秦皇島海域野生牙鲆的遺傳多樣性研究
2017-11-06 20:41:25任建功王青林宮春光
河北漁業 2017年10期
任建功+王青林+宮春光

摘 要:采用微衛星標記的方法對秦皇島海域的野生牙鲆進行了遺傳多樣性分析,結果顯示,12對微衛星標記的多態信息含量PIC為0.784~0.949,平均為0.896;12對標記的等位基因數NA為13~108個,平均為51;本研究中觀測雜合度Ho為0.286~0.957,平均為0.764,期望雜合度He為0.809~0.953,平均為0906。結果表明,秦皇島海域野生牙鲆的遺傳多樣性處于較高水平,是優良的牙鲆種質資源寶庫。
關鍵詞:野生牙鲆;微衛星標記;等位基因;遺傳多樣性
向自然水域投放人工培育的苗種是國內外通行的一種恢復漁業種群資源、改善水域生態環境和增加漁業效益的有效手段[1-2]。《農業部關于做好“十三五”水生生物增殖放流工作的指導意見》指出,“十三五”期間將繼續加大對水生生物資源的投入力度,每年增殖重要漁業資源品種的苗種數量達到400億單位以上。牙鲆是渤海重要的增殖放流魚類,我國自20世紀80年代在該海域進行牙鲆的增殖放流活動,對天然漁業資源的恢復起到了積極的作用[3]。但是大規模放流人工培育苗種對自然群體遺傳多樣性的影響受到人們的關注[4]。《農業部辦公廳關于進一步加強水生生物經濟物種增殖放流苗種管理的通知》(農辦漁[2014]55號)明確指出用于繁育增殖放流苗種的親本應來自該物種原產地天然海域、水產種質資源保護區或省級以上原種場保育的原種。因此,在分子水平闡明野生牙鲆遺傳結構,對開展牙鲆增殖放流以及保護牙鲆種質資源具有重要意義。
1 材料與方法
1.1 實驗材料
1.1.1 實驗魚
實驗魚為2015-2016年收購的從秦皇島海域捕獲的野生牙鲆(作為放流牙鲆的親魚),共計231尾,全長57.73±7.18 cm,體重3 737.55±1 231.55 g,其中雌性109尾,雄性122尾。
1.2 實驗方法
1.2.1 樣品采集和DNA提取 剪取野生牙鲆胸鰭平鋪于采樣紙上,37 ℃烘干后保存。使用海洋動物基因組DNA提取試劑盒(TIANGEN,DP324)提取基因組DNA。提取完成后,采用瓊脂糖凝膠電泳和紫外分光光度計分別檢測DNA的質量和濃度。
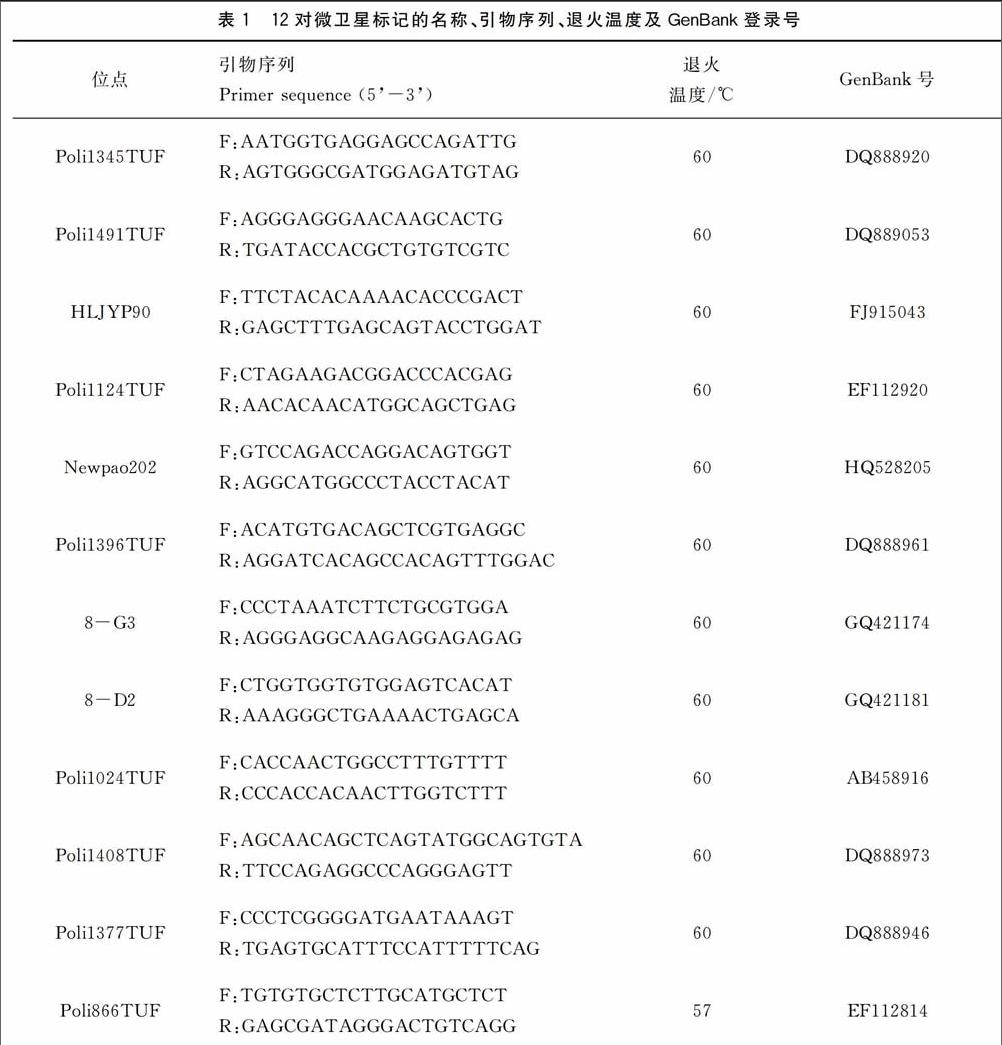
1.2.2 微衛星分析 從馬曉冰等(2012)[5]和劉永新等(2013)[6]發表的72對牙鲆微衛星標記中選取12對多態性高的標記用于本研究。各標記的名稱、引物序列、退火溫度等見表1,引物由上海生工生物工程技術有限公司合成。PCR反應體系為20 μL,包括dH2O 7.4 μL,2×Es Taq MasterMix 10 μL,上下游引物(10 pmol/μL)各0.3 μL,DNA模板2 μL。PCR反應程序為:94 ℃預變性5 min;94 ℃變性30 s,54 ℃復性35 s,72 ℃延伸40 s,共35個循環;最終72 ℃延伸3 min。PCR擴增在PE 9700型PCR儀上進行。將甲酰胺與分子量內標按100∶1的體積比混勻后,取15 μL加入上樣板中,再加入1 μL稀釋10倍的PCR產物。然后使用3730 XL測序儀進行毛細管電泳,利用Genemarker中的Fragment(Plant)片段分析軟件對測序儀得到的原始數據進行分析,將各泳道內分子量內標的位置與各樣品峰值的位置進行比較分析,得到片段大小。
