新疆優質陸地棉資源農藝經濟性狀關聯分析及遺傳構成
2017-11-09 11:41:53路子峰范李萍
西南農業學報 2017年5期
王 莉, 肖 菁*,路子峰,范李萍
(1.新疆農業科學研究院農作物品種資源研究所, 新疆 烏魯木齊 830091;2.新疆農業大學農學院,新疆 烏魯木齊 830091)
新疆優質陸地棉資源農藝經濟性狀關聯分析及遺傳構成
王 莉1, 肖 菁1*,路子峰1,范李萍2
(1.新疆農業科學研究院農作物品種資源研究所, 新疆 烏魯木齊 830091;2.新疆農業大學農學院,新疆 烏魯木齊 830091)
【目的】通過本研究旨在為新疆陸地棉分子標記輔助育種提供理論依據。【方法】關聯作圖也稱連鎖不平衡(Linkage disequilibrium LD)作圖,通過利用性狀與標記之間的相關關系來檢測并定位數量性狀位點。本研究利用94個SSR標記,對64份優質陸地棉資源的基因組進行掃描。在對群體結構分析的基礎上,利用TASSEL軟件的GLM和MLM2種方法對16個農藝經濟性狀進行關聯分析。【結果】①通過STRUCTURE群體結構分析發現這些優質陸地棉種質資源由3個亞群組成,并且這些亞群的劃分與其來源地相關聯,說明地理劃分生態類群有其遺傳基礎。②利用NTSYS基于Nei's 遺傳距離聚類分析發現,64 份陸地棉材料劃分為 3 個亞群,與群體結構劃分結果基本一致。【結論】對16個農藝經濟性狀進行兩種模型的關聯分析,檢測到12個標記均在這兩種模型中出現,其中BNL2448、BNL1404、CGR5108、JESPR158和CIR332這5個標記在2種模型中都檢測到與葉枝數相關聯。
關聯作圖;陸地棉資源;農藝經濟性狀
【研究意義】棉花是我國重要的經濟作物之一,也是世界上重要的自然纖維作物。早熟、產量和纖維品質等性狀是由多基因控制的,屬數量性狀,其遺傳機理復雜且易受外界環境的影響,利用常規育種方法,育種周期長且效率低下。因此利用分子標記輔助育種可以極大的提高育種效率。【前人研究進展】目前,對作物數量性狀的探討大多基于2種途徑:一種是連鎖作圖,另一種就是關聯分析,與連鎖作圖相比,關聯分析有突出的優勢:①不需要專門構建作圖群體,通常以現有的自然群體為試驗對象。②應用廣,對同一座位上的多個基因可以同時檢測。③精度高,可檢測到單基因的差異[1,2]。自2001年,Thornsberry[3]等第一次將關聯分析成功應用于植物研究中,其在植物中的應用也越來越多。最近幾年,隨著測序技術的不斷發展和測序水平的不斷深化,關聯分析技術已經在玉米[4]、水稻[5]、小麥[6]和大麥[7]和大豆[8]中取得一定的成功,受棉花基因組大、多倍性以及序列高度重復的限制[9],棉花關聯分析滯后于其他作物[10-12],但對于陸地棉的研究相對較多, Abdurakhmonov等[13]利用陸地棉資源,包括野生陸地棉、墨西哥棉和來自烏茲別克斯坦的非洲棉共285份進行關聯分析,發現該陸地棉資源具有豐富的遺傳多樣性,群體結構明顯,95個標記中有10~11個標記連鎖不平衡水平顯著,利用MLM模型發掘6 %~13 %的標記與纖維品質性狀相關聯。Abdurakhmonov等[14]對335份陸地棉材料,采用202個SSR標記進行連鎖不平衡和關聯分析,對2個環境的表型基于混合線性模型(MLM)的關聯分析,檢測到平均每個纖維品質性狀與20個SSR標記相關聯。Cai C1等[15]利用97個具有多態性的引物對不同地區的99份陸地棉資源的纖維品質性狀進行關聯分析。結果顯示,檢測到與3個不同環境下的3個纖維品質性狀相關的位點有107顯著性位點,其中與2個或3個環境相關的有70個,僅與一個環境有關的標記有37個。分析70個顯著性位點,發現有52.86 %是已報道的,說明這些是對目標性狀相對穩定的位點。國內研究中,賀道華等[16]和王莉萍等[17]均利用關聯分析方法對棉花纖維品質進行研究,卜愛民[18]對206份耐高溫陸地棉材料的農藝性狀進行關聯分析。【本研究切入點】利用94個SSR共顯性標記,對這64份優質陸地棉資源的基因組進行群體結構分析,并利用TASSEL軟件的GLM和MLM兩種模型對16個農藝經濟性狀進行關聯分析比較,從而得出其農藝經濟性狀更為準確的關聯位點。【擬解決的關鍵問題】通過本研究旨在為新疆陸地棉分子標記輔助育種提供理論依據。
