沙眼衣原體pORF5質粒蛋白抑制腫瘤壞死因子α誘導HeLa細胞凋亡
2017-11-01 21:33:56楊曉玉鄒燕龔思露卜繼常周洲劉良專李忠玉
中華皮膚科雜志 2017年5期
關鍵詞:水平
楊曉玉 鄒燕 龔思露 卜繼常 周洲 劉良專 李忠玉
421001湖南衡陽,南華大學醫學院病原生物學研究所醫學微生物學教研室 特殊病原體防控湖南省重點實驗室
沙眼衣原體pORF5質粒蛋白抑制腫瘤壞死因子α誘導HeLa細胞凋亡
楊曉玉 鄒燕 龔思露 卜繼常 周洲 劉良專 李忠玉
421001湖南衡陽,南華大學醫學院病原生物學研究所醫學微生物學教研室 特殊病原體防控湖南省重點實驗室
目的探討沙眼衣原體質粒蛋白pORF5對腫瘤壞死因子α(TNF-α)誘導HeLa細胞凋亡的影響。方法將含pORF5基因的慢病毒重組表達載體與輔助質粒共轉染293T細胞制備慢病毒,慢病毒收集濃縮后再感染HeLa細胞,流式細胞儀分選獲得pORF5基因穩定轉染細胞株(pORF5-HeLa),同時建立空載體轉染對照細胞株(對照HeLa)。將兩種細胞株分別分為兩組,一組用20 μg/L TNF-α處理(處理組),一組僅用新鮮培養基培養(未處理組),作用6 h,Hoechst33258染色觀察凋亡細胞形態,流式細胞儀檢測細胞凋亡率,實時PCR檢測凋亡相關蛋白Caspase3、Bcl-2和Bax mRNA表達水平,Western印跡檢測Bax、Bcl-2蛋白表達水平。結果TNF-α處理細胞6 h后,Hoechst33258染色發現pORF5-HeLa和對照HeLa細胞中均可見不同程度的核固縮、碎裂,高亮藍色凋亡小體;pORF5-HeLa細胞的凋亡率為(35.5±4.5)%,對照HeLa細胞凋亡率為(63.6±5.8)%,均顯著高于相應未處理組[(9.5±1.5)%和(7.9±0.9)%,t值分別為13.53、32.36,均P<0.01]。處理組pORF5-HeLa細胞中Bax和Caspase3 mRNA表達水平較處理組對照HeLa細胞分別降低72.8%和84.5%(t值分別為35.29,42.25,均P<0.01),但Bcl-2 mRNA的表達水平顯著高于處理組對照HeLa細胞(t=87.12,P<0.01)。處理組pORF5-HeLa細胞中Bax蛋白表達水平亦顯著低于處理組對照HeLa細胞(t=17.58,P<0.01),而Bcl-2蛋白表達水平較對照HeLa細胞增加6.8倍,差異有統計學意義(t=18.93,P<0.01)。結論pORF5可能通過增強抗凋亡蛋白Bcl-2降低促凋亡蛋白Caspase3和Bax的表達,抑制TNF-α誘導的HeLa細胞凋亡。
沙眼衣原體;細胞凋亡;腫瘤壞死因子α;半胱氨酸天冬氨酸蛋白酶3;bcl-2相關X蛋白質;B細胞淋巴瘤/白血病-2蛋白;pORF5質粒蛋白