1.3 數據分析
利用Genepop 4.0軟件分析分型結果,統計各位點等位基因數(NA)、觀測雜合度(Ho)、期望雜合度(He)和多態信息含量(PIC),并分析群體的Hardy-Weinberg遺傳平衡狀態。
2 結果和討論
2.1 實驗結果
12對微衛星標記的遺傳多樣性參數NA、Ho、He、PIC值見表2。其中,12對微衛星標記的多態信息含量PIC為0.784~0.949,平均為0. 896;12對標記的等位基因數NA為13~108個,平均為51;本研究中觀測雜合度Ho為0.286~0.957,平均為0.764,期望雜合度He為0.809~0.953,平均為0.906。
2.2 討論
多態信息含量(PIC)為衡量標記遺傳信息含量高低的主要參數,當PIC>0.5時,表明該遺傳標記具有高度的可提供遺傳信息性,即高度多態;當0.25 該研究中平均NA為51,遠高于馬曉冰等[5]和劉永新等[6]對該海域野生牙鲆的研究,說明野生群體的遺傳信息含量非常豐富。雜合度包括觀測雜合度(Ho)和期望雜合度(He),是反映微衛星標記在群體中遺傳變異程度的最優參數[8],是描述遺傳多樣性的最適參數。雜合度越高,表明群體內遺傳多樣性就越高,遺傳變異程度就越大;反之則群體內遺傳變異程度就小[9]。本研究中觀測雜合度Ho平均為0.764,期望雜合度He平均為0.906,平均He遠高于邵長偉等[10]和馬曉冰等[5]研究中的養殖群體和野生群體,說明秦皇島海域野生牙鲆的遺傳多樣性處于較高水平。利用野生牙鲆作為放流親本繁育增殖放流苗種可以改善放流群體的遺傳多樣性。盡管生產中存在牙鲆親本對后代貢獻率失衡的現象,導致放流群體遺傳多樣性低于親本群體[11]。增加繁殖親本的數量,或者采用生殖細胞移植技術可以有效地解決這個問題[12-13]。 哈迪-溫伯格平衡檢驗結果顯示,在12個位點中有8個顯著偏離哈迪-溫伯格平衡狀態(P<0.05)。遺傳不平衡現象在已發表的關于牙鲆群體的研究中普遍存在,可能的原因很多,包括捕撈、放流以及海區環境污染造成野生群體等位基因丟失,無效等位基因,樣本量較小,性別比例失衡,近交衰退,自然選擇等。考慮到偏離哈迪-溫伯格平衡的位點的觀測雜合度和期望雜合度的差異都比較大,我們推測無效等位基因是引起偏分離的主要原因,但是人為原因導致的環境污染、過度捕撈以及苗種培育過程中的近親繁殖等因素同樣不能忽視。
綜上,秦皇島海域野生牙鲆群體遺傳信息含量豐富、遺傳多樣性水平較高,是優良的牙鲆種質資源寶庫,利用野生牙鲆作為放流親魚可以提高放流群體的遺傳多樣性,同時對保護自然種群遺傳多樣性意義重大。
參考文獻:
[1] Seddon PJ,Armstrong DP,Maloney RF.Developing the science of reintroduction biology[J].Conservation Biology,2007,21(2):303-312.
