
珠芽魔芋葉面球莖的豐產栽培技術
2017-10-23 20:46:50張東華汪慶平
熱帶農業科學 2017年9期
張東華+汪慶平
摘 要 珠芽魔芋植株適宜熱區林下套種,但遮蔭導致高大植株耗去過多養分,葉面球莖數量稀少,繁殖系數受限,制約了種芋的數量。采用多效唑、烯效唑及矮壯素3種矮化劑抑制地下球莖頂芽優勢,促使魔芋植株趨于矮化粗壯,控制魔芋頂芽植株在遮蔭環境條件下過度生長,有效矮化了植株,并使營養物質充分用于葉面球莖中干物質的積累。對于較大的球莖,可將掰除頂芽與噴施化學矮化劑相結合以促進側芽生長和植株矮化,從而達到豐產葉面球莖的目的。對不同重量的繁殖材料,植株矮化率為24%~48%, 葉面球莖數量平均增加率達65%~160%。
關鍵詞 珠芽魔芋 ;矮化 ;葉面球莖 ;繁殖系數
中圖分類號 S632.3 文獻標識碼 A Doi:10.12008/j.issn.1009-2196.2017.09.003
High Yielding Cultural Practices for Producing Bulbils from
Amorphophallus bulbifer by Dwarfing
ZHANG Donghua1) WANG Qingping2)
(1 School of Chemistry & Environment, Yunnan Minzu University, Kunming, Yunnan 650031;
2 Research Institute of Quality Standard and Detection Technology, YAAS,
Kunming, Yunnan 650223)
Abstract Amorphophallus bulbifer is suitable for planting under forests or plantations in the tropical areas, but the shade brings about high uptake of nutrients of the high plants of A. bulbifer and low production of bulbils on the leaves for propagation. A. bulbifer plants were treated with paclobutrazol, uniconazole or chlormequat chloride to inhibit their apical dominance of the corm, which will facilitate their plants to grow dwarf and robust and control their overgrowth under the shade, leading to dwarfing and conversion of the nutrients into dry matter in the bulbils of the leaves. For large corms the apical buds were removed and the corm were sprayed with the chemical agents to allow emergence of lateral buds and dwarf growth of the plants to produce more bulbils on the leaves. For corms of different weight, the treatments produced a plant dwarfing rate of 24% to 48%, and the number of bulbils produced was increased by 65% to 160%.
