全生育期干旱脅迫對棉花形態、生理、光合作用和產量的影響
2017-11-04 07:12:08田又升范術麗龐朝友孫國清張國麗葉春秀馬盼盼苗領娟謝宗銘
華北農學報 2017年5期
關鍵詞:產量
田又升,范術麗,龐朝友,孫國清,張國麗,葉春秀,于 航,馬盼盼,苗領娟,謝宗銘
(1.新疆農墾科學院 生物技術研究所,作物種質創新與基因資源利用兵團重點實驗室,新疆 石河子 832000;2.棉花生物學國家重點實驗室,河南 安陽 455000;3.中國農業大學 農學院,北京 100093)
全生育期干旱脅迫對棉花形態、生理、光合作用和產量的影響
田又升1,范術麗2,龐朝友2,孫國清1,張國麗1,葉春秀1,于 航1,馬盼盼1,苗領娟3,謝宗銘1
(1.新疆農墾科學院 生物技術研究所,作物種質創新與基因資源利用兵團重點實驗室,新疆 石河子 832000;2.棉花生物學國家重點實驗室,河南 安陽 455000;3.中國農業大學 農學院,北京 100093)
為研究全生育期持續干旱處理對棉花生長發育和最終產量的影響,采用盆栽法,對新陸早19號、新陸早27號和新陸早54號進行輕度和中度干旱脅迫處理,在苗期、蕾期、花鈴期和吐絮期測定形態學指標、生理指標和光合作用參數,收獲時調查產量構成因素。結果表明:棉花的株高和莖粗隨干旱脅迫的加強而下降;活性氧含量和抗氧化酶活性在不同生育期存在較大差異,相關性分析顯示,活性氧含量與抗氧化酶活性間存在顯著或極顯著相關性,說明抗氧化酶活性隨活性氧含量有規律變化,以清除體內過多活性氧,防止活性氧傷害;棉花蕾期光合作用隨干旱脅迫的加強而降低,吐絮期(除新陸早27號)則隨干旱脅迫加強而升高,可能由持續干旱處理使棉花生育后期所處發育階段不同造成;干旱脅迫降低新陸早27號和新陸早54號的單株產量和纖維長度,輕度干旱脅迫對單鈴重的影響較小,甚至起促進作用;棉花花鈴期有4項指標與單株產量存在顯著相關性,說明花鈴期可能是受環境影響最大且決定最終產量的關鍵時期。
棉花;干旱脅迫;生理;光合作用;產量
新疆屬典型大陸性干旱氣候,農業生產完全依賴灌溉,干旱缺水已成為新疆綠洲農業的主要限制因子[1]。棉花(GossypiumhirsutumL.)是抗旱性較強的作物,水分脅迫對其生長和代謝的影響是多方面的。各生育期缺水會使棉花株高、莖粗降低,果枝數、果節數、單株成鈴數減少,鈴期變短,脫落增加[2-4]。干旱脅迫下活性氧大量積累并引發活性氧傷害,植物通過激活酶促保護系統和抗氧化物質以清除過多活性氧[5-8],有效地調節活性氧代謝平衡,降低膜脂過氧化程度,保護膜結構的完整性,增強作物干旱脅迫下的抗氧化能力[9]。綠色植物的光合作用是自然界中規模最大的碳素同化作用,是植物物質生產和產量形成的重要生理過程,同時也是受水分脅迫影響最為顯著的生理過程之一[10]。植物受到干旱脅迫后,最直接的生理反應就是光合速率的降低[11-12]。光合產物的生產總量及其在生殖器官和營養器官的分配決定了作物的產量[13]。棉花是新疆的支柱產業,目前對棉花的研究多集中在某一生育期的生理生化指標分析[14-15],而對水分脅迫下各生育期的指標變化及對最終產量影響方面的研究涉及較少,因此,本試驗以此為切入點,研究全生育期水分脅迫對棉花生長發育及產量的影響,揭示棉花對水分脅迫的生理響應和適應性機制,明確棉花自身需水規律,為節水灌溉的實施提供理論依據。
