中國(guó)東部5省豬呼腸孤病毒流行病學(xué)調(diào)查與分析
2017-11-14 09:46:40覃盼王經(jīng)緯王斌雷喜梅李龍黃耀偉
覃盼,王經(jīng)緯,王斌,雷喜梅,李龍,黃耀偉*
中國(guó)東部5省豬呼腸孤病毒流行病學(xué)調(diào)查與分析
覃盼1,王經(jīng)緯1,王斌1,雷喜梅1,李龍2,黃耀偉1*1
(1.浙江大學(xué)動(dòng)物科學(xué)學(xué)院動(dòng)物預(yù)防醫(yī)學(xué)研究所,杭州310058;2.杭州貝爾塔生物技術(shù)有限公司,杭州310004)
從核酸和抗體水平對(duì)我國(guó)東部5省豬呼腸孤病毒的流行情況進(jìn)行研究。針對(duì)L1基因設(shè)計(jì)特異性引物,成功建立了豬呼腸孤病毒巢式反轉(zhuǎn)錄聚合酶鏈?zhǔn)椒磻?yīng)(reverse transcription-polymerase chain reaction,RT-PCR)檢測(cè)方法,利用該方法對(duì)2013—2014年從浙江、江西、山東、河南、黑龍江等5省共58個(gè)豬場(chǎng)采集的224份腹瀉豬糞便樣品進(jìn)行檢測(cè)。同時(shí),建立了血清型3型哺乳動(dòng)物呼腸孤病毒(mammalian orthoreovirus,MRV)中和抗體檢測(cè)方法和以病毒衣殼蛋白δ1重組蛋白為包被抗原的間接酶聯(lián)免疫吸附測(cè)定(enzyme-linked immunosorbent assay,ELISA)的血清學(xué)檢測(cè)方法,并利用該方法分別測(cè)定了37份腹瀉豬血清樣品中和抗體滴度和抗σ1 IgG抗體水平。結(jié)果表明,從55個(gè)豬場(chǎng)中共檢出147份陽(yáng)性樣品,豬場(chǎng)陽(yáng)性率和樣品陽(yáng)性率分別為94.8%和65.6%。在37份血清樣本中,中和抗體效價(jià)高于4的有33份(89.2%);而采用間接ELISA方法測(cè)定在450 nm處的吸光度值高于臨界值(0.32)的樣品為32份(86.49%),二者具有較好的相關(guān)性:說明建立的間接ELISA檢測(cè)方法比較可靠。此外,利用建立的間接ELISA方法檢測(cè)了2015—2016年從江西、河南、浙江、黑龍江和山東等省收集的262份具有腹瀉癥狀的母豬、初生仔豬和3周齡斷奶仔豬血清樣品。結(jié)果表明,抗σ1 IgG抗體檢出率為84.4%。根據(jù)核酸和抗體水平檢測(cè)結(jié)果,我們認(rèn)為呼腸孤病毒廣泛存在于中國(guó)東部5省各個(gè)豬場(chǎng)中,應(yīng)該引起足夠的重視。
L1基因;巢式反轉(zhuǎn)錄聚合酶鏈?zhǔn)椒磻?yīng);中和抗體;σ1蛋白;間接酶聯(lián)免疫吸附測(cè)定;流行病學(xué)調(diào)查
近年來,新生仔豬腹瀉持續(xù)在中國(guó)爆發(fā),發(fā)病仔豬呈現(xiàn)腹瀉、嘔吐等癥狀,嚴(yán)重的能導(dǎo)致仔豬迅速脫水、死亡,給養(yǎng)豬產(chǎn)業(yè)帶來了巨大的經(jīng)濟(jì)損失。目前,引起仔豬腹瀉的病原還沒有完全弄清,各國(guó)學(xué)者普遍認(rèn)為豬流行性腹瀉病毒(porcine epidemic diarrhea virus,PEDV)[1]主要以豬傳染性胃腸炎病毒(transmissible gastroenteritis virus,TGEV)[2]、輪狀病毒(rotavirus,RV)[3]等病毒性病原為主導(dǎo),細(xì)菌性病原體和寄生蟲參與其中。近年來,一些新發(fā)腹瀉病毒如豬丁型冠狀病毒(porcine deltacoronavirus,PDCoV)[4]、哺乳動(dòng)物呼腸孤病毒(mammalian orthoreovirus,MRV)[5-11]也受到各國(guó)學(xué)者的關(guān)注。