沙眼衣原體(Chlamydia trachomatis,Ct)是性傳播疾病的主要病原體之一。Ct感染病程隱匿,往往不易被發覺而延誤治療。Ct嚴格細胞內寄生,為從宿主細胞中獲得營養物質并順利完成其在細胞內的發育周期。已有研究報道,衣原體感染可通過絲裂原活化蛋白激酶/細胞外調節蛋白激酶(MAPK/ERK)途徑誘導高遷移率族蛋白1釋放進而抑制細胞凋亡,也可通過阻止線粒體細胞色素C釋放和抑制半胱天冬酶Caspase的活性而抑制細胞凋亡[1-4]。pORF5是由Ct質粒基因編碼并主要定位于感染宿主細胞胞質的一種分泌性效應蛋白[5],既往研究證實pORF5質粒蛋白與Ct致病密切相關[6-11]。為研究pORF5質粒蛋白是否具有抗凋亡作用,本研究將構建成功的慢病毒表達載體和空載體分別與輔助質粒共同包裝重組慢病毒,建立pORF5基因穩定轉染細胞株和對照細胞株,測定腫瘤壞死因子α(TNF-α)作用后凋亡相關蛋白/基因的表達水平,分析pORF5質粒蛋白對HeLa細胞凋亡的影響,為進一步闡明Ct的致病機制提供依據。
材料與方法
一、主要試劑和細胞
慢病毒載體系統(pMDlg-pRRE、pRsv-REV、pMD2G,PLenO-DCE)為上海英為信生物技術有限公司產品;膜聯蛋白Ⅴ-APC/7-AAD(AnnexinⅤ-APC/7-AAD)雙染細胞凋亡檢測試劑盒為美國eBioscience公司產品;RPMI1640為美國HyClone公司產品;LipofectamineTM2000轉染試劑和Trizol總RNA抽提試劑盒為美國Invitrogen公司產品;HeLa細胞、293T細胞株來自上海中國科學院細胞庫。
二、慢病毒的包裝與穩定轉染細胞株的構建
將已構建成功含pORF5基因的重組慢病毒表達載體PLenO-DCE/pORF5和空載體PLenO-DCE分別與pMDlg-pRRE、pRsv-REV、pMD2G輔助質粒共轉染293T細胞48 h,收集病毒上清并進行濃縮與純化,采用倍比稀釋法檢測病毒滴度。將純化后的慢病毒感染HeLa細胞48 h,流式分選技術篩選穩定轉染細胞株,分別命名為pORF5-HeLa細胞和對照HeLa細胞,采用Western印跡對pORF5蛋白的表達進行鑒定。
三、細胞培養及處理
將pORF5-HeLa細胞和對照HeLa細胞接種于6孔細胞培養板,37℃、5%CO2培養箱中培養24 h后分別分成兩組,第1組為處理組:細胞培養基中加入終濃度為20 μg/L的凋亡誘導劑TNF-α培養6 h;第2組為未處理組,直接更換新鮮細胞培養基。實驗重復3次取均值。
四、細胞凋亡檢測