Keywords Amorphophallus bulbifer ; dwarfing ; bulbils ; propagation coefficient
魔芋是植物中唯一能從其中大量提取葡甘聚糖的植物,僅在亞洲有規模化種植[1]。因種植周期長、繁殖系數低、病害嚴重,傳統種植的魔芋品種屬典型的低產作物。據不完全統計,中國魔芋現有種植面積超過13萬hm2,但年產魔芋精粉僅2.5萬~3萬t,發展數十年產量增長緩慢。盡管如此,中國在全球魔芋市場中排名仍為第一。與花魔芋、白魔芋不同,珠芽類魔芋(A. bulbifer)是染色體屬三倍體,即2n=39的極少數魔芋種[2](約有16個品種),這與國內傳統種植品種花魔芋和白魔芋染色體組為26或28截然不同,植株外觀獨特,植株葉面上長有大小不一的棕色扁球形或長形氣生葉面球莖(bulbils),是主要的繁殖用材料。而葉面球莖獨有的多苗接力生長特性,突破魔芋是獨苗作物的現有概念,可大幅度提高植株的葉面積指數,巧妙延長了植株的生長周期,顯著增加了魔芋的生物產量。前期研究證實[3],該類魔芋的不同繁殖材料生長勢大小順序為:種子>葉面球莖>地下球莖,相應最高膨大倍數分別約為5000∶100∶5~8[4]。盡管種子的膨大率比葉面球莖及地下球莖要大得多,但因其種植技術及所需環境條件要求較高,種植者不易掌握而難以推廣。芋農多習慣選用地下球莖及葉面球莖為繁殖材料。因珠芽魔芋葉面球莖最高膨大系數約達100倍,是地下小球莖膨大率的20倍以上,每公頃用種重量僅為地下球莖的1~3/4,且儲運方便。因此,成本低廉且豐產葉面球莖的方法成為珠芽魔芋生物產量提高的關鍵技術之一。
為解決現有珠芽魔芋在自然生長狀況下出現“頂端優勢”的固有缺陷,可將外源抑制劑與去除頂芽相結合的方法,抑制魔芋組織內過于旺盛的赤霉素分泌,減少母體球莖養分過度用于頂芽新植株葉柄的伸長,控制植株因過快長高長粗而分散養分,使“換頭”后新球莖在營養根長出后,其所吸取的營養物質能在更優化的植株中向利于葉面球莖數量增加的方向轉化,使母體球莖所儲備的“先天”養分及新植株“后續”營養皆可用于植株葉面球莖的生長,以達豐產目的。抑制株高的首要目的是節能,還可抗倒伏及抗低溫,若輔以適量高鉀復合肥,將利于養分充分積淀于葉面球莖上并提高其成熟度,可大幅增加其數量,從而顯著提高珠芽魔芋繁殖系數。endprint
1 林下是喜蔭珠芽魔芋原生棲息地
1.1 魔芋屬喜蔭作物
幾乎所有品種的野生魔芋自然棲息地皆源于林下。40%~60%的遮蔭度被認為是魔芋最佳生長環境。不過,魔芋也能在較大的光照強度范圍內生長。長期以來,花魔芋在日本常被種于中高海拔的山區坡地桑樹下,以減少日光直射時間[5]。據Inaba等[6]報道,50%遮蔭度環境可使花魔芋植株的生長周期由全光照(對照組)的40 d延長至90 d,極利于光合作用的光能轉化進程。盡管出于種植、管理及采收成本多等方面考慮,日本花魔芋種植區現已移至海拔1 000~1 900 m平地或緩坡等光照更充足的環境。若要在低海拔種花魔芋,遮蔭度不足則種植風險極大。Miura等[7]亦發現, 在50%日光透過率條件下,花魔芋塊莖組織中干物質含量隨遮蔭度加大而增加,但當遮蔭度達75%時,花魔芋塊莖重量下降。
1.2 珠芽魔芋尤其需要遮蔭環境