MRV屬于呼腸孤病毒科(Reoviridae)正呼腸孤病毒屬(Orthoreovirus)的雙鏈RNA病毒,其基因組由10條分節(jié)段的雙鏈RNA組成,根據(jù)10個(gè)節(jié)段大小可分為 L(L1,L2,L3)、M(M1,M2,M3)和 S(S1,S2,S3,S4)3種類型,是一類能引起人和哺乳動(dòng)物呼吸道和消化道疾病的病原。自20世紀(jì)50年代起,先后在人類[12]、狗[13]、牛[14]、水貂[15]、果子貍[16]、蝙蝠[17-18]等哺乳動(dòng)物上被分離到。MRV被證實(shí)不僅能引起人類出血性腸炎、急性呼吸道感染、腦炎等疾病,還能導(dǎo)致其他哺乳動(dòng)物如狗、水貂等腹瀉。但是,關(guān)于豬呼腸孤病毒的研究相對(duì)較少,直到近10年,豬呼腸孤病毒才先后在中國(guó)、韓國(guó)、美國(guó)以及歐洲等地的腹瀉豬糞便樣品中被分離到[5-8,11,19]。NARAYANAPPA等[10]用從美國(guó)分離到的豬呼腸孤病毒感染新生仔豬,發(fā)現(xiàn)仔豬出現(xiàn)嚴(yán)重腹瀉,并且死亡率高達(dá)100%;LELLI等[20]對(duì)牛、豬、馬以及狗等的血清進(jìn)行檢測(cè),發(fā)現(xiàn)抗呼腸孤病毒中和抗體廣泛存在于各種哺乳動(dòng)物中;KWON等[8]在對(duì)韓國(guó)78個(gè)豬場(chǎng)進(jìn)行檢測(cè)時(shí)發(fā)現(xiàn),豬呼腸孤病毒樣品陽(yáng)性率達(dá)到19%。上述報(bào)道說明豬呼腸孤病毒可能廣泛流行于各個(gè)豬場(chǎng)。但是,目前關(guān)于我國(guó)豬呼腸孤病毒的流行情況報(bào)道還比較少。因此,本研究從核酸和抗體水平對(duì)我國(guó)東部5個(gè)省份多個(gè)豬場(chǎng)的豬呼腸孤病毒流行情況進(jìn)行調(diào)查,為全面了解豬呼腸孤病毒在我國(guó)的流行情況提供數(shù)據(jù)支持,同時(shí)對(duì)該病毒的防控也具有指導(dǎo)意義。
1 材料與方法
1.1 試驗(yàn)材料
豬呼腸孤病毒(MRV3-ZJ2013,GenBank登錄號(hào) :KY419120~KY419129)、大 腸 埃 希 菌(Escherichia coli)Rosetta菌株、pET-22b(+)載體,豬流行性腹瀉病毒(PEDV)、豬丁型冠狀病毒(PDCoV)的陽(yáng)性血清由筆者之一黃耀偉實(shí)驗(yàn)室保存。一步法RT-PCR試劑盒(EasyScript one-step RT-PCR SuperMix kit)、2×EasyTaq PCR SuperMix酶以及蛋白純化樹脂(ProteinIso Ni-NTA resin)均購(gòu)自北京全式金公司,限制性內(nèi)切酶購(gòu)自美國(guó)NEB公司,TRizol購(gòu)自美國(guó)Life公司,核酸儀用病毒RNA提取試劑盒購(gòu)自西安天隆科技有限公司,四甲基聯(lián)苯胺(tetramethylbenzidine,TMB)單組分顯色液購(gòu)自北京索來寶公司。
224份腹瀉豬糞便樣品由2013—2014年從浙江、江西、山東、河南、黑龍江等省共58個(gè)豬場(chǎng)收集而來。262份腹瀉豬血清樣本由2015—2016年從上述5省共25個(gè)豬場(chǎng)收集而來,其中86份為母豬血清樣本,95份為1周齡仔豬血清樣本,81份為3周齡仔豬血清樣本。
1.2 豬呼腸孤病毒巢式反轉(zhuǎn)錄聚合酶鏈?zhǔn)椒磻?yīng)(reverse transcription-polymerase chain reaction,RT-PCR)檢測(cè)方法的建立與樣品檢測(cè)
1.2.1 巢式RT-PCR方法建立
比對(duì)分析報(bào)道的豬呼腸孤病毒基因序列,選擇L1基因序列設(shè)計(jì)巢式PCR檢測(cè)引物(以MRV3-ZJ2013毒株為準(zhǔn),GenBank登錄號(hào):KY419120),由杭州博尚生物有限公司合成。利用TRizol抽提MRV3-ZJ2013毒株的基因組RNA作為模板摸索檢測(cè)條件。巢式PCR第1輪利用EasyScript One-Step RT-PCR SuperMix試劑盒擴(kuò)增。反應(yīng)體系為:10 μL 2×One-Step Reaction Mix、0.4 μL TransScriptⅡ One-Step Enzyme Mix、2 μL 總 RNA、正向和反向引物(10 μmol/L)各0.4 μL,加雙蒸水至25 μL。反應(yīng)條件為:50℃反轉(zhuǎn)錄30 min;94℃預(yù)變性5 min;94℃變性30 s,退火30 s,72 ℃ 延伸30 s,共擴(kuò)增35個(gè)循環(huán);72℃延伸10 min。將上一輪獲得的PCR產(chǎn)物稀釋100倍作為模板,利用2×EasyTaq PCR SuperMix酶進(jìn)行第2輪擴(kuò)增。反應(yīng)體系為:12.5 μL 2×EasyTaq PCR SuperMix、1 μL 模板、正向和反向引物(10 μmol/L)各0.5 μL,加雙蒸水至25 μL。反應(yīng)條件為:94℃預(yù)變性5 min;94℃變性30 s,退火 30 s,72℃延伸30 s,共擴(kuò)增35個(gè)循環(huán);72℃延伸10 min。在2輪反應(yīng)中,采用溫度梯度PCR摸索最佳退火溫度,PCR產(chǎn)物經(jīng)過1%瓊脂糖凝膠電泳鑒定。