1 材料和方法
1.1試驗材料
試驗材料為新陸早19號、新陸早27號和新陸早54號,均由新疆農墾科學院生物技術研究所提供。
1.2試驗設計
本試驗在2016年5-12月進行,采用盆栽法在晝/夜溫度為28 ℃/23 ℃,光照周期為14 h/10 h的人工氣候室里進行。將大田土曬干粉碎過篩后,與河沙按6∶1的比例進行混合,裝入上下直徑分別為45,30 cm,高度為35 cm的花盆中,每盆裝土37 kg。在每個花盆中下部加入尿素6 g,磷肥2 g作為底肥。為保證各花盆里的土壤含水量一致,每個花盆均澆水6 L,使花盆里的土完全濕透。待土壤墑合適時,翻松花盆表層土,在花盆中央深度2 cm處播種棉花種子4粒,之后覆膜,出苗后間苗,每個花盆留一株。試驗采用完全隨機設計,包括2個因子和3次重復,第1個因子為不同程度干旱處理,即對照(CK:每次澆水2.0 L),輕度干旱脅迫(LS:每次澆水1.5 L)和中度干旱脅迫(MS:每次澆水1.0 L),第2個因子為3個棉花品種(新陸早19號、新陸早27號和新陸早54號)。
1.3測定指標及方法
分別在苗期、蕾期、花鈴期和吐絮期用卷尺和游標卡尺測定棉花株高和莖粗,采用GFS-3000測定棉花光合作用,為消除人工氣候室內光源對光合作用的影響,本試驗測定光合作用時,光源由儀器提供,光強為600 μmol/(m2·s)。之后采用半葉法取樣,樣品剪碎混勻分裝后,液氮速凍用于生理指標的測定。吐絮期調查棉花始節位、單株鈴數、有效果枝數等農藝性狀,收獲后測定棉花單株產量、纖維長度和衣分等指標。
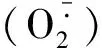
1.4數據分析
試驗數據采用Excel 2007進行整理,Origin 8.0作圖。
2 結果與分析
2.1干旱脅迫對棉花株高和莖粗影響
干旱脅迫降低棉花的株高和莖粗,脅迫強度越大,較對照降低幅度越大。干旱脅迫對棉花株高的抑制在蕾期表現出來,在花鈴期和吐絮期加劇。3個品種中新陸早27號的株高最高,輕度脅迫對其株高影響不顯著,中度脅迫抑制作用明顯。中度脅迫下吐絮期新陸早19號、新陸早27號和新陸早54號的株高分別較對照降低27.69%,33.44%,30.94%(圖1-A)。同一生育期,棉花莖粗在各品種間無明顯差異。干旱脅迫降低棉花莖粗,同一脅迫水平下各品種降低幅度相近(圖1-B)。
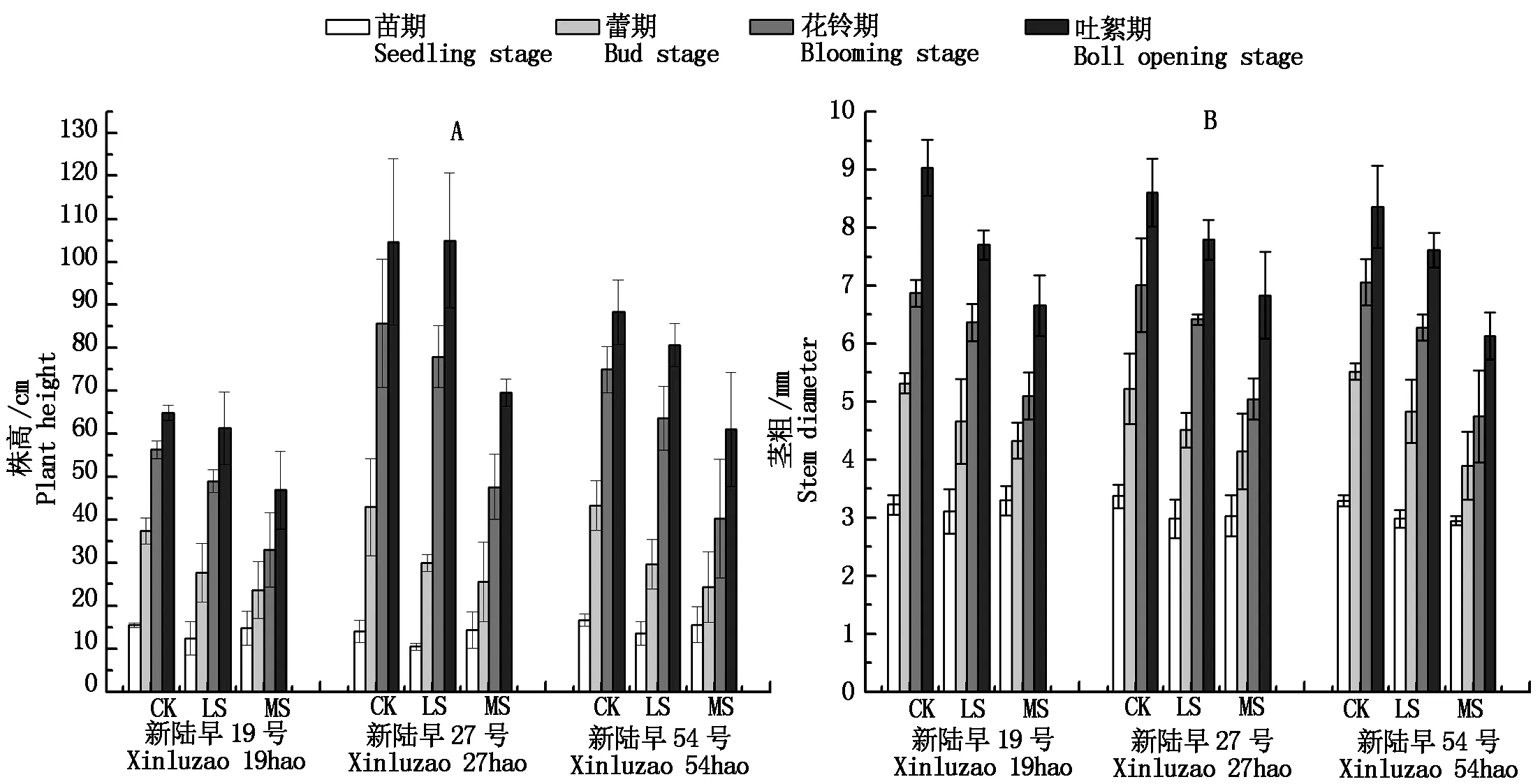
圖1 干旱脅迫對棉花株高和莖粗的影響Fig.1 Effect of drought stress on plant height and stem diameter of cotton
2.2干旱脅迫對棉花膜脂過氧化的影響