pORF5-HeLa細胞和對照HeLa經上述方法處理后,分別用4%的甲醛固定10 min,PBS洗滌后再加入Hoechst 33258(終濃度為10 mg/L)染色30 min,最后用PBS洗滌、封片,干燥,熒光顯微鏡下觀察。同時收集處理后的pORF5-HeLa細胞和對照HeLa細胞,預冷的PBS緩沖液洗滌后,1×結合緩沖液調整細胞數為1×106個/ml,然后將細胞分成4組,第1組為空白對照組,細胞不加任何試劑染料;第2組細胞加入Annexin V-APC;第3組細胞加入7-AAD,第2組和第3組作為單染對照組;第4組細胞同時加入Annexin V-APC和7-AAD。輕輕混勻并避光孵育15 min,2 h內流式細胞儀分析凋亡率。
五、實時PCR檢測Caspase3、Bcl-2和Bax mRNA表達
收集經TNF-α處理的pORF5-HeLa細胞和對照HeLa細胞,參照Trizol總RNA抽提試劑盒說明書提取細胞總RNA。取1 μg總RNA作為模板,M-MuLV反轉錄酶將RNA反轉錄得到cDNA,以cDNA為模板,應用SYBR Green I熒光染料嵌合法進行實時PCR,測定凋亡相關蛋白Caspase3和Bax mRNA的表達,實驗以甘油醛-3-磷酸脫氫酶(GAPDH)作為內源性對照,引物序列見表1。實時PCR反應條件為94℃預變性10 min,進入94℃變性20 s,65℃退火30 s,72℃延伸45 s,40個循環后,分別測定各種目的基因和內參基因GAPDH的PCR產物熒光強度達到一定閾值所需要的循環數(Ct值),采用2-△△Ct方法計算Caspase3、Bcl-2和Bax基因的相對表達量。
六、Western印跡鑒定Bax和Bcl-2蛋白表達
收集經TNF-α處理的pORF5-HeLa和對照HeLa細胞,PBS洗滌后加入含蛋白酶抑制劑的細胞裂解液充分裂解細胞,離心取上清進行十二烷基磺酸鈉-聚丙烯酰胺凝膠電泳(SDS-PAGE)分析,將分離后的蛋白轉移到二氟化樹脂(PVDF)膜上,PVDF膜經5%脫脂奶粉封閉后,加入兔抗Bax和兔抗Bcl-2抗體,4℃冰箱孵育過夜,0.05%TBST洗滌后加入辣根過氧化物酶(HRP)標記羊抗兔IgG,最后加入ECL顯色,膠片曝光顯影定影。Image Quant軟件分析蛋白質條帶的灰度值,計算蛋白質表達的相對強度。
七、統計學分析
運用SPSS13.0軟件進行統計學分析,所有數據均采用±s表示,兩組間均數比較采用獨立樣本t檢驗,P<0.05為差異有統計學意義。
結 果
一、慢病毒包裝及pORF5基因穩定轉染HeLa細胞株的建立
將慢病毒重組表達載體與輔助質粒共轉染到293T細胞48 h后,由于慢病毒載體攜帶GFP報告基因,熒光顯微鏡下可見細胞發出綠色熒光(圖1)。以不同稀釋度的慢病毒感染293T細胞,熒光顯微鏡下可見隨著病毒濃度的降低,綠色熒光細胞數逐漸減少(圖1A~1D)。根據公式計算出慢病毒滴度為1.5×109TU/ml。慢病毒感染HeLa細胞擴大培養后,流式細胞儀多次篩選獲得穩定轉染細胞株,純度達到95%以上(圖1E)。