1.2.2 臨床樣本檢測(cè)
將所采集的具有腹瀉仔豬糞便拭子用300 μL 1×磷酸鹽緩沖液(phosphate buffered saline,PBS)稀釋,反復(fù)凍融3次后,取100 μL用核酸提取儀NP968(西安天隆科技有限公司)按照天隆科技病毒RNA提取試劑盒說明書提取病毒總RNA。用1.2.1節(jié)摸索的最佳條件進(jìn)行巢式PCR檢測(cè),每一輪設(shè)置陰性對(duì)照和陽(yáng)性對(duì)照,其中陽(yáng)性對(duì)照為MRV3-ZJ2013病毒總RNA。
1.3 基于豬呼腸孤病毒δ1蛋白間接酶聯(lián)免疫吸附測(cè)定(enzyme-linked immunosorbent assay,ELISA)方法的建立
1.3.1 δ1蛋白原核表達(dá)及純化
設(shè)計(jì)特異性引物擴(kuò)增S1基因片段(F:5'-GGA ATTCCATATGGATCCTCGTTTACGTG-3';R:5'-CC CAAGCTTCGTGAAACTGCGTGGATA-3'),經(jīng)NdeⅠ和HindⅢ雙酶切后克隆于pET22b(+)載體中,并轉(zhuǎn)化Rosetta感受態(tài)細(xì)胞,經(jīng)菌落PCR篩選出陽(yáng)性克隆并測(cè)序驗(yàn)證。將驗(yàn)證正確的重組菌擴(kuò)大培養(yǎng)至D(600 nm)等于0.6時(shí),用1 mmol/L異丙基-β-D-硫代半乳糖苷(isopropylthio-β-D-galactoside,IPTG)誘導(dǎo)表達(dá)。用Ni-NTA層析柱純化蛋白,并用十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(sodium dodecyl sulfatepolyacrylamide gel electrophoresis,SDS-PAGE)檢驗(yàn)純化。
1.3.2 陰性和陽(yáng)性血清樣本篩選
由分離MRV3-ZJ2013毒株的豬場(chǎng)提供的具有腹瀉癥狀的20份仔豬血清用于篩選陽(yáng)性血清,由本實(shí)驗(yàn)室自行培育的后備母豬所產(chǎn)的10份仔豬血清用于篩選陰性血清。參考本實(shí)驗(yàn)室已建立的方法篩選豬呼腸孤病毒陰性和陽(yáng)性血清樣本[21]。
1.3.3 反應(yīng)條件優(yōu)化
采用棋盤滴定法,確定最佳的抗原包被濃度、血清稀釋度以及二抗稀釋度。用碳酸鹽包被緩沖液(pH 9.6)2倍梯度稀釋?duì)?蛋白,然后包被不同稀釋度蛋白,4℃過夜。每孔用300 μL PBS-T洗滌緩沖液(0.01 mol/L PBS,0.05%Tween-20,pH 7.4)洗滌3次,用PBS-T配制的5%脫脂乳37℃封閉2 h。將 MRV3陽(yáng)性豬血清按照 1∶10、1∶100、1∶200、1∶400、1∶800、1∶1 600稀釋后每孔加入100 μL,每組6個(gè)重復(fù),并在37℃孵育1 h。然后用PBS-T洗滌緩沖液(0.01 mol/L PBS,0.05%Tween-20,pH 7.4)洗滌3次。將按1∶1 000、1∶2 000、1∶4 000、1∶5 000、1∶8 000、1∶10 000等6個(gè)梯度稀釋的HRP標(biāo)記的山羊抗豬Ig G以100 μL/孔分別加入上述6個(gè)板中,37℃孵育30 min。每孔加入100 μL TMB單組分顯色液,并于室溫孵育15 min。最后每孔加入50 μL 2 mol/L硫酸終止反應(yīng),在450nm波長(zhǎng)處讀數(shù)。上述試驗(yàn)重復(fù)3次以確定最佳反應(yīng)條件。
1.3.4 臨界(cut-off)值確定
利用構(gòu)建好的間接ELISA方法測(cè)定陰性血清在450 nm波長(zhǎng)處的吸光度值,并計(jì)算標(biāo)準(zhǔn)差(standard deviation,SD)。臨界值為平均值+3SD。收集的血清樣本在450 nm波長(zhǎng)處的吸光度值若低于臨界值則判定為陰性,若等于或者高于臨界值則判定為陽(yáng)性。