干旱脅迫對蕾期新陸早19號和新陸早27號的H2O2含量(以鮮質量計)無顯著影響,新陸早54號在輕度干旱脅迫下降低,較對照降低42.82%,中度干旱脅迫下升高,較對照升高26.17%。花鈴期新陸早19號和新陸早54號 H2O2含量均隨干旱脅迫強度增加而上升,中度脅迫下,新陸早19號、新陸早27號和新陸早54號的H2O2含量分別較對照上升110.55%,91.02%,104.88%。干旱脅迫對吐絮期新陸早19號和新陸早54號的H2O2含量無顯著影響,新陸早27號隨干旱脅迫的加強而降低,輕度和中度脅迫下分別較對照下降18.13%和41.08%(圖2-A)。除了吐絮期新陸早54號的超氧陰離子含量在干旱脅迫下降低(輕度和中度脅迫下分別較對照降低22.60%和16.92%)外,各生育期各品種超氧陰離子含量隨干旱脅迫強度增加而增加(圖2-B)。干旱脅迫提高新陸早19號各生育期的MDA含量(以鮮質量計);新陸早27號苗期和蕾期的MDA含量在中度干旱脅迫下升高,分別較對照上升14.67%和35.91%,花鈴期和吐絮期則下降,分別較對照降低16.56%和9.59%;除花鈴期新陸早54號的MDA含量在輕度脅迫下較對照下降6.16%外,其他時期,干旱脅迫下的MDA含量高于對照(圖2-C)。新陸早54號苗期和蕾期的可溶性蛋白含量隨干旱脅迫的加強而提高;花鈴期和吐絮期,新陸早27號和新陸早54號的可溶性蛋白含量在輕度干旱脅迫下高于對照,中度干旱脅迫下低于對照(圖2-D)。
2.3干旱脅迫對棉花抗氧化系統的影響
棉花APX活性(以鮮質量計)在不同生育期差異較大,在花鈴期的活性最高,苗期最低,因品種和脅迫程度的原因相差20~70倍不等;吐絮期各品種的APX活性在中度干旱脅迫下升高,新陸早19號、新陸早27號和新陸早54號的APX活性在中度脅迫下分別較對照提高50.43%,95.00%,74.26%(圖3-A)。除吐絮期新陸早54號的SOD活性隨干旱脅迫加強而降低外,各品種不同生育期的SOD活性均是在中度干旱脅迫下高于對照,在苗期和蕾期則有規律地隨干旱脅迫的加強而升高(圖3-B)。各品種CAT活性(以鮮質量計)在蕾期最高,除新陸早54號苗期CAT活性和新陸早19號、新陸早27號吐絮期的CAT活性隨干旱脅迫加強而升高外,其他無明顯規律(圖3-C)。各品種的POD活性(以鮮質量計)在干旱脅迫下有規律的變化,在苗期和蕾期隨干旱脅迫的加強而升高,花鈴期和吐絮期則隨干旱脅迫的加強而降低(圖3-D)。
2.4干旱脅迫對棉花光合作用的影響
干旱脅迫對棉花光合作用的影響如表1所示。 在蕾期和花鈴期,棉花各品種葉片SPAD值基本隨干旱脅迫的加強而增加,吐絮期,新陸早27號的SPAD值在脅迫下降低,輕度和中度脅迫下分別較對照低1.89%和0.85%,新陸早19號和新陸早54號的SPAD值則在輕度脅迫下高于對照,分別較對照高出7.81%和6.10%,中度脅迫下低于對照,分別較對照降低2.07%和2.00%。
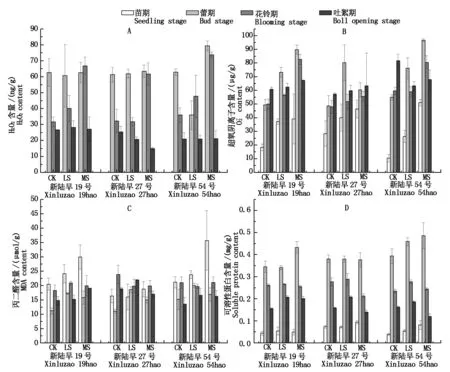
圖2 干旱脅迫對棉花膜脂過氧化的影響Fig.2 Effect of drought stress on membrane lipid peroxidation of cotton
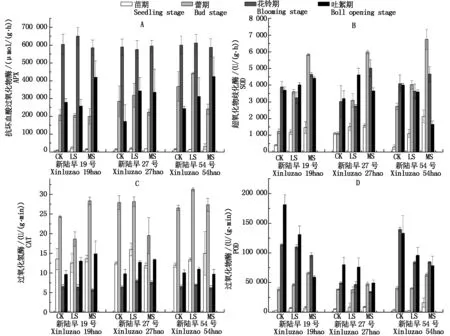