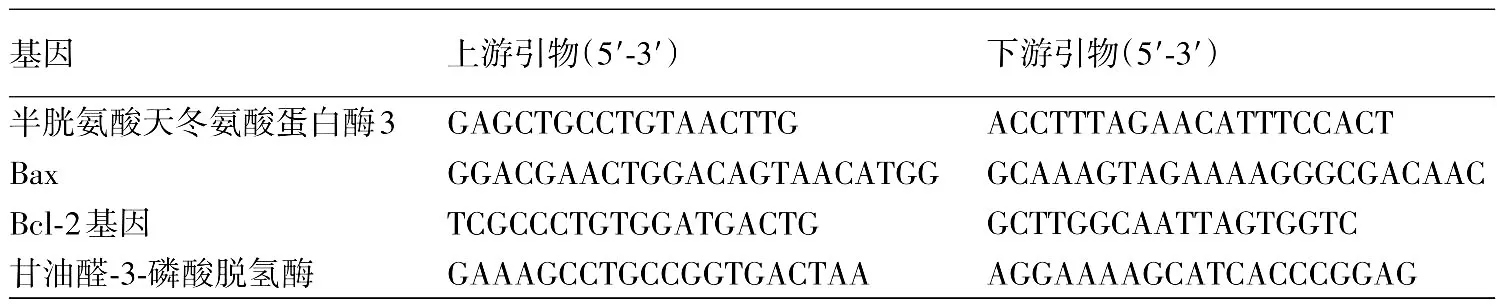
表1 基因實時PCR引物序列
二、穩定轉染HeLa細胞中pORF5質粒蛋白表達的鑒定
Western印跡分析發現,pORF5-HeLa細胞在相對分子質量28 000處出現一條特異性條帶,對照HeLa細胞無條帶出現(圖2)。
三、pORF5對TNF-α誘導HeLa細胞凋亡的影響
pORF5-HeLa和對照HeLa細胞經TNF-α處理后,Hoechst染色結果發現兩組細胞均出現不同程度的核固縮、碎裂,高亮藍色凋亡小體(圖3)。流式細胞儀檢測顯示,處理組pORF5-HeLa和對照HeLa細胞凋亡率分別為(35.5±4.5)%和(63.6±5.8)%,顯著高于相應未處理組[(9.5±1.5)%和(7.9±0.9)%,t值分別為13.53、32.36,均P< 0.01],但TNF-α處理后pORF5-HeLa細胞凋亡率較對照HeLa細胞凋亡率降低28.1%(t=7.05,P<0.01)。見表2。
四、TNF-α處理后Hela細胞Bax、Caspase3和Bcl-2 mRNA表達
由表2可以看出,TNF-α處理后兩種細胞Bax和Caspase3 mRNA水平均顯著高于相應未處理組(均P<0.01);處理組pORF5-HeLa細胞中Bax和Caspase3 mRNA表達水平較處理組對照HeLa細胞分別降低72.8%和84.5%,差異有統計學意義(t值分別為35.29、42.25,均P<0.01),但 Bcl-2 mRNA表達水平顯著高于處理組對照HeLa細胞(t=87.12,P< 0.01)。
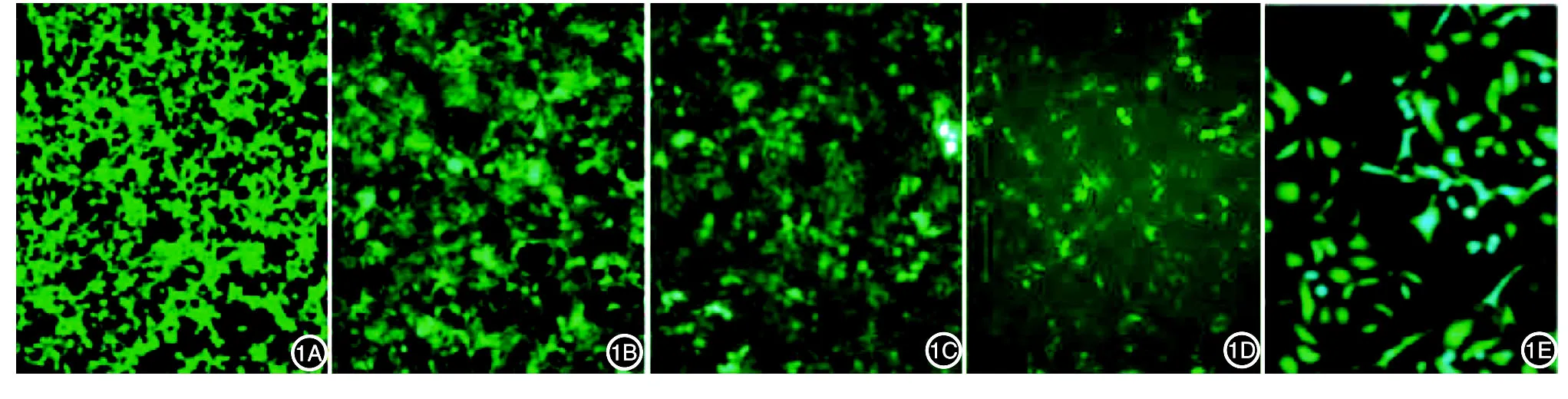
圖1 慢病毒包裝及穩定轉染Hela細胞株的構建 采用倍比稀釋法稀釋慢病毒,然后以不同稀釋度的慢病毒感染293T細胞,熒光顯微鏡下觀察病毒感染情況。1A~1D:分別為慢病毒1∶10、1∶100、1∶1 000、1∶10 000稀釋;1E:穩定轉染的HeLa細胞株
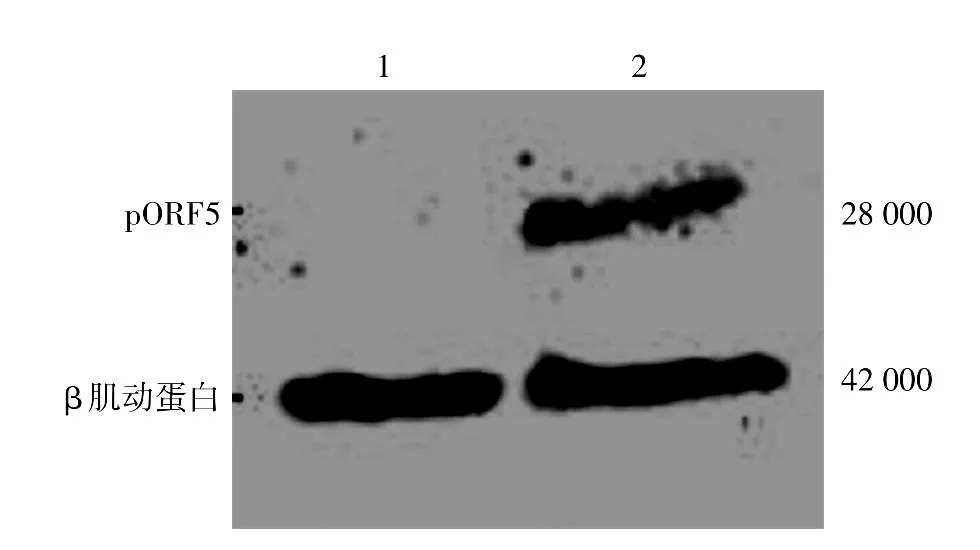
圖2 Western印跡鑒定pORF5蛋白的表達 1:對照HeLa細胞;2:pORF5-HeLa細胞
圖3 Hoechst染色觀察凋亡細胞形態 pORF5-HeLa和對照HeLa細胞經TNF-α處理6h,Hoechst染色觀察細胞凋亡情況,可見兩組細胞均出現不同程度的核固縮、碎裂和高亮藍色凋亡小體等細胞凋亡現象(箭頭)
五、Hela細胞Bax和Bcl-2蛋白表達水平