1.3.5 間接ELISA檢測(cè)方法可靠性驗(yàn)證
從待檢血清樣品中隨機(jī)選取37份測(cè)定抗MRV3中和抗體。用DMEM(Dulbecco’s modified Eagle medium)2倍系列稀釋血清樣品,每個(gè)樣品設(shè)置12個(gè)稀釋梯度,每個(gè)梯度8個(gè)重復(fù)。將稀釋的血清放入56℃水浴鍋中滅活30 min。將MRV3-ZJ2013病毒液用維持培養(yǎng)基(maintenance medium,MM)稀釋到100 TCID50(50%tissue culture infective dose,半數(shù)組織感染劑量)。取100 μL稀釋后的病毒與100 μL滅活的各稀釋度的血清混合均勻,置于37℃孵育1 h后加入到96孔板中,37℃孵育2 h。棄上清液,加入添加胰酶的維持培養(yǎng)基(maintenance medium with trypsin,MMT)繼續(xù)培養(yǎng)72 h。每份血清設(shè)3個(gè)重復(fù)。觀察細(xì)胞病變并記錄,按Reed-Muench法計(jì)算血清中和效價(jià)。
1.3.6 臨床樣品檢測(cè)
應(yīng)用建立的ELISA檢測(cè)方法,對(duì)收集具有腹瀉癥狀的262份豬血清樣品進(jìn)行檢測(cè),分別設(shè)置陽(yáng)性和陰性對(duì)照。
2 結(jié)果
2.1 巢式RT-PCR檢測(cè)方法的建立
針對(duì)L1基因共設(shè)計(jì)3對(duì)引物,組合之后進(jìn)行條件摸索(圖1),最終確定的檢測(cè)引物如表1所示。反應(yīng)條件為第1輪循環(huán)數(shù)35,熔解溫度(tm)=55℃,第2輪循環(huán)數(shù)30,tm=57℃。
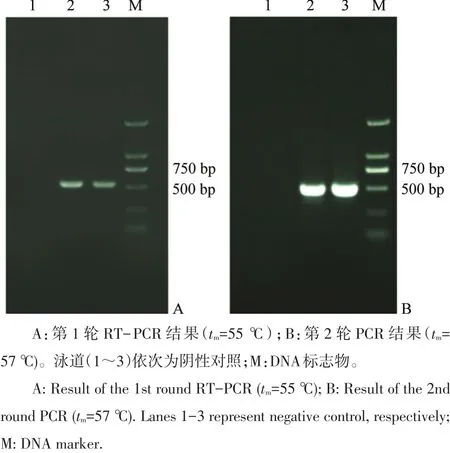
圖1 巢式RT-PCR檢測(cè)條件摸索結(jié)果Fig.1 Electropherogram of the nested RT-PCR product
2.2 巢式RT-PCR檢測(cè)結(jié)果
對(duì)從浙江、江西、山東、河南、黑龍江等5省共58個(gè)豬場(chǎng)采集的224份腹瀉仔豬糞便樣品進(jìn)行巢式RT-PCR檢測(cè)。結(jié)果(附表1,http://www.zjujournals.com/agr/EN/article/showSupportInfo.do?id=10520)顯示:從55個(gè)豬場(chǎng)中共檢出147個(gè)陽(yáng)性樣品,豬場(chǎng)陽(yáng)性率和樣品陽(yáng)性率分別為94.8%和65.6%,其中第1輪從16個(gè)豬場(chǎng)(陽(yáng)性率為27.5%)中檢出陽(yáng)性樣品28個(gè)(陽(yáng)性率為12.5%);從地區(qū)來看,不同地區(qū)采集樣本量不同,樣本量最小的是黑龍江省,其陽(yáng)性率也最高,達(dá)75%,浙江省采集的樣本量最大,從43個(gè)豬場(chǎng)中共采集到159份腹瀉樣品,其中在41個(gè)豬場(chǎng)中檢測(cè)到豬呼腸孤病毒,樣品陽(yáng)性率達(dá)67.9%,且第1輪檢出陽(yáng)性樣品的比例也最高,達(dá)15.7%(表2)。
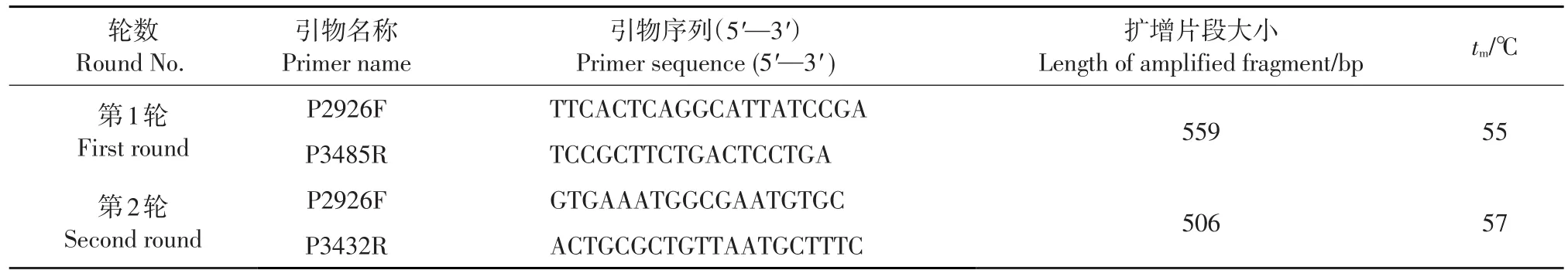
表1 巢式RT-PCR檢測(cè)引物Table 1 Primers of the nested RT-PCR
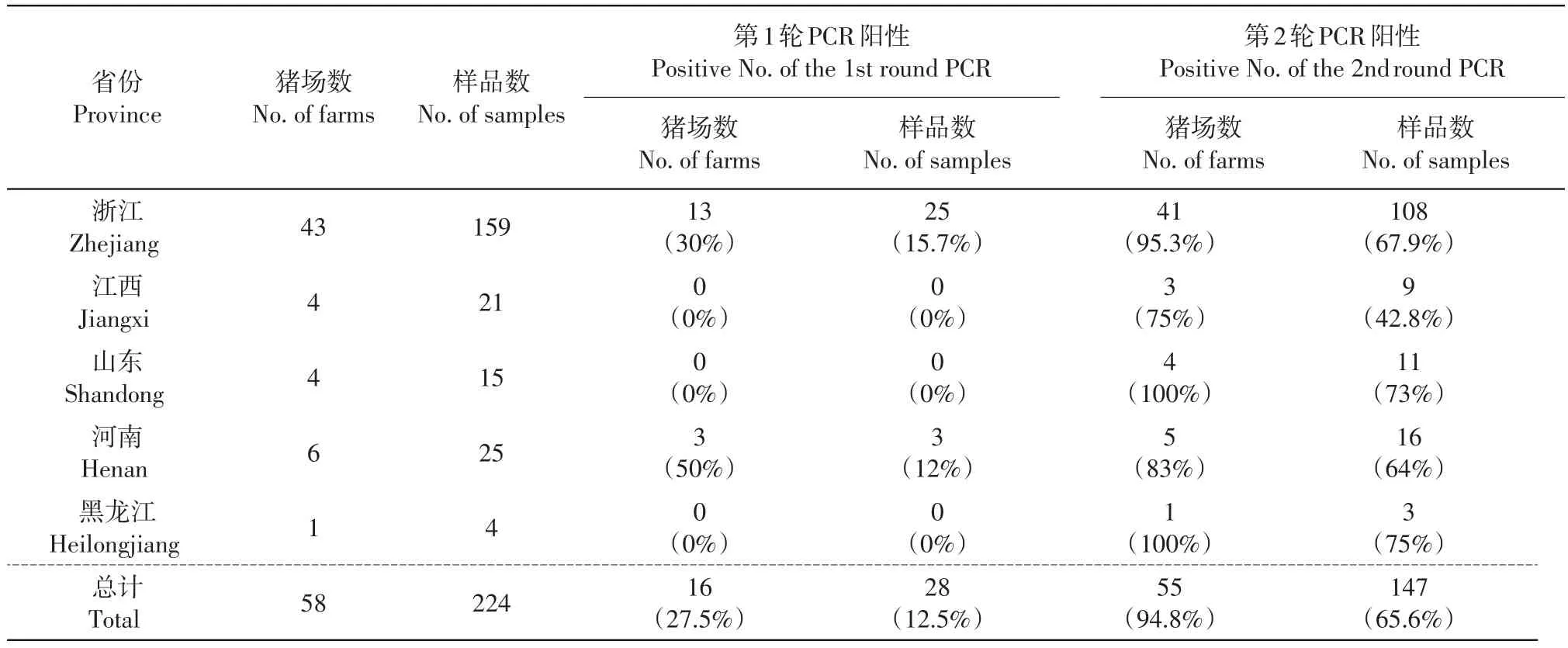
表2 各地區(qū)樣品MRV3檢測(cè)結(jié)果匯總Table 2 Summary of positive rate of the MRV3
2.3 間接ELISA方法的建立
參考本實(shí)驗(yàn)室已經(jīng)建立的方法,從20份具有腹瀉癥狀的豬血清中最終篩選到3份陽(yáng)性血清,結(jié)果如圖2所示。同時(shí)對(duì)初生仔豬血清進(jìn)行篩選,確定了4份陰性血清。將上述3份陽(yáng)性血清混合,用于摸索基于δ1蛋白間接ELISA檢測(cè)方法的最佳條件。最終確定的條件如下:抗原最佳包被量為0.387 ng/孔,血清樣品稀釋度為1∶100,HRP標(biāo)記的羊抗豬抗體按照1∶2 000稀釋。間接ELISA檢測(cè)的臨界(cut-off)值為對(duì)照組(4份陰性血清)在450 nm波長(zhǎng)處的吸光度平均值+3SD,即為0.32。若收集的血清樣本在450 nm波長(zhǎng)處的吸光度值低于0.32則判定為陰性,若等于或者高于0.32則判定為陽(yáng)性。