珠芽魔芋適宜在低海拔高溫高濕環境中生長,否則其生物學特性難以充分展現。在緬甸、印尼海拔0~700 m高遮蔭度熱帶雨林,花魔芋難以成活,雜草也明顯受到抑制,但珠芽魔芋卻長勢良好,植株高大[8-9]。 據Santosa等[10]在印尼爪哇島連續3 a的種植試驗表明,完全裸露于陽光照射的條件下,珠芽魔芋植株在第一、第二及第三年種植過程中的枯死率分別高達50%、55%及60%,而當植株處于25%、50%及75%遮蔭度時,均未觀察到葉片出現日灼或植株整體受損的現象。種植試驗顯示,在無遮蔭環境下,魔芋球莖生物產量最低,隨著遮蔭度增加地下球莖個頭亦增大,在遮陰度 75%時達最大,之后球莖生物產量會隨遮蔭度進一步增加呈現下降的趨勢[11],這是由于在較強光照環境中,葉片易出現日灼傷害,降低了光合作用效率,從而致使干物質量減少。與花魔芋不同,缺乏高溫高濕條件的較高海拔區域不適合珠芽魔芋生長,而要滿足其在低海拔生長的氣候特征,適當遮蔭成為不可或缺的環境要素。因此,上坡入林尋找適宜遮蔭環境是種植珠芽類魔芋的理性選擇[12-13],這也能解釋為何在低緯度區域,珠芽魔芋總是棲息于林下。若無高桿作物遮蔭,利用遮陽網通常是種植珠芽魔芋所需的輔助手段。
2 高遮蔭度導致的問題及其控制
2.1 植株易倒伏
魔芋獨特的生長方式是,新植株的長出必然伴隨母體球莖的所謂“換頭”過程,即母體在供養過程中原球莖快速萎縮并與新植株脫離,完成“新老交替”生長循環。在此進程中,因外部環境的溫濕度參數已適宜魔芋頂芽的萌發生長,母體中養分開始快速分解,從而使得魔芋頂芽呈現出生長優勢;而一旦頂芽出土后,新植株的根系所覆蓋的面積亦開始快速延伸,也促使“魔芋筍”葉柄急速向上生長,即使在無遮蔭的環境,魔芋亦多呈現出高植株的外部特征。
高溫高濕條件是珠芽魔芋適宜的生長環境,而遮蔭度的提高必然導致高大植株頻現,葉片的蒸騰作用亦會顯著加快,極易出現失水倒苗現象,尤其當遇到大風天氣時,植株成片倒伏造成生長因此中斷,致使魔芋大面積減產甚至絕收。
2.2 養分分散致葉面球莖數量少、干物質含量低
對珠芽魔芋而言,高植株對應的總是偏小的葉面積,體現在植株上葉面球莖數量稀少且個頭偏小;相反,一些葉柄較細且高僅50~60 cm的植株卻長有較多葉球,有些甚至出現葉柄無法承受過多葉面球莖而倒伏的現象。從能量守恒方面分析,營養物質過多耗費于植株生長,致使用于葉面球莖生長的養分不足,導致葉面球莖數量稀少(一般不超過10枚)且個頭較小,其中往往干物質含量不足,在儲存期間葉面球莖易發生脫水干癟,致使種芋數量進一步受到制約。
2.3 遮蔭度的控制
在0~700 m低海拔高溫高濕環境,魔芋植株具有天生的旺長特性。需在利于魔芋生長的林下遮蔭度(40%~60%)環境下對其進行有效矮化,使植株長勢達最佳狀態,并延長其生長期,最終獲得最大生物產量。若遮蔭度大于75%,植株矮化效果不佳,亦不利于地下球莖生物產量的積累;若位于1 100 m以上海拔環境,珠芽魔芋植株雖可進行凈種而不必遮蔭,但過多的光照仍不利于地下球莖生物產量的積累。
3 珠芽魔芋矮化種植方法
植物的矮化多采用生長延緩劑如多效唑(PP333)、烯效唑(S3307)、矮壯素(CCC)進行,以抑制植株體內赤霉素的生物合成,現已被廣泛用于糧食作物[14-16]及花卉種植中[17],可達到控制植物細胞伸長、縮短節間距、促進分蘗及矮化株高的目的。對珠芽魔芋種植而言,Santosa等[18]采用植物生長激素赤霉素(GA3)刺激植株進行育種,但以化學抑制劑控制株高、促進地下球莖膨大及葉面球莖豐產的相關研究尚未見報道。由于珠芽魔芋繁殖材料多樣,植株矮化方法及特點差異頗大。
3.1 不同種芋應采用差別化矮化方法
3.1.1 針對較大地下球莖(500 g以上)應將去頂抑制與化學抑制結合