[2] 李繼龍,王國偉,楊文波,等.國外漁業資源增殖放流狀況及其對我國的啟示[J].中國漁業經濟,2009,27(3):111-123.
[3] 金顯仕,邱盛堯,柳學周,等.黃渤海漁業資源增殖基礎與前景[M].北京:科學出版社,2014,1-407.
[4] Taranger GL,Karlsen ,Bannister RJ,et al.Risk assessment of the environmental impact of Norwegian Atlantic salmon farming[J].ICES Journal of Marine Science:Journal du Conseil,2015,72(3):997-1021.
[5] 馬曉冰,王桂興,劉海金,等.秦皇島海域野生牙鲆群體遺傳多樣性分析[J].中國水產科學,2012,19(6):963-969.
[6] 劉永新,朱以美,劉英杰,等.牙鲆渤海自然群體的遺傳多樣性分析[J].水產學報,2013,37(11):1609-1617.
[7] McDonald GM,Wiley J.Space,Time and Life[M].Seattle:University of Washington Press,2003:109-120.
[8] 劉海金,朱曉琛,孫效文,等.牙鲆5個養殖群體的遺傳多樣性分析[J].中國水產科學,2008,15(1):30-37.
[9] Nei M,Maruyama T,Chakraborty R.The bottleneck effect and genetic variability in populations[J].Evolution,1975,29(1):1-10
[10] 邵長偉,廖小林,田永勝,等.牙鲆3個養殖群體遺傳結構的微衛星分析[J].漁業科學進展,2009,30(1):41-46.
[11] 陳睿毅,王桂興,劉海金,等.牙鲆親本對子代貢獻率的實驗研究[J].中國水產科學,2013,20(4):698-705.
[12] Lacerda S,Costa GMJ,Campos-Junior PHA,et al.Germ cell transplantation as a potential biotechnological approach to fish reproduction[J].Fish Physiology and Biochemistry,2013,39(1):3-11.
[13] Silva MA,Costa GMJ,Lacerda S,et al.Successful xenogeneic germ cell transplantation from Jundia catfish (Rhamdia quelen) into adult Nile tilapia (Oreochromis niloticus) testes[J].General and Comparative Endocrinology,2016,230:48-56.endprint