1 材料與方法
1.1 供試材料
試驗材料由64份優質陸地棉資源材料組成(表1),其中包括新疆材料20份,占31.25 %,美國材料14份,21.87 %,其他國外材料共6份,占9.38 %,內地材料10份,占15.63 %,該研究中還篩選了未知來源但纖維品質優良且穩定的材料14份,占21.88 %。
該試驗材料于2010-2012年在新疆庫爾勒巴州農科所試驗地繁殖,常規大田管理,田間試驗按間比法排列, 2行區,行長5 m,株行設置(30 cm+60 cm+30 cm),株距12 cm,2次重復,對照品種(CK)新陸中9號,四周設保護區,采用膜下滴灌。
每個品種隨機調查10株,調查生育期、株高、第一果枝節位、果枝類型、果枝數、葉枝數;并在成熟前期調查株高、株鈴數以、果枝數、果枝始節臺數及高度。成熟期后,每小區收取中部正常吐絮的50個鈴,測定單鈴重、子指及衣分。并由農業部棉花品質監督檢驗測試中心(HVICC標準)測定棉花纖維品質性狀6個:上半部平均長度、斷裂比強度、馬克隆值、伸長率、可紡紗系數、整齊度指數。
1.2 優質陸地棉資源的SSR分子標記
依據改良后CTAB法[19],選用棉花的幼嫩葉片,提取全基因組DNA。先用738對SSR引物對隨機選取的8份試驗材料進行初步的篩選,最終用于試驗的是擴增效果好、條帶差異明顯、背景清晰的94對SSR引物。與He等[20]所用SSR反應體系及程序的一致, PCR (Polymerase chain reaction) 反應體系為 10 μl, 包含10 μmol·L-1的上游引物 1.0 μl,10 μmol·L-1的下游引物 1.0 μl, 模 板 DNA 30 ng,10 mmol·L-1dNTP 0.2 μl ,10×PCR Buffer 1.0 μl, 5 U·μl-1TaqDNA 聚合酶0.14 μl ,用dd H2O補足至 10 μl。 PCR 反應程序為4 min 95 ℃預變性;40 s 94 ℃變性,45 s 52~58 ℃退火,運行33 個循環;8 min 72 ℃延伸;4 ℃保存。產物擴增后處理,利用 8 %非變性聚丙烯酰胺凝膠電泳進行分離,依據張軍等[21]的銀染方法程序對膠片進行拍照保存,最后對分離后的擴增產物進行觀察并統計數據。本實驗在讀取條帶時采用 0、1方法,其中0標記無帶,1標記有帶。
1.3 優質陸地棉資源遺傳結構分析
應用STRUCTURE 2.3.1軟件,根據基因數學模型的原理,對該試驗群體的遺傳結構進行分析,先估算每份資源相對于該群體的Q值(即第i份資源來自于第k群體中的概率),假設群體數(K)在2~8,同時也假定這些位點彼此獨立,其他參數可按照文自翔等[22]的設置,把MCMC(Markov chain monte carlo)中原始的不作數迭代(Length of burn-in period)設置為10 000次,同時也將不作數迭代后的MCMC設為100 000次,再利用最大似然值原則取一個適合的K值,一般計算設20次重復。根據最大似然值原理確定的K值,如果隨K值增大似然值也不斷增大,則利用Evanno 等[23]的方法,用ΔK值取適合K值:
ΔK=m[|L(K+1)-2L(K)+L(K-1)|]/s|L(K)|

表1 64份優質陸地棉種質資源信息表
續表1 Continued table 1