在地下球莖儲存期間,陽光的照射會刺激赤霉素的合成,增加球莖開花幾率。要避免球莖開花,宜先進行赤霉素抑制預處理:選擇500 g以下的球莖,將其浸泡于80 mg/L多效唑溶液中10 h;取出后置于溫度為15~35℃的蔭涼干燥處存放,這樣可抑制球莖頂芽過快生長,利于側芽葉芽的萌發及生長。對500 g以上的球莖,須進行人工“打頂”,即將頂芽扳掉再將其浸泡于50 mg/L多效唑溶液10 h。當室外夜間最低溫度高于15℃時即可將球莖下地。
魔芋頂芽優勢過強,大球莖尤其如此。因此,需對500 g以上球莖進行人工“打頂”,以利側芽分化,去頂后的球莖一般會長出3~6棵側芽植株,這是對大球莖母體養分進行人為再分配的結果;當側苗長出后,以多效唑抑制植株葉柄的過度生長,并適時加蓋遮陽網以保護葉片免受日光過度照射;將大球莖按株行距80 cm×80 cm種植于30~35 cm深的土壤穴壇或起垅種植;在魔芋筍出土后的展葉期,以100 mg/L多效唑溶液對整個植株進行噴施,即可抑制植株葉柄的過度生長,得到矮化的利于葉面球莖生長的株型。endprint
3.1.2 針對中小地下球莖(150 g以下)采用化學抑制
選用100~150 g地下球莖作為種芋時,可保留其頂芽,按株行距60 cm×60 cm種植于25~30 cm深的穴壇或起垅種植。化學抑制劑的噴施應在新苗出土展葉時即時噴施,矮壯素噴施濃度為250~300 mg/L。因頂芽出土后生長極快,矮化處理需在葉片展開前即時進行。
3.1.3 以葉面球莖為種芋時在出苗后進行抑制
采用葉面球莖作為種芋時,植株具有2~3苗接力生長特性,前后苗展葉間隔一般為2~3月之久,應在每一苗植株葉片從鱗片伸出但尚未展開前,用100 mg/L的多效唑溶液對整體植株進行噴施,以達預期矮化效果。
3.2 抑制劑種類及其濃度的選擇
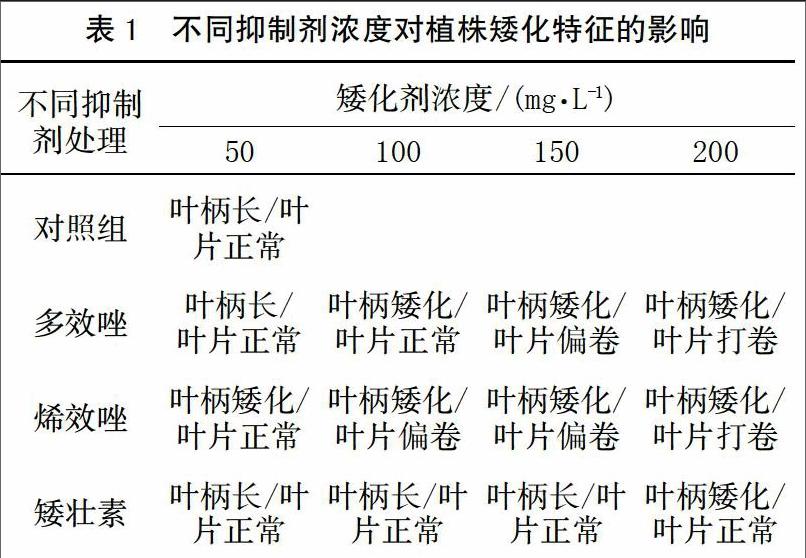
選用多效唑、烯效唑及矮壯素3種化學抑制劑對珠芽魔芋植株進行矮化處理。表1表明,只要以上抑制劑使用濃度適當,植株高度皆可得到有效控制,植株內部營養成分亦可進行轉化,葉面球莖增加所需能源由植株高度受抑制所節省的養分而來。
從表1可知,不同濃度多效唑、烯效唑及矮壯素對魔芋植株的矮化效果差異明顯。其中,多效唑在100 mg/L、烯效唑在50 mg/L即對植株有明顯矮化作用,而矮壯素需在200 mg/L以上方可表現出明顯的抑制效果。矮化劑濃度過高會導致植株葉片打卷,表明抑制過度。
3.3 矮化時機的選擇
魔芋在遮蔭環境下植株具趨光特點,高腳苗呈現較普遍。用化學抑制劑促使頂芽植株盡可能矮化,抑制葉芽葉柄的生長高度,這樣方可利于側芽長出多棵植株,從而實現葉面球莖豐產的目的。矮化劑施用時機十分關鍵,在植株剛出土但葉片尚未伸出鱗片前,植株矮化效果不佳;但亦不能過晚噴施抑制劑,以免因植株葉柄已過高而難以再對其進行抑制。應在葉片伸出鱗片1~2 d但尚未完全展開時即用100~120 mg/L多效唑溶液噴施植株,以防止葉柄因過快伸長而消耗母體球莖養分,有效控制植株高度。
3.4 矮化后葉面球莖生長需輔以施肥
因矮化植株在相對短時期內供葉面球莖生長所需的養分量大,所以需加施足夠農家肥及高鉀復合肥,以利葉面球莖組織中干物質積累,增加飽滿度。
4 植株矮化后所具備的優勢
4.1 植株矮化后抗倒伏效果顯著
魔芋屬淺表根系作物,葉片覆蓋面較大,極易出現倒伏甚至葉柄折斷的現象。在高溫高濕低緯度地區,風災天氣相對頻繁,植株矮化后可顯著降低風災對其造成的損害,對確保足夠的生長周期及生物產量有重要意義。
4.2 植株矮化后葉面球莖數量顯著增加
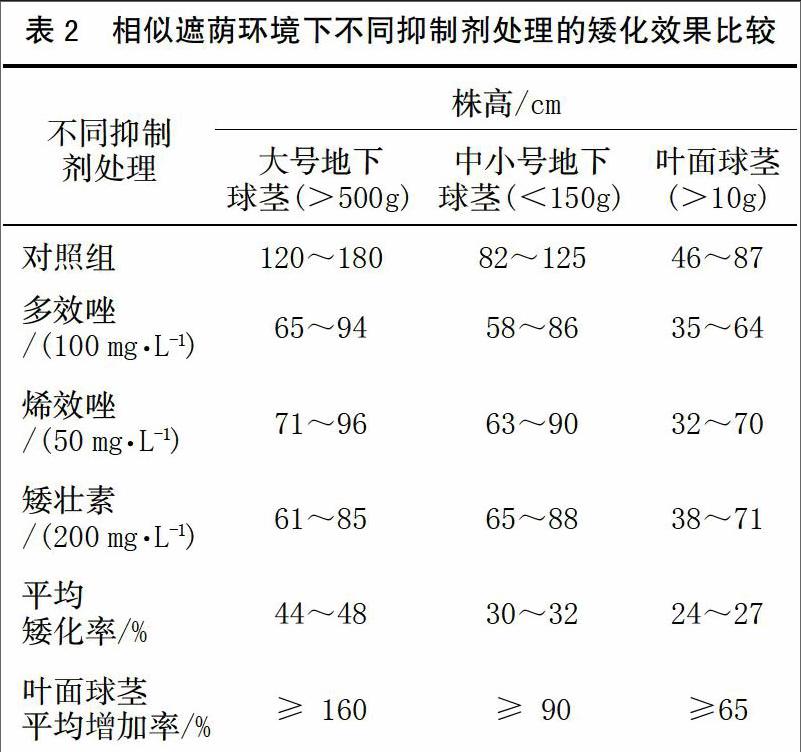