TNF-α處理后pORF5-HeLa細胞中Bax蛋白表達水平(1.12±0.15)顯著低于對照HeLa細胞組(7.95± 0.62)(t=17.58,P< 0.01),而Bcl-2蛋白表達水平(7.88±0.99)較對照HeLa細胞組(1.01±0.23)增加6.8倍,差異有統計學意義(t=18.93,P<0.01)。見圖4。
討 論
pORF5體外能誘導白介素(IL)-1β、IL-6、腫瘤壞死因子(TNF)-α等多種炎癥因子產生,體內能誘導小鼠生殖道免疫病理損傷[7-10],是Ct重要的毒力蛋白。為探討pORF5質粒蛋白對細胞凋亡的影響,我們用慢病毒系統建立高表達pORF5蛋白的穩定轉染HeLa細胞株。由于慢病毒能夠有效感染非周期性和有絲分裂后的細胞,可實現在細胞中特異而穩定的基因表達,為在細胞中快速而高效地研究pORF5基因功能或pORF5基因表達產物功能提供了有利條件。
細胞凋亡是宿主抗病原體感染的重要防御機制。除了在正常的生長發育和維持組織內穩態方面起重要作用外,細胞凋亡可激活炎癥反應,限制病原體傳播。Ct為一種細胞內寄生菌,為順利完成其在細胞內的發育周期,Ct通過多種機制抑制細胞凋亡[1-4]。為研究pORF5質粒蛋白是否具有抗凋亡作用,本研究用凋亡誘導劑TNF-α刺激pORF5穩定轉染的HeLa和對照HeLa細胞,結果發現,TNF-α能誘導pORF5-HeLa和對照HeLa細胞凋亡,但pORF5-HeLa細胞凋亡率顯著低于對照HeLa細胞,說明pORF5能抑制TNF-α介導的細胞凋亡。
Bcl-2是細胞凋亡信號轉導途徑中關鍵的凋亡調節因子[12],主要通過改變線粒體膜對一些離子、小分子或蛋白質的通透性而參與細胞凋亡的調控。Bax為促凋亡蛋白,當細胞受到凋亡信號刺激后,Bax可誘導線粒體釋放細胞色素C,激活Caspase3,而Caspase3是天冬氨酸特異性半胱氨酸酶家族關鍵的下游成員,Caspase3激活后可引起細胞凋亡。抗凋亡蛋白和促凋亡蛋白的比率決定了細胞對凋亡信號的敏感性[13]。本研究發現TNF-α處理細胞后,pORF5-HeLa細胞中促凋亡因子Bax和Caspase3 mRNA表達水平較對照HeLa細胞顯著減少,Bax蛋白表達水平也顯著低于對照組,而抗凋亡因子Bcl-2蛋白表達水平顯著增加,說明pORF5質粒蛋白能調節凋亡相關蛋白的表達,通過上調抗凋亡蛋白和下調促凋亡蛋白表達,從而實現抗細胞凋亡作用。
我們利用慢病毒載體系統,建立了pORF5基因穩定轉染的HeLa細胞系和對照HeLa細胞系,穩定細胞轉染細胞經TNF-α處理后,發現pORF5可以誘導HeLa細胞中Bcl-2蛋白表達升高,同時降低Bax和Caspase3的mRNA表達水平,初步證實Ct質粒蛋白pORF5可抑制TNF-α介導的細胞凋亡。由于細胞凋亡是多蛋白嚴格控制的過程,pORF5質粒蛋白如何抑制細胞凋亡,凋亡抑制后又如何參與Ct感染性疾病的的發生和發展尚待進一步研究。