圖3 干旱脅迫對棉花抗氧化系統的影響Fig.3 Effect of drought stress on antioxidant system of cotton
棉花各品種蕾期葉室內相對濕度(Rh)、蒸騰速率(Tr)、氣孔導度(Gs)、光合速率(Pn)、和胞內CO2濃度(Ci)均隨干旱脅迫的加強而下降。花鈴期各品種的葉室內相對濕度隨著干旱脅迫的加強而降低;花鈴期除新陸早27號蒸騰速率在輕度脅迫下降低外,各品種蒸騰速率在干旱脅迫下增加;新陸早19號的氣孔導度在輕度脅迫下升高,較對照升高7.33%,在中度脅迫下降低,較對照下降3.32%,新陸早27號在脅迫下低于對照,輕度和中度干旱脅迫下分別較對照降低14.31%,新陸早54號則是在干旱脅迫下升高,輕度和中度脅迫下分別較對照升高54.18%和26.79%;新陸早19號和新陸早27號花鈴期光合速率和胞內二氧化碳濃度在干旱脅迫下降低,新陸早54號則在中度干旱脅迫下升高,分別較對照升高7.96%和15.27%。吐絮期新陸早19號和新陸早54號的葉室內相對濕度、蒸騰速率、氣孔導度、光合速率均隨干旱脅迫的加強而上升,新陸早19號的胞內二氧化碳濃度隨干旱脅迫的加強而下降;新陸早27號的葉室內相對濕度、蒸騰速率、氣孔導度、光合速率和胞內二氧化碳濃度均在輕度干旱脅迫下最大,分別較對照高出16.37%,55.83%,61.49%,52.55%和24.12%。
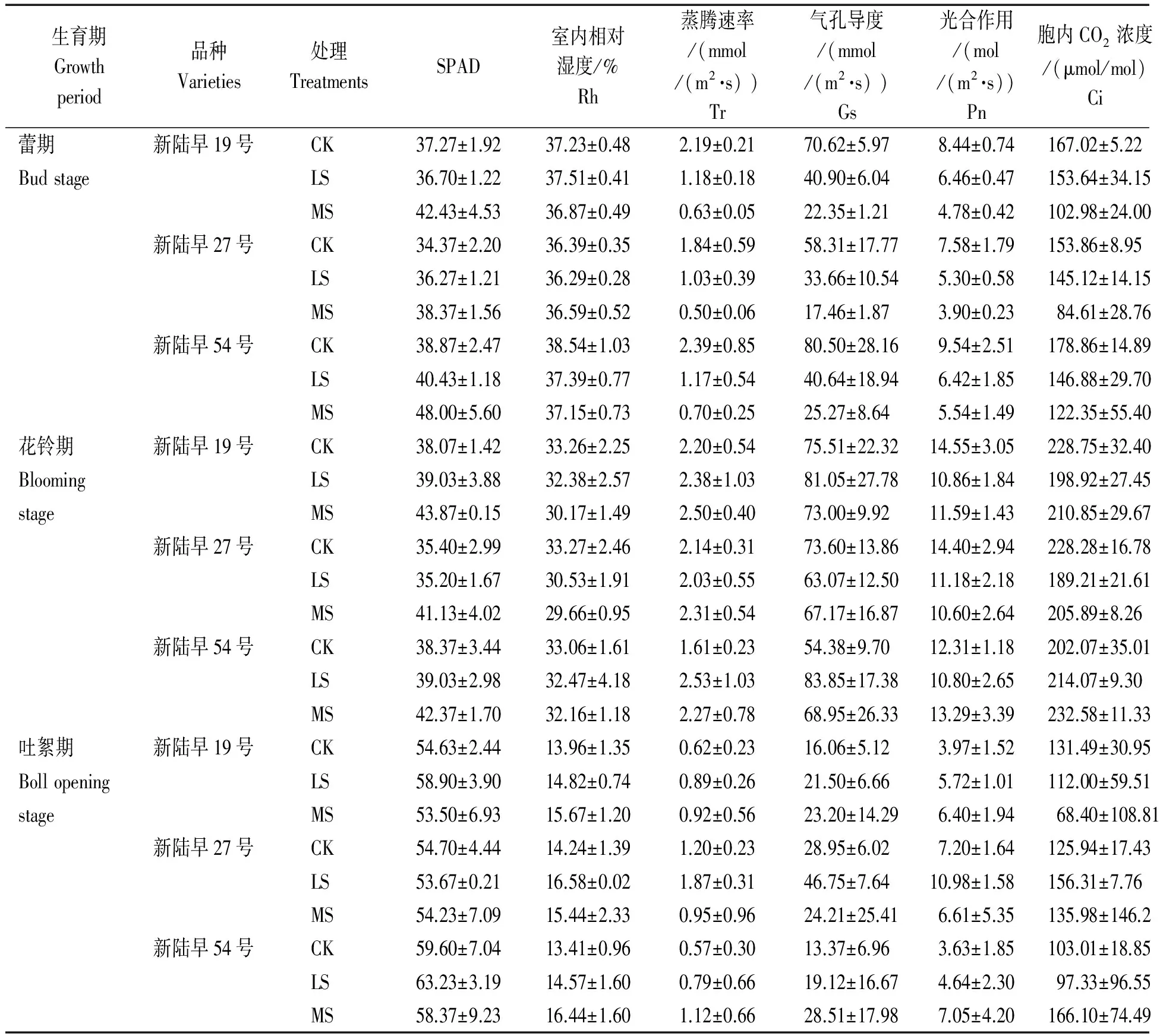
表1 干旱脅迫對棉花光合作用的影響Tab.1 Effects of drought stress on cotton photosynthesis
2.5干旱脅迫對棉花產量構成因素的影響
干旱脅迫對棉花產量構成因素的影響如表2所示。干旱脅迫降低新陸早19號和新陸早27號的第一果枝節位,新陸早54號的第一果枝節位在輕度干旱脅迫最高,較對照和中度脅迫均高出10.50%。新陸早19號和新陸早27號的有效果枝數在輕度干旱脅迫下最高,分別較對照高16.67%和3.41%,新陸早54號則隨干旱脅迫的加強逐漸下降,輕度和中度脅迫分別較對照降低12.00%和27.97%。棉花各品種單株鈴數均隨干旱脅迫的加強而逐漸下降。新陸早19號和新陸早54號的單鈴重在輕度脅迫下最高,分別較對照高32.49%和4.48%,新陸早27號則隨干旱脅迫強度的加強而下降,輕度和中度脅迫分別較對照降低3.09%和8.25%。除中度脅迫下新陸早19號的纖維長度與對照一樣,其余則均是在干旱脅迫下低于對照。干旱脅迫對不同棉花品種衣分的影響存在差異,新陸早19號的衣分在脅迫下增加,輕度和中度脅迫分別較對照高4.74%和1.84%,新陸早27號則隨干旱脅迫強度的增強而增加,輕度和中度脅迫下分別較對照高1.21%和1.69%,新陸早54號則在對照條件下最高,輕度和中度脅迫下分別較對照降低5.94%和4.62%。3個棉花品種中,新陸早27號的單株產量最高,新陸早19號的單株產量在輕度干旱脅迫最高,較對照高22.29%,在中度干旱脅迫下明顯降低,較對照降低57.23%,新陸早27號和新陸早54號則隨干旱脅迫的加強而下降。