序號No.品種(系)名稱Varieties(lines)name來源Origins品種(系)Genotype系譜Pedigreesource40GK8-2中農院生技中心China'sagriculturalscienceresearchinstitute品系Strain41文⑤(Wen⑤)未知Unknown品系Strain42中棉所49CCRI49中棉所CottonresearchinstituteofCAAS品種Variety中棉所35×中5150443D194未知Unknown品系Strain44ND9807新疆農業大學XinjiangAgriculturalUniversity品系Strain455-1未知Unknown品系Strain46NS029未知Unknown品系Strain479736未知Unknown品系Strain48K-1未知Unknown品系Strain4999-293未知Unknown品系Strain50CK新陸中9號Xinluzhong9新疆庫車XinjiangKuqa品種Variety51新陸中22號Xinluzhong22新疆紅太陽種業XinjiangRedsunindustry品種Variety新陸中4號系選52新陸早21號Xinluzao21新疆農業大學XinjiangAgriculturalUniversity品種Variety新陸中8號×9658(泗棉3號抗病系)新900×貝爾斯諾53新陸中16號Xinluzhong16新疆阿拉爾XinjiangArael品系Strain協作92-36×中棉所1754D256未知Unknown品系Strain55新陸中18號Xinluzhong18新疆天彩科技XinjiangTiancaiScienceandtechnology品系Strain新陸中8號×遼棉10號56新高抗5號Xingaokang5未知Unknown品系Strain57L-14未知Unknown品系Strain58岱字棉55Deltapine55美國America品系Strain59t-22未知Unknown品系Strain6020新疆尉犁XinjiangYuli品系Strain61美BMeiB美國America品系Strain62南6Nan6新疆Xinjiang品系Strain63新陸中34號Xinluzhong34新疆庫爾勒XinjiangKorla品種Variety64長絨67-12Changrong67-12河南安陽HenanAnyang品系Strain8316×中99后代9736株系的自然變異單株
并基于該模型,做出群體遺傳結構圖。
1.4 優質陸地棉資源遺傳多態性聚類
應用NTSYS-pc(Version 1.8)軟件,基于離差平方和的方法以及Nei's 遺傳距離的鄰接法(Neighor-joining,NJ)對94對多態性SSR引物標記進行系統聚類,從而得到該資源群體的結構圖。
1.5 農藝經濟性狀與SSR標記的關聯分析
利用TASSEL軟件中的兩種數學模型:GLM(General linear model)和MLM(Mix linear mode),通過運用個體的Q值作為協變量進行群體結構調整,根據這些優質陸地棉資源16個農藝經濟性狀的3年平均農藝經濟性狀的表現型數據分別與SSR多態性標記位點進行回歸分析。
2 結果與分析
2.1 優質陸地棉資源群體的遺傳結構
對于同一類作物群體來說,均存在的亞群情況,又加上各個亞群的混合,使得群體遺傳結構較為復雜,從而導致整個群體的LD水平增加,進而使目標性狀與不相關的基因座間表現出關聯,因此,在進行關聯分析之前對群體的結構進行分析和處理是必要的[24]。用STRUCTRE2.3.3軟件中混合模型的聚類方法分析這些種質的群體結構,從圖1可以看出,lnP(D)值隨著K值的增加而不斷增大(圖1a)。按照Evanno等提出的方法進一步確定亞群數目。如圖1b所示,ΔK值在K=3和K=4~7中,K=3出現最大值,因此推測該群體有3個亞群。
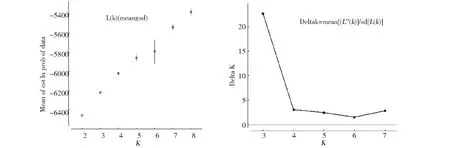
圖1 Structure 模型分析的lnP(D)值(a)和ΔK值(b)Fig.1 The lnP(D)value(a) and ΔK value (b) of structure analysis
利用0.6作為閾值,將所選材料分為3個亞群和一個混合群。