從收集的血清樣品中隨機(jī)選取37份血清樣品測(cè)定中和效價(jià)。經(jīng)過多次試驗(yàn)認(rèn)定,中和抗體滴度小于等于4時(shí),無法判斷是否由非特異性結(jié)合產(chǎn)生干擾。在37份血清樣本中,中和抗體效價(jià)高于4的有33份(89.2%)。而用建立的間接ELISA方法測(cè)定血清樣品的陽(yáng)性率為86.49%(表3),二者具有較高的一致性,說明建立的間接ELISA方法比較可靠。

圖2 陽(yáng)性血清免疫印跡結(jié)果Fig.2 Result of Western blot analysis of positive serum
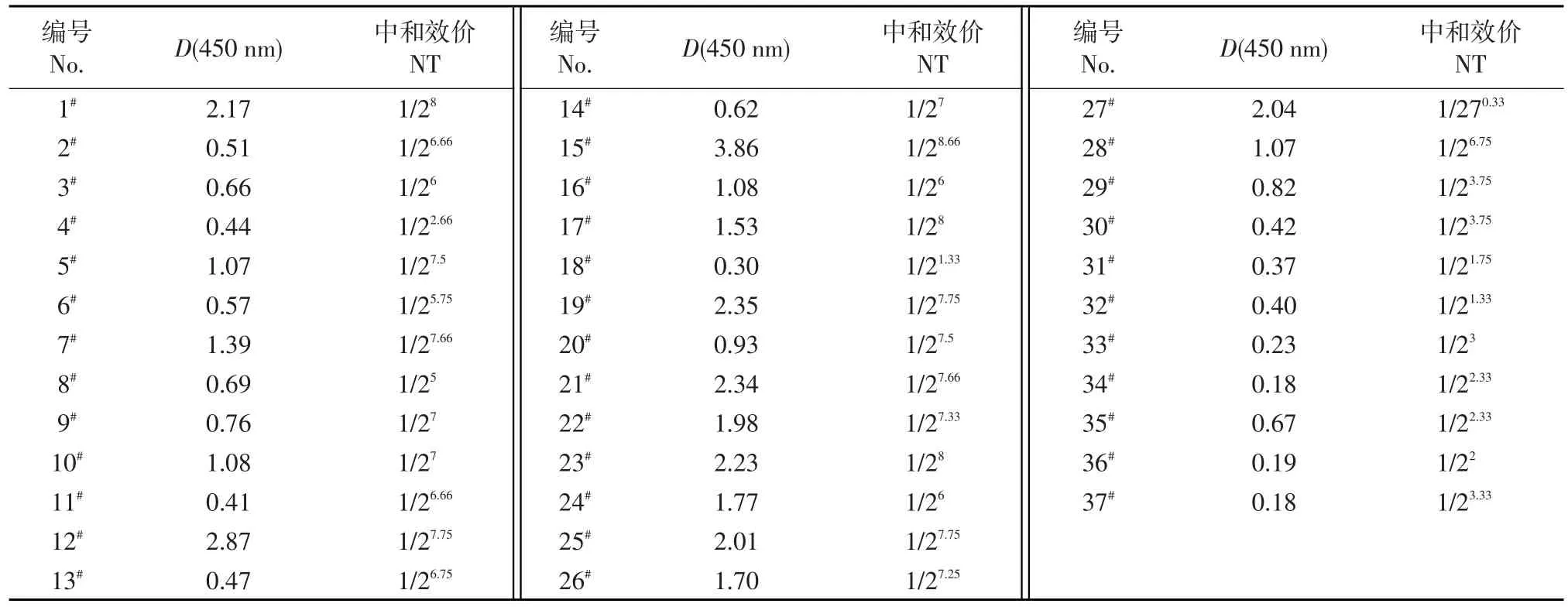
表3 血清樣品在450 nm處的吸光度值和中和抗體效價(jià)對(duì)比結(jié)果Table 3 Comparison of absorbance value at 450 nm and neutralization antibody titer(NT)of serum samples
2.4 臨床樣本間接ELISA檢測(cè)結(jié)果
利用建立的間接ELISA對(duì)收集的262份血清樣品進(jìn)行檢測(cè)。結(jié)果(表4)表明:從整體來看,中國(guó)東部5省不同年齡段的豬中MRV3抗體陽(yáng)性率比較高,達(dá)到84.4%。從不同年齡段來看,陽(yáng)性率最高的是1周齡以內(nèi)仔豬,達(dá)到89.5%;最低的是3周齡仔豬,陽(yáng)性率為75.3%。從地區(qū)來看,江西省陽(yáng)性率最高,達(dá)到95.7%;河南省最低,為72.5%。但是在450 nm處的吸光度平均值各省差異不明顯,最高的是山東省,而最低的是黑龍江省(圖3)。
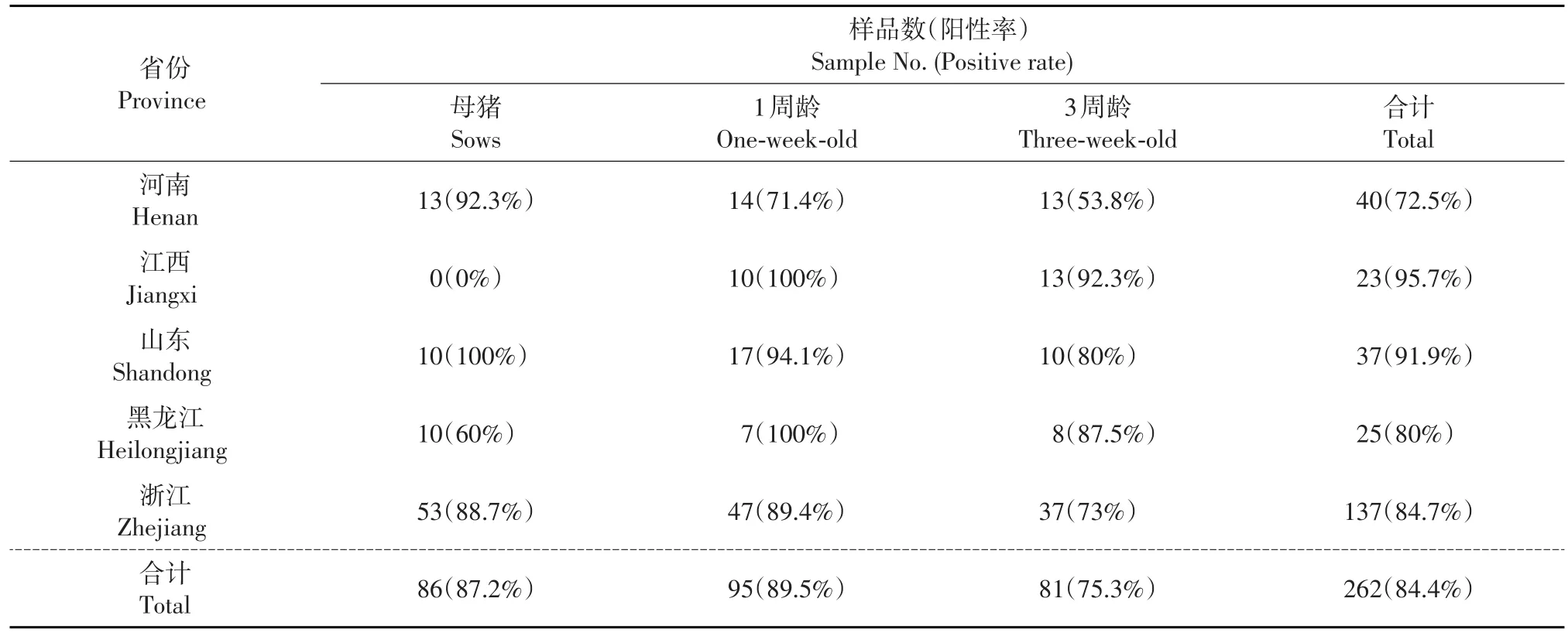
表4 不同地區(qū)不同年齡豬血清樣品中MRV3抗體陽(yáng)性率匯總Table 4 Summary of antibody positive rates of the MRV3 in pig serum samples of different ages from different regions
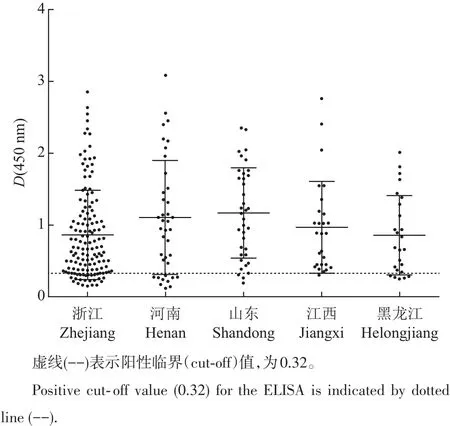
圖3 不同地區(qū)血清抗體水平散點(diǎn)分布圖Fig.3 Distribution of σ1-based ELISA results(anti-MRV3 IgG antibodies)among serum samples collected from different regions
3 討論
仔豬腹瀉一直是困擾全球養(yǎng)豬產(chǎn)業(yè)發(fā)展的一大難題,對(duì)于引起腹瀉的病原也存在爭(zhēng)議。過去認(rèn)為,引起仔豬腹瀉的病毒性病原主要是PEDV、TGEV、RV以及一些腸道病毒(EV),但是,近年來一些使用過疫苗的豬場(chǎng)仍然再次暴發(fā)腹瀉疫情,特別是一些新發(fā)病原例如PDCoV、MRV,引起了各國(guó)學(xué)者的注意。
目前針對(duì)豬呼腸孤病毒的報(bào)道主要集中在基因組分析以及致病性等,對(duì)于豬呼腸孤病毒的流行病學(xué)調(diào)查比較少。談晨[22]用RT-PCR方法檢測(cè)了2012年3月到2013年2月從上海、安徽、廣西、江蘇、浙江、廣東、四川、山東、湖北等省共31個(gè)規(guī)模豬場(chǎng)收集的102份腹瀉病料樣品,結(jié)果顯示,豬場(chǎng)陽(yáng)性率為45.16%,病料樣品陽(yáng)性率為26.47%。KWON等[8]對(duì)2004年1月至2005年12月從韓國(guó)78個(gè)豬場(chǎng)收集的237份2日齡至70日齡腹瀉豬糞便樣品檢測(cè)發(fā)現(xiàn),呼腸孤病毒陽(yáng)性率達(dá)到19%。XIAO等[19]利用熒光定量PCR技術(shù)檢測(cè)了2015年4月至5月從美國(guó)41個(gè)豬場(chǎng)收集的277份腹瀉糞便樣品,結(jié)果顯示,從8個(gè)豬場(chǎng)中(8/41,19.5%)檢測(cè)到了16份陽(yáng)性樣品(16/277,5.7%)。