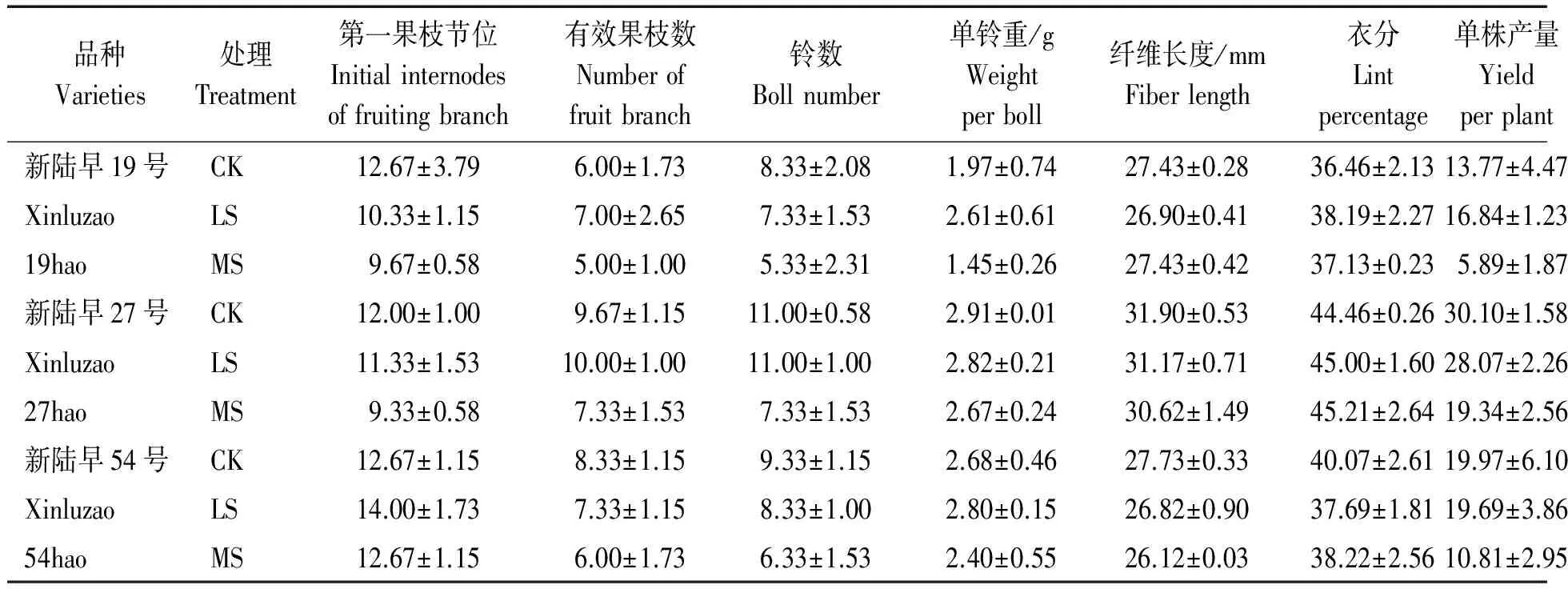
表2 干旱脅迫對棉花產量及其構成因素的影響Tab.2 Effect of drought stress on cotton yield and its component factors
2.6棉花各指標間的相關性分析
將棉花在不同程度干旱脅迫處理下各生育期的指標進行相關性分析(表3)。株高與莖粗間存在極顯著正相關,相關系數為0.93。H2O2含量與CAT活性呈顯著正相關,相關系數為0.44,與POD活性呈顯著負相關,相關系數為-0.41。超氧陰離子含量與SOD活性和POD活性呈極顯著正相關,相關系數分別為0.81和0.53,與APX呈顯著正相關,相關系數為0.46。H2O2含量與超氧陰離子含量間的相關系數為0.35,未達到顯著水平。MDA含量與超氧陰離子含量和抗氧化酶活性間均為負相關,但都未達到顯著水平。光合速率與MDA和APX呈極顯著正相關,相關系數分別為0.63和0.81,與CAT呈極顯著負相關,相關系數為-0.52,與超氧陰離子呈顯著負相關,相關系數為-0.42。
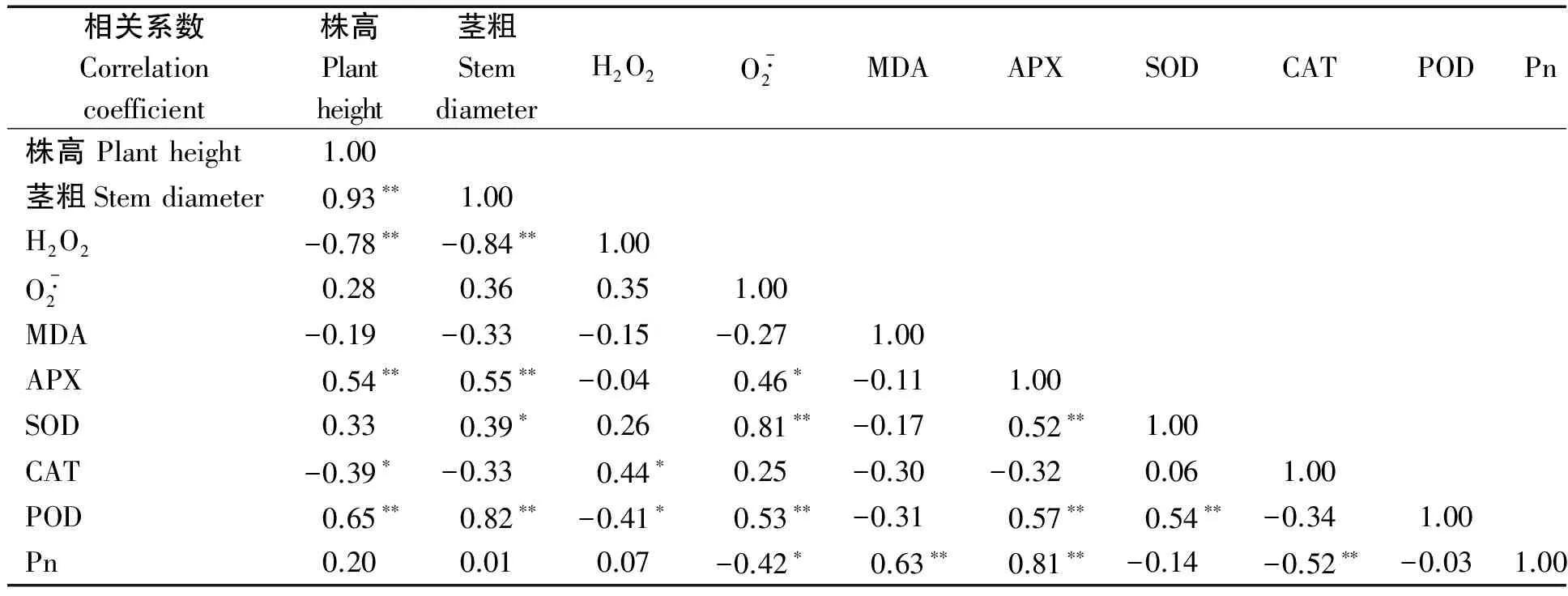
表3 棉花各項指標的相關性分析Tab.3 Correlation analysis of cotton each indexes
2.7棉花產量與各生育期指標間的相關性分析
將各品種在不同水分處理下收獲的單株產量與各生育期的指標進行相關性分析(表4),單株產量與苗期MDA含量呈極顯著負相關,相關系數為-0.82,與蕾期POD活性呈極顯著負相關,相關系數為-0.80,與花鈴期株高呈極顯著正相關,相關系數為0.91,與超氧陰離子含量呈極顯著負相關,相關系數為-0.79,與H2O2和SOD活性呈顯著負相關,相關系數分別為-0.68和-0.71,與吐絮期株高呈極顯著正相關,相關系數為0.95,與APX活性呈顯著負相關,相關系數為-0.67。
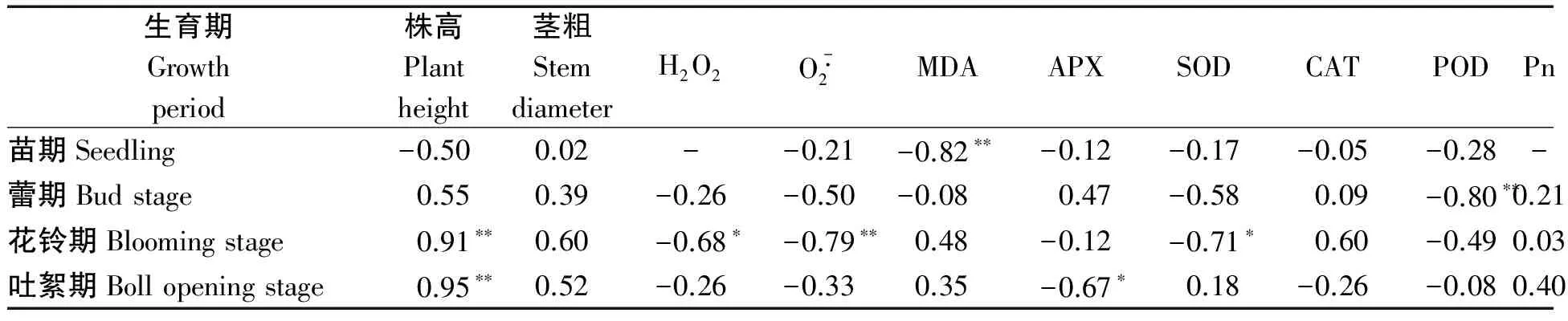
表4 棉花產量與各生育期指標的相關性分析Tab.4 Correlation analysis between cotton yield and index of each growth period
3 討論
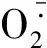
章杰等[28]在研究特早熟棉花光合特性及產量性狀對干旱的適應性時發現籽棉產量、單株鈴數、皮棉產量對水分較為敏感,而單鈴重和衣分變化不大。杜傳莉等[29]總結了干旱對產量構成因素的影響程度為單株成鈴數成鈴率單鈴重,本試驗中,新陸早19號和新陸早54號的單鈴重在輕度干旱脅迫下高于中度干旱脅迫和對照,新陸早27號的單鈴重在輕度干旱脅迫下僅比對照下降3.09%,說明適度干旱脅迫對部分棉花品種單鈴重提高有促進作用。相關性分析顯示,花鈴期測定指標中有4個與單株產量存在顯著或極顯著相關性,而苗期、蕾期和吐絮期各項指標中與單株產量存在顯著相關性的分別為1,1,2個,說明棉花的花鈴期可能是受環境影響最大且決定最終產量的關鍵時期。