第一亞群包括主要包括新疆品種和內地品種,含有2份美國材料和1份墨西哥材料,共15份,分別有:巴州5628、新庫87317、新庫88209、新庫88251、巴州6501、 Coker 5110、蘭布來特L-X-28、霍皮卡爾、魯棉9號、莘棉5號、CK新陸中9號、GK8-2、文⑤、(CK)新陸中9號、t-22。
第二亞群包括國外材料:6份美國材料和2份國外材料;2份新疆材料和2份未知材料,共12份,分別為: 蘇聯K202、HC4-75、Acala1517-70、lineF、夏費特、新培育品系、 DPL-SOZ #6、9901、中長絨2000-1、D194、D256、岱字棉55。
第三亞群包括新疆品種和內地品種,1份美國品種和8份未知來源地品種,共20份,包括:SicalaV1、140系、MO-78-344、冀92-70、冀棉22號、ND9807、5-1、NS029、9736、K-1、99-293、新陸中22號、新陸早21號、新陸中16號、新陸中18號、新高抗5號、L-14、20南6 、長絨67-12。
混合亞群包括17份材料,包括國內及國外引進材料,分別為:新庫85252、新庫86593、貝爾斯諾 、庫克、Acala1517-77、HC-B4-75、C-83165、毅行2號、SI2、117169-6、冀92-79、新陸中8號、Acala1517-2、97-145、中棉所、美B、新陸中34號。
2.2 聚類分析結果
基于SSR標記的分型數據,利用NTSYS-pc(Version 1.8)軟件中的離差平方和法進行聚類,從而得到UPGMA的樹狀聚類圖(圖2)。在遺傳相似系數為0.734處將這些優質陸地棉資源分為了3大類,與上述分類基本一致,但此方法分類更為詳細。
2.3 與農藝經濟性狀相關聯的SSR標記
2.3.1 利用GLM 程序檢測關聯位點 利用GLM 程序檢測到與其農藝經濟性狀顯著(P<0.01)關聯的 SSR 位點, 如表2所示,只檢測到32個位點與13個性狀相關聯,還有3個性狀籽指、馬克隆值及果枝數沒有關聯到。解釋率最小的是CM45只有8.11 %,與衣分相關聯;最大的33.31 % 是NAU5190與生育期相關聯。有6個標記分別與兩個性狀相關聯,它們是NAU5190、NAU3052、NAU5024、CIR332、BNL2569和DPL0461。BNL2448則與3個性狀相關聯。其中與絨長和葉枝數關聯的位點最多,達到6個標記,解釋率最高分別為22.62 %和21.61 %。橫向比較分析發現,與纖維品質性狀相關的12個標記中有6個位于D亞組染色體上;產量性狀方面,與其相關聯的標記有5個;與農藝性狀相關的標記有12個,與生育期相關的標記最多有3個,其中 NAU5190標記對生育期的解釋率高達33.31 %。
2.3.2 利用MLM 程序檢測關聯位點 利用 MLM 程序檢測到與其農藝經濟性狀顯著(P<0.01)關聯的 SSR 位點(表3),有14個標記與6個性狀相關聯,未檢測到與鈴重、籽指、斷裂比強度、馬克隆值、第一果枝節位、可紡系數、株高、果枝數、單株鈴數和果枝類型10個性狀相關聯的位點。解釋率最小的是衣分的關聯標記NAU5218僅為5.15 %,最大的為與生育期相關聯的標記NAU5190為14.43 %。與葉枝數相關聯的位點最多為5個,分別為BNL1404、BNL2448、CGR5108、JSPR158、CIR332,解釋率最高為13.13 %;其中NAU5190與兩個性狀相關聯,分別是絨長和生育期這2個性狀。
2.3.3 GLM 和 MLM 關聯分析比較 采用的檢測模型不同也會導致不同的結果,本研究中利用MLM模型可檢測到與長度整齊度顯著相關的位點CM3以及與斷裂伸長率顯著相關聯的位點NAU5387,而用 GLM模型則檢測不到。

表2 與農藝經濟性狀顯著(P<0.01)關聯的 SSR 位點(GLM)