表2 兩組HeLa細胞凋亡率以及Bax、Caspase3和Bcl-2基因mRNA表達水平比較(±s)
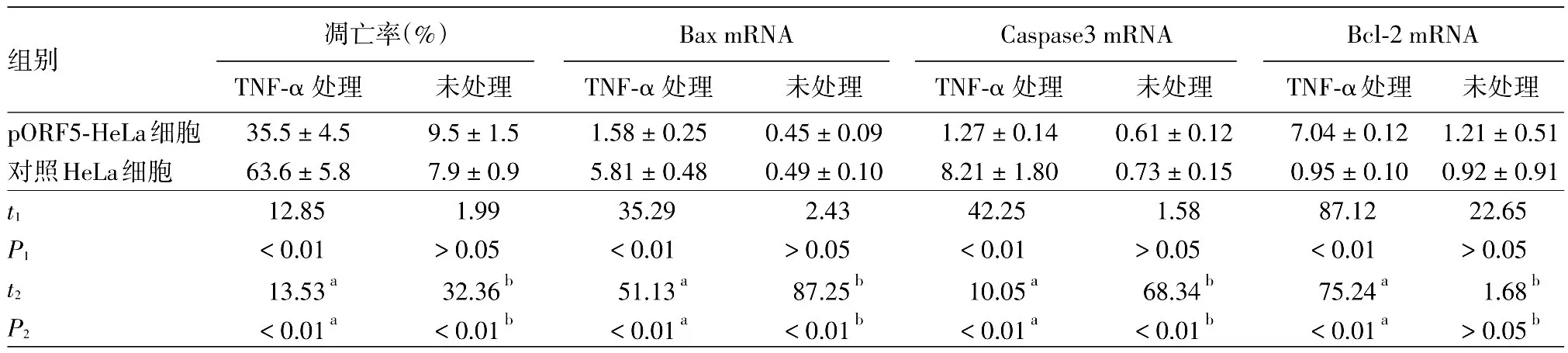
表2 兩組HeLa細胞凋亡率以及Bax、Caspase3和Bcl-2基因mRNA表達水平比較(±s)
注:n=3。t1、P1:pORF5-HeLa細胞與對照HeLa細胞之間比較;t2、P2:同一種細胞TNF-α 處理組與未處理組比較。a:pORF5-HeLa細胞比較結果;b:對照HeLa細胞比較結果。TNF-α:腫瘤壞死因子α
組別pORF5-HeLa細胞對照HeLa細胞t1 P1 t2 P2凋亡率(%)TNF-α 處理35.5±4.5 63.6±5.8 12.85<0.01 13.53a<0.01a未處理9.5±1.5 7.9±0.9 1.99>0.05 32.36b<0.01b Bax mRNA TNF-α 處理1.58±0.25 5.81±0.48 35.29<0.01 51.13a<0.01a未處理0.45±0.09 0.49±0.10 2.43>0.05 87.25b<0.01b Caspase3 mRNA TNF-α 處理1.27±0.14 8.21±1.80 42.25<0.01 10.05a<0.01a未處理0.61±0.12 0.73±0.15 1.58>0.05 68.34b<0.01b Bcl-2 mRNA TNF-α處理7.04±0.12 0.95±0.10 87.12<0.01 75.24a<0.01a未處理1.21±0.51 0.92±0.91 22.65>0.05 1.68b>0.05b
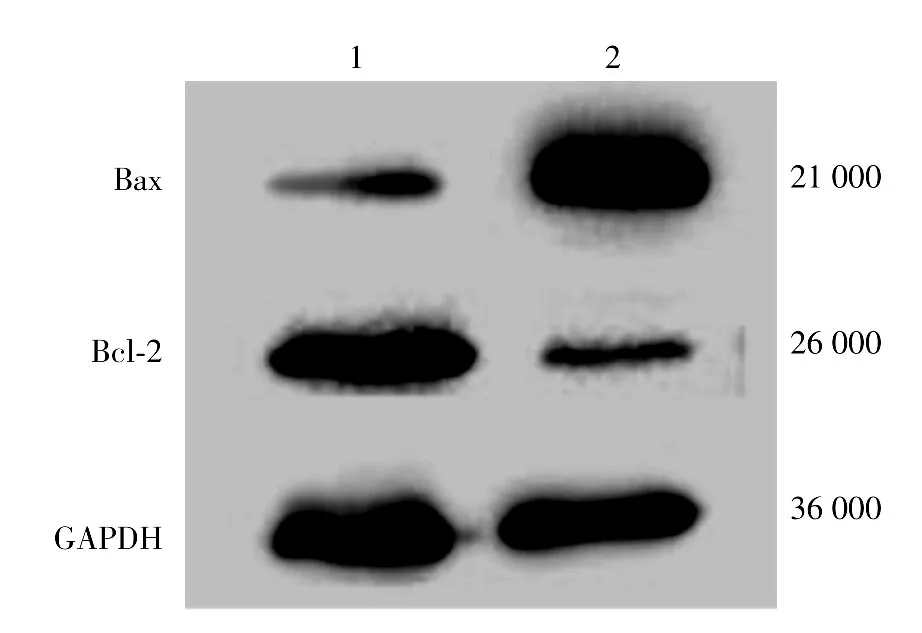
圖4 Western印跡分析Bax和Bcl-2蛋白表達 1:pORF5-HeLa細胞;2:對照HeLa細胞
[1]R?del J,Grosse C,Yu H,et al.PersistentChlamydia trachomatisinfection of HeLa cells mediates apoptosis resistance through aChlamydiaprotease-like activity factor-independent mechanism and induces high mobility group box 1 release[J].Infect Immun,2012,80(1):195-205.DOI:10.1128/IAI.05619-11.