本研究針對(duì)MRV L1基因保守區(qū)設(shè)計(jì)引物建立了巢式RT-PCR檢測(cè)方法。L1基因編碼的λ1蛋白具有RNA依賴的RNA聚合酶活性,在不同血清型呼腸孤病毒之間具有較高的保守性。理論上,針對(duì)L1基因保守區(qū)設(shè)計(jì)引物能夠檢測(cè)到不同血清型的呼腸孤病毒。S1基因是呼腸孤病毒基因組中保守性最低的基因,不同血清型MRV中的S1序列差異比較大,因此,針對(duì)MRV3 S1基因設(shè)計(jì)的檢測(cè)引物可能不能有效地檢測(cè)到其他血清型MRV。因此,針對(duì)L1基因建立的檢測(cè)方法靈敏性更高。這可能是XIAO等[19]的檢測(cè)結(jié)果明顯低于本研究以及其他報(bào)道的原因之一。本研究建立的巢式PCR第2輪引物位于第1輪PCR產(chǎn)物內(nèi)部,而非目的片段包含2套引物結(jié)合位點(diǎn)的可能性極小,因此確保了第2輪PCR產(chǎn)物幾乎或者完全沒有引物配對(duì)特異性不強(qiáng)造成的非特異性擴(kuò)增的污染。因此,相對(duì)于KWON等[8]和談晨[22]針對(duì)L1建立的傳統(tǒng)RTPCR方法,本研究建立的檢測(cè)方法特異性更高。
本研究利用巢氏RT-PCR檢測(cè)了2013—2014年從浙江、江西、山東、河南、黑龍江等省共58個(gè)豬場(chǎng)收集的腹瀉樣品。結(jié)果顯示,從55個(gè)豬場(chǎng)中檢出了147份陽(yáng)性樣品,豬場(chǎng)陽(yáng)性率和樣品陽(yáng)性率分別為94.8%和65.6%。這說明豬呼腸孤病毒廣泛存在于具有腹瀉癥狀的仔豬體內(nèi),可能單獨(dú)或者與其他病原共感染引起腹瀉。
本研究還建立了呼腸孤病毒中和抗體測(cè)定方法以及以δ1蛋白為包被抗原的間接ELISA檢測(cè)方法,首次從抗體水平調(diào)查了我國(guó)豬MRV3流行情況。間接ELISA檢測(cè)結(jié)果顯示,在2015—2016年從山東、河南、江西、黑龍江和浙江等省共25個(gè)豬場(chǎng)收集到的262份腹瀉豬血清樣品中,陽(yáng)性率達(dá)84.4%,并且各地區(qū)陽(yáng)性樣品率以及在450 nm處的吸光度平均值差異不顯著。這從另一角度說明呼腸孤病毒廣泛存在于各個(gè)豬場(chǎng)中。
間接ELISA檢測(cè)樣品主要來自具有腹瀉癥狀的母豬、1周齡內(nèi)仔豬以及3周齡仔豬。從不同年齡段來看,1周齡內(nèi)仔豬樣品陽(yáng)性率最高,母豬次之,3周齡仔豬最低。新生仔豬體內(nèi)抗體主要是通過奶水從母體獲得的母源抗體。因此,我們推測(cè)母豬很有可能是仔豬感染的病毒源。到3周齡斷奶時(shí)母源抗體基本消失,病原入侵時(shí)自身免疫系統(tǒng)將會(huì)產(chǎn)生免疫應(yīng)答,這可能是出現(xiàn)上述現(xiàn)象的原因之一。
盡管血清樣品和腹瀉糞便樣品采集時(shí)間以及來源有所差異,上述數(shù)據(jù)不能完全準(zhǔn)確反映真實(shí)的感染情況,但可以說明豬呼腸孤病毒廣泛存在于中國(guó)東部5省各豬場(chǎng)中,在實(shí)際生產(chǎn)中應(yīng)該做好該病毒的防控。
[1] SONG D,PARK B.Porcine epidemic diarrhoea virus:A comprehensive review of molecular epidemiology,diagnosis,and vaccines.Virus Genes,2012,44(2):167-175.
[2] KIM L,HAYES J,LEWIS P,et al.Molecular characterization and pathogenesis of transmissible gastroenteritis coronavirus(TGEV)and porcine respiratory coronavirus(PRCV)field isolates co-circulating in a swine herd.Archives of Virology,2000,145(6):1133-1147.
[3] SANTOS N,HOSHINO Y.Global distribution of rotavirus serotypes/genotypes and its implication for the development and implementation of an effective rotavirus vaccine.Reviews in Medical Virology,2005,15(1):29-56.