能夠用GLM與MLM這2種模型均能檢測到的關聯位點有12個,其中:BNL3582、BNL598、NAU5190與絨長相關聯; NAU5190與整齊度和生育期相關聯;NAU3052則與長度整齊度、株高這2個性狀相關聯;BNL4108與伸長率相關聯;NAU5190、BNL3103與株高相關聯;NAU5218與衣分相關聯;BNL2448、BNL1404、CGR5108、JESPR158、CIR332與葉枝數相關聯。
3 討 論
群體結構的劃分不僅是研究作物種質資源遺傳構成的依據,更是對關聯分析結果產生影響主要因素。研究一個群體內亞群的遺傳結構組成時,會因為亞群的混合而使整個群體的連鎖不平衡水平增強,在這種情況下假陽性結果就可能會增加[22],如果群體結構比較簡單,那么假陽性關聯結果出現的可能性就會減小[25],反之群體結構越復雜則更容易導致這種基因型與表現型之間的假陽性關聯,繼而產生更為復雜的關聯分析結果[26],因此,在進行關聯分析前進行群體結構分析和調節是很有必要的。

表3 與農藝經濟性狀顯著(P<0.01)關聯的 SSR 位點(MLM)
該研究通過比較GLM 和 MLM 2種模型的關聯分析結果,發現MLM模型檢測到的關聯位點明顯比以GLM模型少,這是因為MLM模型是以 Q+K 為協變量,而GLM模型僅以Q為協變量,該結果與 Wang 等[27]結論一致。這說明MLM模型利用Q+K 為協變量可以更多的減少由于群體分層原因引起的假陽性關聯[28]。 關聯得到的位點又以P<0.01 為標準,確定與性狀關聯的 SSR位點,通過這2種原則提高了研究結果的可信度。
根據王榮煥等[29]的論述,全基因組關聯分析指的是對分布于全基因組的高通量標記進行某物種大群體所有基因的掃描,本研究則通過94個SSR標記對這些優質陸地棉資源進行了初步的全基因關聯分析,由于所選群體及所篩選到的標記數量不同,本研究與前人研究相比未發現重合位點,因此可以為分子標記輔助育種提供科學的理論依據。
賀道華等[16]利用132標記對92個棉花種質資源進行棉花纖維品質的關聯分析,檢測到30個位點,其中有5個標記與2個或2個以上的性狀相關聯。王莉萍等[17]利用65個SSR標記對81份陸地棉纖維品質性狀進行研究,結果顯示,檢測到有5個標記并同時與2個或2個以上的性狀相關聯。本研究采用TASSEL軟件的GLM和MLM2種模型,籽指、馬克隆值和果枝數3個性狀均沒有檢測到關聯位點。通過2種模型關聯到葉枝數的位點最多,其中BNL1404在2種模型中均可以檢測到,且解釋率均大于13 %;利用2種模型檢測到與生育期相關的標記是NAU5190,該標記類型是EST-SSR,通過對標記的分析,得知該標記保守性較好,且具有較高的信息量,通過關聯分析,說明EST標記與該性狀連鎖,故猜測該標記可能是直接影響生育期性狀的基因的一個片段。同時發現,利用GLM模型共檢測到7個標記與2個及2個以上性狀相關聯,而通過MLM模型只檢測到1個標記與2個性狀相關聯,該現象可以解釋為性狀之間的相關或者是某一QTL所具有“一因多效”。考慮到本研究所用標記較少,后期試驗可以在該研究基礎上,尤其是2個模型都可以檢測到的位點附件適當的增加標記,從而為今后陸地棉分子標記輔助育種提供依據。
4 結 論
本研究利用TASSEL軟件的2種模型分別進行檢測,GLM模型中檢測到7個標記與2個及2個以上標記相關聯,MLM模型中檢測到1個標記與2個性狀相關聯。該現象可以解釋為性狀之間的相關或者是某一QTL所具有的“一因多效”。目前所用標記較少,因而后期可在該研究的基礎上增加標記,尤其是在2個模型都可以檢測到的位點附加,旨在為今后陸地棉分子標記輔助育種提供更可靠的依據。

圖2 基于 SSR 分子標記的64份優質陸地棉資源聚類圖Fig.2 64 upland cotton germplasm resourse of the clustering map based on SSR markers
[1]Flint-Garcia S A, Thorns berry J M, Buckler E S. Structure of linkage disequilibrium in plants[J]. Annu Rev. Plant Biol.,2003, 54:357-374.