本試驗測定的指標中,各品種苗期和蕾期的可溶性蛋白含量、超氧陰離子含量、MDA含量、SOD活性、POD活性在干旱脅迫下增加,其中,個別品種的指標有規律地隨干旱脅迫的加強而升高,光合速率在蕾期隨干旱脅迫的加強而降低,與王娟等[30]的研究結果一致,然而這種規律未出現在花鈴期和吐絮期,吐絮期部分品種的光合速率甚至隨干旱脅迫的加強而提高。前人研究結果表明[31],干旱脅迫加速棉花的生育進程,本試驗的水分處理貫穿棉花整個生育期,各項指標測定在同一時間段進行,干旱脅迫加速棉花生育進程的效果在苗期和蕾期表現不明顯,隨著干旱脅迫的持續,這種效果將加大處在不同水分處理下棉花生育進程的差異,導致花鈴期和吐絮期各項指標測定時,各水分脅迫下的棉花所處生育階段有差異,最終影響棉花花鈴期和吐絮期的試驗結果。在對照吐絮期時,中度干旱脅迫下的棉花已完成吐絮,此時水分處理仍在繼續(對照每澆2 L水時,輕度和中度脅迫分別澆1.5,1.0 L水),少量的水分供給促使干旱脅迫下的棉花在完成吐絮后進行“二次發育”,可能是造成吐絮期光合速率隨干旱脅迫的加強而提高的原因。
[1] 張向娟,張亞黎,張天宇,等.棉花長期干旱后復水葉片光合特性及碳氮含量的變化[J].石河子大學學報:自然科學版,2014,32(2):138-142.
[2] 陳玉梁,石有太,羅俊杰,等.干旱脅迫對彩色棉花農藝,品質性狀和水分利用效率的影響[J].作物學報,2013,39(11):2074-2082.
[3] 羅俊杰,石有太,陳玉梁,等.甘肅不同色彩陸地棉抗旱指標篩選及評價研究[J].核農學報,2012,26(6):952-959.
[4] 陳玉梁,石有太,羅俊杰,等.甘肅彩色棉花抗旱性農藝性狀指標的篩選鑒定[J].作物學報,2012,38(9):1680-1687.
[5] Miao Y,Lv D,Wang P,et al.AnArabidopsisglutathione peroxidase functions as both a redox transducer and a scavenger in abscisic acid and drought stress responses[J].The Plant Cell,2006,18(10):2749-2766.
[6] Miller G,Suzuki N,Ciftci-Yilmaz S,et al.Reactive oxygen species homeostasis and signalling during drought and salinity stresses[J].Plant,Cell & Environment,2010,33(4):453-467.
[7] 田又升,王志軍,于 航,等.干旱脅迫對不同抗旱性棉花品種抗氧化酶活性及基因表達的影響[J].西北植物學報,2015,35(12):2483-2490.
[8] 戴茂華,劉麗英,龐昭進,等.花鈴期干旱脅迫對棉花生理生化指標的影響[J].江西農業學報,2015,27(7):19-21.