[4] JUNG K,HU H,SAIF L J.Porcine deltacoronavirus infection:Etiology,cellculture forvirusisolation and propagation,molecular epidemiology and pathogenesis.Virus Research,2016,226:50-59.
[5] 曾智勇,郭萬柱,徐志文,等.仔豬腹瀉糞樣中豬呼腸孤病毒的分離鑒定.畜牧獸醫(yī)學(xué)報(bào),2007,38(6):574-580.ZENG Z Y,GUO W Z,XU Z W,et al.Isolation,identification of porcine reovirus from diarrhea feces of pigs.Acta Veterinaria et Zootechnica Sinica,2007,38(6):574-580.(in Chinese with English abstract)
[6] ZHANG C F,LIU L C,WANG P Z,et al.A potentially novel reovirus isolated from swine in northeastern China in 2007.Virus Genes,2011,43(3):342-349.
[7] DAI Y M,ZHOU Q F,ZHANG C W,et al.Complete genome sequence of a porcine orthoreovirus from southern China.Journal of Virology,2012,86(22):12456.
[8] KWON H J,KIM H H,KIM H J,et al.Detection and molecular characterization of porcine type 3 orthoreoviruses circulating in South Korea.Veterinary Microbiology,2012,157(3/4):456-463.
[9] 何小明,姚火春,張洪彪,等.一株豬源1型呼腸孤病毒的分離及鑒定.中國(guó)獸醫(yī)科學(xué),2013,43(1):22-29.HE X M,YAO H C,ZHANG H B,et al.Isolation and identification of a strain of porcine reoviruses serotype 1 in China.Chinese Veterinary Science,2013,43(1):22-29.(in Chinese with English abstract)
[10]NARAYANAPPA A T,SOORYANARAIN H,DEVENTHIRAN J,et al.A novel pathogenic mammalian orthoreovirus from diarrheic pigs and swine blood meal in the United States.MBio,2015,6(3):e00593-15.
[11]LELLI D,BEATO M S,CAVICCHIO L,et al.First identification of mammalian orthoreovirus type 3 in diarrheic pigs in Europe.Virology Journal,2016,13:139.
[12]CHUA K B,VOON K,CRAMERI G,et al.Identification and characterization of a new orthoreovirus from patients with acute respiratory infections.PLoS One,2008,3(11):e3803.
[13]DECARO N,CAMPOLO M,DESARIO C,et al.Virological and molecular characterization of a mammalian orthoreovirus type 3 strain isolated from a dog in Italy.Veterinary Microbiology,2005,109(1/2):19-27.