[2]Yu J M, Buckler E S. Genetic association mapping and genome organization of maize[J]. Curr.Opin. Biotechnol.,2006,17:16.
[3]Thornsberry J M, Goodman M M, Doebley J, et al.Dwarf8 polymorphisms associated with variation in flowering time[J]. Nat Genet, 2001,28(3): 286-289.
[4]Belo A, Zheng P, Luck S. Whole genome scan detects an allelic variant offad2 associated with increased oleic acid levels in maize[J].Mol Genet Genomics,2008,279:1-10.
[5]Virk P S, Ford-lloyd B V, Jackson M T, et al. Predicting quantitative variation within rice germplasm using molecular markers[J]. Heredity, 1996, 76(3): 296-304.
[6]Pawan Kulwa ,Goro Ishikawa,David Bendscher, er al.Association mapping for pre-harvest sprouting resistance in white winter wheat[J].Theor Appl Genet,2012,125(4):793-805.
[7]Igartua E, Casas AM, Ciudad F, et al. RFLP markers associated with major genes controlling heading date evaluated in a barley germ plasm pool[J]. Heredity, 1999, 83(5): 551-559.
[8]文字翔.中國栽培和野生大豆的遺傳多樣性、群體分化和演化及其育種性狀QTL的關聯分析[D].南京:南京農業大學,2008.
[9]周仲華,王 峰,陳金湘.棉花基因組學研究進展[C].中國棉花學會,2011:48-50.
[10]Hagenblad J, Nordborg M. Sequence variation and haplotype structure surrounding the flowering time locus FRI inArabidopsisthaliana[J].Genetics, 2002, 161: 289-298.
[11]Tenaillon M I, Sawkins M C, Long AD,et al. Patterns of DNA sequence polymorphism along chromosome 1 of maize (Zeamaysssp. mays L.)[J].Proc. Nat. Acad. Sci., 2001, 98(16): 9161-9166.
[12]Garris A J, McCouch S R, and Kresovich S. Population structure and its effects on haplotype diversity and linkage disequilibrium surrounding the xa5 locus of rice (OryzasativaL.)[J].Genetics, 2003,165: 759-769.
[13]Abdurakhmonov I Y, Kohel R J, Yu J Z, et al. Molecular diversity and association mapping of fiber quality traits in exotic G.hirsutumL. germplasm[J]. Genomics, 2008a,92(6): 478-487.
[14]Abdurakhmonov I Y, Saha S, Jenkins J N, et al. Linkage disequilibrium based association mapping of fiber quality traits in G.hirsutumL.variety germplasm[J].Genetica,2008b,136(3): 401-417.
[15]Cai C1, Ye W, Zhang T, et al.Association analysis of fiber quality traits and exploration of elite alleles in Upland cotton cultivars/accessions (GossypiumhirsutumL.)[J]. Integr Plant Biol., 2014,56(1):51-62.
[16]賀道華,邢宏宜,趙俊興,等.棉花資源群體結構的推測與纖維品質的關聯分析[J].西北農林科技大學學報:自然科學版,2011,39(1):103-112.
[17]王莉萍,孫國清,梁亞軍,等.棉花纖維品質性狀與SSR標記的關聯分析[J].中國農業科技導報,2013,15(4):110-120.
[18]卜愛民. 陸地棉雄蕊發育耐高溫種質資源篩選及農藝性狀與SSR標記關聯分析[D].南京:南京農業大學,2010.
[19]宋國立,崔榮霞,王坤波,等.改良CTAB法快速提取棉花DNA[J].棉花學報,1998,10(5):272-275.
[20]He D H,Lin Z X,Zhang X L,et al.QTL mapping for economic traits based on a dense genetic map of cotton with PCR-based markers using the interspecific cross ofGossypiumhirsutum×G.barbadense[J].Euphytica,2007,153:181-197.
[21]張 軍,張天真,武耀廷,等.棉花微衛星標記的PAGE/銀染快速檢測[J].棉花學報,2000,12(5):267-269.
