]Harper K J,Mc Neill D M.Direct in vitro measurement of i NDF to improve the prediction of milk production from subtropical forages[A].Mc Neill,D.M.,Bell,A.Proceedings of the 8th International Workshop on Modelling Nutrient Digestion and Utilization in Farm Animals[C].Cairns,Australia.CSIRO Publishing.2014.21.
[12]Jung H G,Engels F M.Alfalfa stem tissues:Rate and extent of cell-wall thinning during ruminal degradation[J].Neth.J.Agric.Sci,2001,49:3 ~ 13.
[13]Jung H G.Forage lignins and their effects on fiber digestibility[J].Agron.J,1989,81:33 ~ 38.
[14]Jung H G,Phillips R L.Putative seedling ferulate ester (sfe) maize mutant:morphology,biomass yield,and stover cell wall composition and rumen degradability[J].Crop Sci,2010,50:403 ~ 418.
[15]Jung H G,Allen M S.Characteristics of plant-cell walls affecting intake and digestibility of forages by ruminants[J].Anim.Sci,1995,73:2774 ~ 2790.
[16]Kramer M,Weisbjerg M R,Lund P.Estimation of indigestible NDF in feedstuffs for ruminants.Available online:http://forskningsbasen.deff.dk/Share.external?sp=Sce3d98ad-7aaa-444b-b101-2c4e7b1490ab&sp=Sau(accessed on 18 April 2015).
[17]Lippke H.Regulation of voluntary intake of ryegrass and sorghum forages in cattle by indigestible neutral detergent fiber[J].J.Anim.Sci,1986,63:1459 ~1468.
[18]Lund P,Weisbjerg M R,Ahvenjarvi S,et al.Nordic ringtest on INDF content and NDF degradation characteristics in three feeds[J].J.Anim.Feed Sci,2004,13:139 ~ 142.
[19]Moore K J,Hatfield R D.Carbohydrates and Forage Quality[A].Alliance ofcrop,soul,and environmentalscience society[C].Madison,WI,USA.1994.229~280.
[20]Mertens D R.Regulation of forage intake[A].Forage Quality,Evaluation,and Utilization[C].American Society of Agronomy,WI,USA.1994.450 ~493.
[21]Mentink R L,Hoffman P C,Bauman L M.Utility of near-infrared reflectance spectroscopy to predict nutrient composition and in vitro digestibility of total mixed rations[J].Dairy Sci,2006,89:2320 ~ 2326.
[22]Nousiainen J,Rinne M,Hellamaki M,et al.Prediction of the digestibility of the primary growth of grass silages harvested at different stages of maturity from chemical composition and pepsin-cellulase solubility[J].Anim.Feed Sci.Technol,2003,103:97 ~ 111.
[23]NRC.Nutrient Requirements of dairy Cattle,7th ed[M].Washington,DC,USA:National Academy Press,2001.
[24]Nousiainen J,Ahvenjarvi S,Rinne M,et al.Prediction of indigestible cell wall fraction of grass silage by near infrared reflectance spectroscopy[J].Anim.Feed Sci.Technol,2004,115:295 ~ 311.
[25]Poppi D P.Nutritional constraints for grazing animals and the importance of selective grazing behaviour[A].Grassland Productivity and Ecosystem Services[C].CABI:Wallingford,UK.2011.19 ~ 26.
[26]Poppi D P,Minson D J,Ternouth J H.Studies of cattle and sheep eating leaf and stem fractions of grasses.2:Factors controlling the retention of feed in the reticulo-rumen[J].Aust.J.Agric.Res,1981,32:109 ~ 121.
[27]Poppi D P,Minson D J,Ternouth J H.Studies of cattle and sheep eating leaf and stem fractions of grasses.1.The voluntary intake,digestibility and retention time in the reticulo-rumen[J].Aust.J.Agric.Res.1981,32,99 ~ 108.
[28]Poppi D P,France J,Mc Lennan,S R.Intake,passage and digestibility[A].Feeding Systems and Feed Evaluation Models[C].Theodorou M K,France J,.CAB International:Oxfordshire,UK,2000.35 ~ 52.
[29]Rymer C,Huntington J A,Williams B A,et al.In vitro cumulative gas production techniques:History,methodological considerations and challenges[J].Anim.Feed Sci.Technol,2005,123:9 ~ 30.
[30]Terashima N,Fukushima K,He L F,et al.Comprehensive model of the lignified plant cell wall[A].Forage Cell Wall Structure and Digestibility[C].ASA-CSSA-SSSA:Madison,WI,USA.1993.247 ~ 270.
[31]Tylutki T P,Fox D G,Durbal V M,et al.Cornell Net Carbohydrate and Protein System.A model for precision feeding of dairy cattle[J].Anim.Feed Sci.Technol,2008,143:174 ~ 202.
[32]Vanzant E S,Cochran R C,Titgemeyer E C.Standardization of in situ techniques for ruminant feedstuff evaluation[J].J.Anim.Sci,1998,76:2717 ~2729.
[33]Waldo D R,Smith L W,Cox E L.Model of cellulose disappearance from the rumen[J].J Dairy Sci,1972,55:125 ~ 129.
[34]Wilkins R J.Potential digestibility of cellulose in grasses and its relationship with chemical and anatomical parameters[J].J.Agric Sci,1972,78:457 ~464.
[35]Wilson J R.Influence of plant anatomy on digestion and fibre breakdown[A].Microbial and Plant Opportunities to Improve the Utilization of Lignocellulose by Ruminants[C].Elsevier Science Publishing Company:New York,NY,USA.1990.99~ 117.
[36]Wilson J R.Plant structures:Their digestive and physical breakdown[A].Recent Advances on the Nutrition of Herbivores[C].Malaysian Society of Animal Production:Kuala Lumpur,Malaysia,1991.207 ~ 216.
[37]Wilson J R.Organization of forage plant tissues[A].Forage Cell Wall Structure and Digestibility[C].Jung H G,Buxton D R,Hatfield R,et al.ASACSSA-SSSA:Madison,WI,USA,1993.1 ~ 32.
[38]Wilson J R,Mertens D R.Cell-wallaccessibility and cellstructure limitations to microbial digestion of forage[J].Crop Sci,1995,35:251 ~ 259.
[39]Weimer P J.Why don’t ruminal bacteria digest cellulose faster[J].J.Dairy Sci,1996,79,1496 ~ 1502.
[40]Wilson J.R,Kennedy P M.Plant and animal constraints to voluntary feed intake associated with fibre characteristics and particle breakdown and passage in ruminants.Aust[J].J.Agric.Res,1996,47:199 ~ 225.
注:因版面所限僅提供部分參考文獻,如有需要,可來函索取。電子郵箱:aaaaa412@163.com
(譯自2015年第5期Agriclture,778~790頁)
S816.11
A
1004-3314(2017)18-0038-06
10.15906/j.cnki.cn11-2975/s.20171809
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