[9] Mckersie B D,Chen Y,De B M,et al.Superoxide dismutase enhances tolerance of freezing stress in transgenic alfalfa (MedicagosativaL.)[J].Plant Physiology,1993,103(4):1155-1163.
[10] Chaves M M,Flexas J,Pinheiro C.Photosynthesis under drought and salt stress:regulation mechanisms from whole plant to cell[J].Annals of Botany,2009,103(4):551-560.
[11] Lawlor D W,Tezara W.Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells:a critical evaluation of mechanisms and integration of processes[J].Annals of Botany,2009,103(4):561-579.
[12] Pinheiro C,Chaves M M.Photosynthesis and drought:can we make metabolic connections from available data?[J].Journal of Experimental Botany,2011,62(3):869-882.
[13] 王天澤.光合作用與作物產量[J].植物生理學報,1988(1):54-56.
[14] 田又升,謝宗銘,張建新,等.干旱復水對棉花苗期抗氧化系統及光合熒光參數的影響[J].干旱地區農業研究,2016,34(6):209-214.
[15] 薛惠云,張永江,劉連濤,等.干旱脅迫與復水對棉花葉片光譜,光合和熒光參數的影響[J].中國農業科學,2013,46(11):2386-2393.
[16] 張憲政.植物生理學實驗技術[M].沈陽: 遼寧科學技術出版社,1989:89-98.
[17] Elstner E F,Heupel A.Inhibition of nitrite formation from bydroxyl ammonium chloride:a simple assay for superoxide dismutase[J].Analytical Biochemistry,1976,70(2):616-620.
[18] Cakmak I,Marschner H.Magnesium deficiency and high light intensity enhance activities of superoxide dismutase,ascorbate peroxidase,and glutathione reductase in bean leaves[J].Plant Physiology,1992,98(4):1222-1227.