從表2可看出,多效唑、烯效唑及矮壯素3種化學抑制劑皆對珠芽魔芋植株高度有明顯矮化效果;針對同一種材料,不同抑制劑的矮化效果并無明顯差異。與未施用化學抑制劑的植株相比,大號地下球莖(>500 g)矮化效果最明顯,植株平均矮化率為44%~48%,這與球莖先經人工去頂芽有一定關系;中小號地下球莖(<150 g)與葉面球莖(>10 g)矮化率亦分別達30%~32%及24%~27%,表明抑制劑對魔芋植株矮化效果明顯。經與對照組比較,矮化后植株上葉面球莖生長數量皆有大幅增加,以大號地下球莖、中小號地下球莖與葉面球莖為種芋獲得的葉面球莖數量平均增長率約為160%、90%及65%,其中,大號單體球莖平均收獲葉面球莖26~28枚,最多一株長有102枚,葉面球莖單體最大重量為156 g;中小號地下球莖平均收獲葉面球莖8~21枚;以葉面球莖為種芋平均收獲葉面球莖6~15枚。
4.3 延長營養生長期,提高生物產量
在遮蔭環境下,珠芽魔芋的生長周期會明顯長于未遮蔭的對照組植株[7]。采用化學抑制劑矮化后的珠芽魔芋植株為繁殖材料,葉面球莖數量皆會顯著增加,繁殖系數也因此得到提高。種植試驗表明,葉面球莖越多的植株,其地下球莖高度/球莖最大周長亦越大(外觀表現為更飽滿),地下球莖生物產量亦會同比增加。這是由于地面植株在矮化后,在葉面積指數顯著增大的同時,根系也更為發達粗壯,葉柄粗壯,葉片厚度增加,色澤加深為墨綠色,植株總體光合作用效能得到提高,明顯延長了植株有效生長期,可見二者共同作用的結果促使地下球莖生物產量增加。
4.4 矮化植株葉面球莖飽滿、成熟度增加
在印尼、緬甸及中國云南西雙版納林下珠芽魔芋產區,據當地魔芋種植大戶統計,從柚木林、橡膠林下采集的自然脫落葉面球莖,在長達4~5個月的存放期內,平均脫水干癟率約為37%,即1/3以上難以作為種植用種,能作繁殖材料的葉面球莖脫水率也在25%~30%。而經矮化后的魔芋植株,其成熟脫落的葉面球莖飽滿度顯著提高,球莖在經約6個月的儲存后,完全干癟率低于5%,平均脫水率僅在13%~18%,這表明葉面球莖組織中干物質含量明顯增加,組織持水能力有效提高。
5 結論
對珠芽魔芋植株進行化學矮化,以達到大幅度增加葉面球莖數量及其成熟度的目的,能顯著增加魔芋繁殖系數,方便獲得批量生產用種。將植物生長抑制劑與日光照射相結合對地下球莖頂芽進行抑制,用多效唑或唑類植物生長抑制劑降低魔芋球莖內源赤霉素的合成,促使魔芋植株矮化體壯,防止母體球莖養分過度用于新植株葉柄的伸長,控制植株因過快長高長粗而分散植株養分,使新球莖長出的根系所吸取的營養物質能在高度更優化的植株中進行最有利于葉面球莖數量增加及營養物質積累的轉化,將母體球莖所儲備的“先天”養分及新植株產生的“后天”營養向有利于植株葉面球莖生長的方向定向輸送。以唑類化合物作為高效植物生長抑制劑,可促進魔芋植株的矮化粗壯,控制魔芋頂芽植株在遮蔭環境條件下的快速過度生長,即減緩植物頂端分生組織的生長,使細胞伸長變慢,以抑制母體球莖中赤霉素的合成,但不影響葉片的形成,從而有利于植株營養物質在葉面球莖中的快速積累。與對照組比較,矮化后植株上葉面球莖生長數量皆有大幅增加,以大號地下球莖、中小號地下球莖與葉面球莖為種芋獲得的葉面球莖數量平均增長率約為160%、90%及65%。用葉面球莖作為種植用種,植株具有抗性強、耐高溫高濕、優質高產等優勢,且其繁殖系數比傳統白魔芋、花魔芋提高8~10倍以上,改變了現有種植品種花魔芋、白魔芋僅能以小球莖或繁殖根作為種芋的傳統低效途徑,給魔芋種植方式帶來全新突破,方法切實有效,經濟效益好,頗具推廣價值。endprint
參考文獻
[1] 劉佩瑛. 魔芋學[M]. 北京 中國農業出版社,2004.