[2]Fan T,Lu H,Hu H,et al.Inhibition of apoptosis in chlamydiainfected cells:blockade of mitochondrial cytochrome c release and caspase activation[J].J Exp Med,1998,187(4):487-496.DOI:10.1084/jem.187.4.487.
[3]Kun D,Xiang-Lin C,Ming Z,et al.Chlamydiainhibit host cell apoptosis by inducing Bag-1 via the MAPK/ERK survival pathway[J].Apoptosis,2013,18(9):1083-1092.DOI:10.1007/s10495-013-0865-z.
[4]Waguia KC,Tzivelekidis T,Gentle IE,et al.Infection of epithelial cells withChlamydia trachomatisinhibits TNF-induced apoptosis at the level of receptor internalization while leaving non-apoptotic TNF-signalling intact[J].Cell Microbiol,2016,18(11):1583-1595.DOI:10.1111/cmi.12598.
[5]Li Z,Chen D,Zhong Y,et al.The chlamydial plasmid-encoded protein pgp3 is secreted into the cytosol of Chlamydia-infected cells[J].Infect Immun,2008,76(8):3415-3428.DOI:10.1128/IAI.01377-07.
[6]Song L,Carlson JH,Whitmire WM,et al.Chlamydia trachomatisplasmid-encoded Pgp4 is a transcriptional regulator of virulenceassociated genes[J].Infect Immun,2013,81(3):636-644.DOI:10.1128/IAI.01305-12.
[7]Liu Y,Huang Y,Yang Z,et al.Plasmid-encoded Pgp3 is a major virulence factor forChlamydia muridarumto induce hydrosalpinx in mice[J].Infect Immun,2014,82(12):5327-5335.DOI:10.1128/IAI.02576-14.
[8]Ramsey KH,Schripsema JH,Smith BJ,et al.Plasmid CDS5 influences infectivity and virulence in a mouse model ofChlamydia trachomatisurogenital infection[J].Infect Immun,2014,82(8):3341-3349.DOI:10.1128/IAI.01795-14.
[9]Zhou H,Huang Q,Li Z,et al.PORF5 plasmid protein ofChlamydiatrachomatisinduces MAPK-mediated proinflammatory cytokines via TLR2 activation in THP-1 cells[J].Sci China Life Sci,2013,56(5):460-466.DOI:10.1007/s11427-013-4470-8.
[10]鄧紅玉,李忠玉,吳移謀,等.沙眼衣原體pORF5質粒蛋白誘發小鼠生殖道免疫損傷初步研究[J].中華微生物學和免疫學雜志,2013,33(2):107-111.DOI:10.3760/cma.j.issn.0254-5101.2013.02.008.
[11]馬康康,李忠玉,粟盛梅,等.pORF5質粒蛋白拮抗LL37抗菌肽增強沙眼衣原體感染的初步研究[J].中華皮膚科雜志,2014,47(10):703-707.DOI:10.3760/cma.j.issn.0412-4030.2014.10.005.
[12]Scarfò L,Ghia P.Reprogramming cell death:BCL2 family inhibition in hematological malignancies[J].Immunol Lett,2013,155(1-2):36-39.DOI:10.1016/j.imlet.2013.09.015.
[13]Ghiotto F,Fais F,Bruno S.BH3-only proteins:the deathpuppeteer′s wires[J].Cytometry A,2010,77(1):11-21.DOI:10.1002/cyto.a.20819.