[19] Wang Y,Wu W H.Plant sensing and signaling in response to K+-deficiency[J].Molecular Plant,2010,3(2):280-287.
[20] Nakano Y,Asada K.Hydrogen peroxide is scavenged by Acerbate-specific peroxidase in spinach chloroplasts[J].Plant and Cell Physiology,1981,22(5):867-880.
[21] 崔慧萍,周 薇,郭長虹.植物過氧化物酶體在活性氧信號網絡中的作用[J].中國生物化學與分子生物學報,2017,33(3):220-226.
[22] Alscher R G,Erturk N,Heath L S.Role of superoxide dismutases (SODs) in controlling oxidative stress in plants[J].Journal of Experimental Botany,2002,53(372):1331-1341.
[23] Shannon L M,Kay E,Lew J Y.Peroxidase isozymes from horseradish roots[J].Journal of Biological Chemistry,1966,241(9):2166-2172.
[24] 程 華,李琳玲,王 燕,等.銀杏過氧化物酶基因POD1的克隆及表達分析[J].華北農學報,2010,25(6):44-51.
[25] Veljovic-Jovanovic S,Kukavica B,Stevanovic B,et al.Senescence- and drought-related changes in peroxidase and superoxide dismutase isoforms in leaves ofRamondaserbica[J].Journal of Experimental Botany,2006,57(8):1759-1768.
[26] Maria H C.Drought stress and reactive oxygen species-production,scavenging and signaling[J].Plant Signaling & Behavior,2008,3(3):156-165.
[27] 馮 晴,徐朗萊.小麥葉片衰老過程中CAT和APX活力及其同工酶譜的變化[J].南京農業大學學報,1997,20(2):95-99.
[28] 章 杰,劉江娜,鄧曉艷,等.干旱對特早熟陸地棉光合特性與產量的影響[J].新疆農業科學,2010,47(7):1397-1401.
[29] 杜傳莉,黃國勤.棉花主要抗旱鑒定指標研究進展[J].中國農學通報,2011,27(9):17-20.
[30] 王 娟,危常州,朱金龍,等.不同生育期干旱脅迫對棉花葉片生理指標及生物量的影響[J].新疆農業科學,2014,51(4):596-604.
[31] 張珠明,葉堯良,陸勇偉,等.浙北棉區棉花不同播期露地直播栽培試驗[J].棉花科學,2014,36(1):22-25.
EffectsofDroughtStressintheWholePeriodonCottonMorphology,Physiology,PhotosynthesisandYield
TIAN Yousheng1,FAN Shuli2,PANG Chaoyou2,SUN Guoqing1,ZHANG Guoli1,YE Chunxiu1,YU Hang1,MA Panpan1,MIAO Lingjuan3,XIE Zongming1
(1.Biotechnology Research Institute,Xinjiang Academy of Agricultural and Reclamation Science,Xinjiang Production & Construction Group Key Laboratory of Crop Germplasm Enhancement and Gene Resources Utilization,Shihezi 832000,China;2.State Key Laboratory of Cotton Biology,Anyang 455000,China;3.College of Agronomy and Biotechnology,China Agricultural University,Beijing 100093,China)
In order to study the effects of continuous drought treatment in whole growth period on cotton growing development and final yield,in this experiment,pot-culture method was taken and Xinluzao 19hao,Xinluzao 27hao and Xinluzao 54hao as materials,the treatments were light and moderate drought stress,then measured morphological indexes,physiological indexes and photosynthetic parameters at seedling stage,bud stage,blooming stage and boll opening stage,and investigated yield components at harvest time.Results showed:cotton height and stem diameter decreased with the strengthen of drought stress; The content of reactive oxygen species and antioxidant enzyme activities were great difference at different growth stage,correlation analysis showed that there were significant or extremely significant correlation between active oxygen content and antioxidant enzyme activity,indicted that antioxidant enzymes activity changed regularly with the content of reactive oxygen species,so as to eliminate the excessive oxygen speciesinvivoand prevent the damage of reactive oxygen species; The photosynthesis of cotton at bud stage decreased with the strengthen of drought stress,but increased at boll opening stage,except Xinluzao 27hao which possibly due to persistent drought treatment caused the different developmental stage of cotton at later growth stage; Drought stress reduced the yield per plant and fiber length of Xinluzao 27hao and Xinluzao 54hao,light drought stress had little effect on single boll weight,even played a role in promoting; There were four indexes of cotton at blooming stage had significant correlation with yield per plant,indicted that cotton blooming stage might be the key period which easy to be affected by environment and determined the final yield.
Cotton; Drought stress; Physiology; Photosynthesis; Yield
2017-08-13
棉花生物學國家重點實驗室開放課題(CB2016A16);兵團國際科技合作計劃項目(2015AH001);兵團應用基礎研究計劃項目(2016AG005)
田又升(1987-),女,重慶涪陵人,助理研究員,碩士,主要從事植物逆境生理與分子生物學研究。
謝宗銘(1971-),男,甘肅民勤人,研究員,博士,碩士生導師,主要從事作物遺傳育種研究。
Q945.78;S562.01
A
1000-7091(2017)05-0224-08
10.7668/hbnxb.2017.05.033
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
今日農業(2021年14期)2021-11-25 23:57:29
今日農業(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
中國化肥信息(2019年2期)2019-01-18 15:24:35
中國化肥信息(2019年1期)2019-01-17 21:31:12
中國化肥信息(2019年4期)2019-01-17 18:47:06