[2] Hetterscheid W, Ittenbach S. Aroideana[M].International Aroid Society, Inc, 1996: 13-16.
[3] 張東華,汪慶平,楊妹霞. 珠芽魔芋種子5苗接力生長當年形成商品芋技術[J]. 資源開發與市場,2010,26(4):299-301, 318.
[4] Zhang D H, Wang Q P, George S S. Mechanism of staggered multiple seedling production from Amorphophallus bulbifer and Amorphophallus muelleri and its application to cultivation in Southeast Asia[J]. Japanese Journal of Tropical Agriculture, 2010, 54(3): 84-90.
[5] Takeuchi T. History of konjac in Japan [M]. Kodanhsa, Tokyo, 2006: 206.
[6] Inaba K, Chonan N. The effect of light intensity on ultrastructure of chloroplasts in konjak(Amorphophallus konjac K. Koch)[J]. Japanese Journal of Crop Science, 1984, 53(4): 503-509.
[7] Miura K, Osada A. Effect of shading on photosynthesis, respiration, leaf area and corm weight in konjac plants(Amorphophallus konjac K. Koch)[J]. Japanese Journal of Crop Science, 1981, 50(4): 553-559.
[8] 張東華,汪慶平. 緬甸魔芋(Amorphophallus Muelleri)資源分布及產業基本概況[J]. 熱帶農業科學,2013,33(4):46-51.
[9] 張東華,汪慶平,馬晉林. 印度尼西亞食用魔芋(A. muelleri)資源分布及產業發展[J]. 長江蔬菜,2011(14):9-13
[10] Santosa E, Sugiyama N, Nakata M, et al. Growth and corm production of Amorphophallus at different shading levels in Indonesia[J]. Japanese Journal of Tropical Agriculture, 2006, 50: 87-91.
[11] Pushpakumari R, Sasidhar V K. Yield variations of yams and aroids as influenced by shade intensities[J]. Indian Journal of Plant Physiology, 1992, 34: 345-350.
[12] Santosa E, Sugiyama N, Hikosaka S, et al. Cultivation of Amorphophallus muelleri Blume in timber forests of East Java, Indonesia[J]. Tropical Agriculture & Development, 2010,47: 190-197.
[13] Santosa E, Sugiyama N, Hikosaka S, et al. Intercropping practices in cacao, rubber and timber plantation in West Java, Indonesia[J]. Japanese Journal of Tropical Agriculture, 2005, 49: 21-29.
[14] Fujisawa Y, Kato T, Ohki S, et al. Suppression of the heterotrimeric G protein causes abnormal morphology, including dwarfism, in rice[J]. Proceedings of the National Academy of Sciences of the United States of America, 1999, 96 (13): 7 575.
[15] Gao Z Y, Qian Q, Liu X H, et al. Dwarf 88, a novel putative esterase gene affecting architecture of rice plant[J]. Plant Mol Biol, 2009, 71(3): 265-276.
[16] 李文廣,李文欣,師愛英,等. 試論我國小麥矮化育種及其新成就[J]. 安徽農學通報,2008,14(11):78-83.
[17] 潘遠智. 植物生長延緩劑對花卉植物矮化效應的研究[J]. 四川林業科技,2000,21(4):36-39.
[18] Santosa E, Sugiyama N, Nakata M, et al. Flower induction in elephant foot yams using gibberellic acid(GA3)[J]. Tropical Agriculture & Development, 2006, 50: 82-86.endprint