[22]文自翔,趙團結,鄭永戰,等.中國栽培和野生大豆農藝品質性狀與SSR 標記的關聯分析: I.群體結構及關聯標記[J]. 作物學報, 2008, 34(7): 1169-1178.
[23]Evanno G,Regnaut S,Goudet J.Detecting the number of clusters of individuals using the software STRUCTURE:a simulation study[J].Mol Ecol,2005,14(8):2611-2620.
[24]Flint-Garcia S A, Thuillet AC , Yu J M, et al. Maize association population: a high resolution platform for quantitative trait locus dissection[J]. The Plant Journal, 2005,44(6):1054-1064.
[25]Wang Ju,Mc Clean P E,Lee R,et al.Association mapping of iron deficiency chlorosis loci in soybean(GlycinemaxL.Merr.)advanced breeding lines[J].Theor Appl Genet, 2008,116(6):777-787.
[26]Yu Jianming,Pressoir G,Briggs W H,et al.A unified mixed-model method for association mapping that accounts for multiple levels of relatedness[J].Nat Genet, 2006,38(2):203-208.
[27]Cardon L R,Palmer L J.Population stratification and spurious allelic association[J]. The Lancet in The Plant,2003,361(9375):598-604.
[28]Knowler W C,Williams R C,Pettitt D J,et al.Gm3,5,13,14 and type 2 diabetes mellitus: an association in American Indians with genetic admixture[J].Am J Hum Genet, 1998,43(4):520-526.
[29]王榮煥,王天宇,黎 裕.關聯分析在作物種質資源分子評價中的應用[J].植物遺傳資源學報,2007,8(3):366-372.
AssociationAnalysisofHighQualityUplandCottonResourcesinXinjiangAgriculturalEconomicCharactersandGeneticFormation
WANG Li1,XIAO Jing1*, LU Zi-feng1, FAN Li-ping2
(1. Institute of Crop Germplasm Resource, Xinjiang Academy of Agricultural Sciences, Xinjiang Urumqi 830091,China;2.Agricultural College,Xinjiang Agricultural University, Xinjiang Urumqi 830091,China)
【Objective】This study could provide a theoretical basis for the molecular marker assisted breeding. 【Method】Association mapping, also known as Linkage disequilibrium (Linkage disequilibrium LD), through use of the relationship between traits and markers detected and located quantitative trait locis.This study scanned the genome of 64 upland cotton of high quality materials, using 94 SSR markers. On the basis of the analysis of population structure, TASSEL software had two models, GLM and MLM. which were used to carried out on the 16 agronomic and economic traits. 【Result】 (i)STRUCTURE analysis showed that 64 upland cotton germaplasm resources was composed of three subgroups, which was associated with the place of origin, and confirmed according to the geographical division of ecological groups had its genetic basis.(ii) NTSYS based on Nei's genetic distance cluster analysis showed that 64 upland cotton germaplasm resources was divided into three subsets, consistent basically with structure classification results.【Conclusion】12 markers were detected in the two models, which were associated with 16 agronomic and economic characters, among these markers BNL2448, BNL1404, CGR5108, JESPR158 and CIR332 were detected in the two models, which were associated with leaf branch number.
Association mapping; Upland cotton cultivars; Agronomic and economic traits
1001-4829(2017)5-1000-09
10.16213/j.cnki.scjas.2017.5.004
2016-04-26
新疆農業科學院優秀青年科技人才基金項目 (xjnkq-2013028);新疆維吾爾自治區科研院所改革與發展專項資金項目(KYGY20161242)
王 莉(1976- ),女,山東膠南人,助理研究員,從事農作物種質資源研究, E-mail:wli06@126.com,Tel:18999216489,*為通訊作者。
S562
A
(責任編輯 李 潔)
猜你喜歡
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
當代陜西(2021年17期)2021-11-06 03:21:36
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
學苑創造·A版(2018年11期)2018-02-01 06:29:20
讀者(2017年5期)2017-02-15 18:04:18
山東工業技術(2016年15期)2016-12-01 05:31:22
海峽科技與產業(2016年3期)2016-05-17 04:32:12