[14]ANBALAGAN S,SPAANS T,HAUSE B M.Genome sequence of the novel reassortant mammalian orthoreovirus strain MRV00304/13,isolated from a calf with diarrhea from the United States.Genome Announcements,2014,2(3):e00451-14.
[15]LIAN H,LIU Y,ZHANG S F,et al.Novel orthoreovirus from mink,China,2011.Emerging Infectious Diseases,2013,19(12):1985-1988.
[16]LI Z J,SHAO Y H,LIU C G,et al.Isolation and pathogenicity of the mammalian orthoreovirus MPC/04 from masked civet cats.Infection,Genetics and Evolution,2015,36:55-61.
[17]HU T S,QIU W,HE B,et al.Characterization of a novel orthoreovirus isolated from fruit bat,China.BMC Microbiology,2014,14:293.
[18]YANG X L,TAN B,WANG B,et al.Isolation and identification of bat viruses closely related to human,porcine,and mink orthoreoviruses.Journal of General Virology,2015,96(12):3525-3531.
[19]XIAO C T,MENG X J,OPRIESSNIG T.Prevalence of the novel orthoreovirus 3(MRV3)in diarrhea cases in US pigs in 2015//24th International Pig Veterinary Society Congress and 8th European Symposium of Porcine Health Management 2016,Dublin,2016.Berlin Royal Dublin Society,Dublin,Ireland.
[20]LELLI D,MORENO A,STEYER A,et al.Detection and characterization of a novel reassortant mammalian orthoreovirus in bats in Europe.Viruses,2015,7(11):5844-5854.
[21]HUANG Y W,HARRALL K K,DRYMAN B A,et al.Expression of the putative ORF1 capsid protein of Torque teno sus virus 2(TTSuV2)and developmentofWestern blotand ELISA serodiagnostic assays:Correlation between TTSuV2 viral load and IgG antibody level in pigs.Virus Research,2011,158(1/2):79-88.
[22]談晨.初生仔豬腹瀉兩種相關(guān)病毒分子流行病學(xué)調(diào)查及病毒分離和鑒定.南京:南京農(nóng)業(yè)大學(xué),2013:69-77.TAN C.Molecular epidemiological investigation,isolation and identification of two viruses in neonatal piglets with epidemic dirrhea.Nanjing:Nanjing Agricultural University,2013:69-77.(in Chinese with English abstract)
Epidemiological survey and analysis of mammalian orthoreovirus 3 in diarrheic pigs from five provinces of east China.Journal of Zhejiang University(Agric.&Life Sci.),2017,43(5):631-638
QIN Pan1,WANG Jingwei1,WANG Bin1,LEI Ximei1,LI Long2,HUANG Yaowei1*
(1.Institute of Preventive Veterinary Medicine,College of Animal Sciences,Zhejiang University,Hangzhou 310058,China;2.Hangzhou Belta-Biotechnology Co.,Ltd.,Hangzhou 310004,China)
L1 gene;nested reverse transcription-polymerase chain reaction;neutralization antibody;σ 1 protein;indirect enzyme-linked immunosorbent assay;epidemiological survey
S 855.3
A
10.3785/j.issn.1008-9209.2017.04.211
Summary Diarrhea is a common disorder in pigs,and associated dehydration is a leading cause of mortality among piglets,causing substantial economic losses.Studies have found that the etiology of diarrhea is varied,including multiple viral,bacterial,and parasitic pathogens,but viruses such as porcine epidemic diarrhea virus(PEDV),transmissible gastroenteritis virus(TGEV),rotavirus(RV)and other enteroviruses(EV),are the predominant factors in most cases.Repeated outbreaks have also been reported on farms in which piglets had been immunized by vaccines targeting these viruses.Recently,newly emerging viruses,such as mammalian orthoreovirus(MRV),which can cause diarrhea alone or in co-infections with other known pathogens,have been of great concern in China,South Korea,USA and Europe.Fewresearches on MRV,especially its epidemiological survey and analysis have been carried out in China.
國(guó)家重點(diǎn)研發(fā)計(jì)劃(2017YFD0500103)。
黃耀偉(http://orcid.org/0000-0001-9755-8411),E-mail:yhuang@zju.edu.cn
(First author):覃盼(http://orcid.org/0000-0003-0545-3044),E-mail:qinpan@zju.edu.cn
2017-04-21;接受日期(Accepted):2017-05-03
In order to understand the epidemicity of MRV in China,molecular and seroepidemiological survey of MRV type 3 in diarrheic pigs from five provinces of east China was performed in this study.A nested reverse transcription-polymerase chain reaction(RT-PCR)method was designed to target the L1 gene.A total of 224 diarrheic fecal samples collected from 58 pig farms from Zhejiang,Jiangxi,Shandong,Henan and Heilongjiang provinces in 2013—2014 were examined by this method.Our data revealed that porcine orthoreovirus was present in 147 of 224(65.6%)diarrheic porcine fecal samples obtained from 55 of 58(94.8%)pig farms.At the same time,a neutralization test as well as an indirect enzyme-linked immunosorbent assay(ELISA)with a purified σ1 protein as a coat antigen was developed.The neutralization antibody levels and the anti-σ1 IgG antibody levels of 37 serum samples were determined.A good linear relationship between the anti-σ1 antibodies and virus neutralization(VN)antibodies was observed,demonstrating the correlation between them.Then the σ1-based ELISA was applied to the subsequent seroepidemiological survey of MRV3.A total of 262 serum samples collected from the sows,one-week-old piglets as well as three-week-old weaning piglets in 2015—2016 were tested.The result showed these samples had a high positive rate of anti-MRV IgG(84.4%).
Overall,the preliminary results suggest that MRVs are widespread in piglets from five provinces of east China and may collectively contribute to the enteric disease along with other porcine pathogens.
猜你喜歡
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中老年保健(2021年3期)2021-08-22 06:50:04
昆明醫(yī)科大學(xué)學(xué)報(bào)(2021年1期)2021-02-07 01:06:36
現(xiàn)代臨床醫(yī)學(xué)(2021年1期)2021-01-26 00:56:02
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中華養(yǎng)生保健(2020年4期)2020-11-16 01:31:40
中西醫(yī)結(jié)合肝病雜志(2020年2期)2020-10-27 02:18:50