《中國皮膚性病學書目提要及著者傳略》征文及研修班招生
為了系統總結我國皮膚性病學文獻及介紹做出重要貢獻的著者,廖萬清院士、禤國維國醫大師領銜編輯《中國皮膚性病學書目提要及著者傳略》。凡為該書提供書目及著者傳略者、向籌建的中國皮膚科博物館贈送文物者,可獲贈《中國皮膚科學史》(600元)1冊。
為推廣我國政策規定的外用中藥臨方調劑、中醫中藥療法,2017年繼續舉辦全國皮膚美容化妝品制劑研修班,馬振友、李斌傳授實用處方,示教實習,學會為止。劉巧、劉紅霞等名中醫傳授示教火療、火針、臍療、薰蒸、煙薰、熨烙、鮮藥、刺絡拔罐、太乙神針等實用療法,學員互教互學。報名注冊者獲贈《皮膚美容化妝品制劑手冊》和《皮膚病中醫方劑制劑手冊》各1冊。贈送臨方調配乳膏基質及蒼龍嶺牌等皮膚外用產品試用。對索取征文及招生信息和獲取試用藥品者,來函必復、必贈。
陜西馬振友藥業有限公司,手機/微信:13227015533,13379033002;QQ:386966727。
Inhibitory effect of Chlamydia trachomatis plasmid-encoded protein pORF5 on HeLa cell apoptosis induced by tumor necrosis factor-alpha
Yang Xiaoyu,Zou Yan,Gong Silu,Bu Jichang,Zhou Zhou,Liu Liangzhuan,Li Zhongyu
Institute of Pathogenic Biology,Medical College,University of South China,Hunan Provincial Key Laboratory for Special Pathogens Prevention and Control,Hengyang,421001,China
Li Zhongyu,Email:lzhy1023@hotmail.com
ObjectiveTo evaluate inhibitory effect ofChlamydia trachomatisplasmid-encoded protein pORF5 on HeLa cell apoptosis induced by tumor necrosis factor-alpha(TNF-α).MethodsThe recombinant lentiviral expression vector containing pORF5 gene and helper plasmids were co-transfected into 293T cells to prepare the recombinant lentivirus.Then,the lentivirus particles were collected and concentrated,and used to infect HeLa cells.Flow cytometric screening identified stable pORF5-expressing HeLa(pORF5-HeLa)cells.Meanwhile,the empty plasmid was transfected into HeLa cells to prepare control HeLa cells.The two cell lines were both divided into two subgroups to be treated with 20 μg/L TNF-α and fresh culture medium respectively for 6 hours.Then,Hoechst 33258 staining was performed to observe morphological changes of apoptotic cells,flow cytometry to detect cell apoptosis,real-time PCR to measure the mRNA expression of Caspase3,Bax and Bcl-2,and Western blot analysis to determine the protein expression of Bax and Bcl-2.ResultsAfter 6-hour treatment with TNF-α,Hoechst 33258 staining showed variable degrees of karyopyknosis and karyorrhexis,and highly-refractive blue apoptotic bodies in the pORF5-HeLa cells and control HeLa cells.The pORF5-HeLa cells and control HeLa cells both showed significantly higher apoptosis rate in the treated subgroup than in the untreated subgroup(pORF5-HeLa cells:35.5%±4.5%vs.9.5%±1.5%,t=13.53,P<0.01;control HeLa cells:63.6%±5.8%vs.7.9%±0.9%,t=32.36,P< 0.01).Compared with treated control HeLa cells,treated pORF5-HeLa cells showed significant decreases in mRNA expression of Bax(72.8%)and Caspase 3(84.5%)(t=35.29,42.25,respectively,bothP< 0.01),as well as in Bax protein expression(t=17.58,P< 0.01),but significant increases in Bcl-2 mRNA and protein(6.8 times)expression(t=87.12,18.93,respectively,bothP<0.01).ConclusionpORF5 plasmid protein can inhibit TNF-α-induced HeLa cell apoptosis likely by increasing the expression of anti-apoptotic protein bcl-2 and decreasing the expression of pro-apoptotic proteins Caspase-3 and Bax.
Chlamydia trachomatis;Apoptosis;Tumor necrosis factor-alpha;Caspase3;bcl-2-Associated X Protein;B-cell lymphoma/Leukemia-2;pORF5 plasmid protein
李忠玉,Email:lzhy1023@hotmail.com
10.3760/cma.j.issn.0412-4030.2017.05.007
國家自然科學基金(31470277、81102230、31300156)
Fund program:National Natural Science Foundation of China(31470277,81102230,31300156)
2016-11-25)
(本文編輯:尚淑賢)
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年6期)2019-10-08 08:55:48
人大建設(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(2017年6期)2017-09-26 11:50:44
學苑創造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
