29種毒力基因在91株食源性單核細(xì)胞增生李斯特氏菌中的分布
2017-12-13 07:39:18,,,,,
中國(guó)人獸共患病學(xué)報(bào) 2017年11期
, , , , ,
·論著·
29種毒力基因在91株食源性單核細(xì)胞增生李斯特氏菌中的分布
李秀娟,趙冬,潘琢,高偉利,徐保紅,秦麗云
目的了解河北省食源性單核細(xì)胞增生李斯特菌(Listeriamonocytogenes,Lm)毒力基因分布特征。方法采用普通PCR方法對(duì)Lm的29個(gè)毒力基因(包括毒力島Ⅰ在內(nèi)的6個(gè)位點(diǎn):prfA、plcA、plcB、hlyA、mpl和actA;內(nèi)化素家族的10個(gè)位點(diǎn):inlA、inlB、inlC、inlD、inlE、inlF、inlG、inlH/C2、inlI和inlJ;其他13個(gè)毒力相關(guān)位點(diǎn):bsh、srtA、iap、sigB、virR、mprF、dltA、dltB、dltC、dltD、srtB、fbpA和hpt)進(jìn)行檢測(cè)。結(jié)果在91株Lm中,有23個(gè)毒力基因的檢出率為100%。26株Lm的29種毒力基因全部檢出,65株Lm存在inlD、inlF、inlG、inlH/C2、inlJ和mpl等6個(gè)毒力基因的不同缺失。缺失最嚴(yán)重的為inlG和inlF,缺失率分別為60.44%和54.95%;其次為mpl基因,缺失率為19.78%。根據(jù)毒力基因缺失情況,91株菌可分為10個(gè)基因型,優(yōu)勢(shì)毒力基因型為具有全部23種毒力基因的Ⅰ型。石家莊地區(qū)的毒力菌株缺失率(41/52)高于河北北部地區(qū)(11/22)。結(jié)論河北省食源性Lm的毒力基因攜帶率高,毒力基因缺失具有多樣性且存在地域差異。
單核細(xì)胞增生李斯特菌;毒力基因;分布特征
單核細(xì)胞增生李斯特氏菌(Listeriamonocytogenes,Lm)是一種重要的食源性人獸共患病原菌,是一種兼性胞內(nèi)寄生菌,可引起人和多種動(dòng)物的胃腸炎、腦膜炎、敗血癥、流產(chǎn)等,新生兒、孕婦、老年人和免疫功能低下者更易感染發(fā)病。Lm可穿越宿主的3個(gè)屏障系統(tǒng)進(jìn)入機(jī)體,引起侵襲性感染,而該侵襲性由多種毒力基因和酶有關(guān),目前研究的毒力基因主要有與黏附和侵襲有關(guān)的因子、與胞內(nèi)感染有關(guān)的因子,以及毒力調(diào)控因子和應(yīng)答調(diào)控因子等。研究報(bào)道顯示,Lm的毒力基因攜帶率較高,但也有不同程度的基因缺失現(xiàn)象,缺失情況各有不同[1-3]。為了能夠進(jìn)一步了解該菌毒力基因的分布特點(diǎn),本研究對(duì)分離自河北省的91株食源性Lm進(jìn)行了29個(gè)毒力基因的檢測(cè),為該菌致病性的評(píng)估和研究提供進(jìn)一步參考。
1 材料和方法
1.1 材料
1.1.1菌株 本研究檢測(cè)的91株食源性Lm,均于2005-2011年間(無(wú)2009年)分離自來(lái)源于河北省不同地區(qū)的生禽肉、生畜肉、水食產(chǎn)品、速凍米面制品及涼拌菜。其中52株Lm分離株由石家莊市疾病預(yù)防控制中心檢出,其余39株Lm由河北省疾病預(yù)防控制中心饋贈(zèng)。所有Lm菌株均經(jīng)API Listeria 鑒定試劑條進(jìn)行鑒定、確認(rèn), 并經(jīng)多重PCR血清型分型[4], 其中 1/2a型(1/2a和3a) 41株、1/2b 型(1/2b、3b和7)30株、1/2c型(1/2c和3c)7株、4b 型(4b、4d和4e)13株。
1.1.2試劑 GoTaq○RColorless Master Mix(美國(guó)Promega公司,批號(hào):00000390447);PCR 引物(大連TAKARA公司合成)。
1.1.3儀器9700型PCR儀(美國(guó)ABI公司);Qiaxcel 毛細(xì)管電泳儀(德國(guó) Qiagen 公司)。
1.2 方法
1.2.1毒力基因擴(kuò)增引物 參考文獻(xiàn)[5-14],篩選出29個(gè)毒力基因,包括毒力島Ⅰ(IPL-1)在內(nèi)的6個(gè)位點(diǎn)prfA、plcA、plcB、hlyA、mpl和actA;毒力島Ⅱ(IPL-2)即內(nèi)化素家族的10個(gè)位點(diǎn)inlA、inlB、inlC、inlD、inlE、inlF、inlG、inlH/C2、inlI和inlJ;其他與毒力相關(guān)的13個(gè)毒力位點(diǎn):bsh、srtA、iap、sigB、virR、mprF、dltA、dltB、dltC、dltD、srtB、fbpA和hpt。引物設(shè)計(jì)采用Primer5.0軟件,根據(jù)NCBI上已經(jīng)公布的序列,選擇保守性較高的區(qū)域作為引物設(shè)計(jì)區(qū)域(表1)。
表1 29種毒力基因檢測(cè)用PCR引物
Tab.1 Primers of 29 virulence genes of Lm
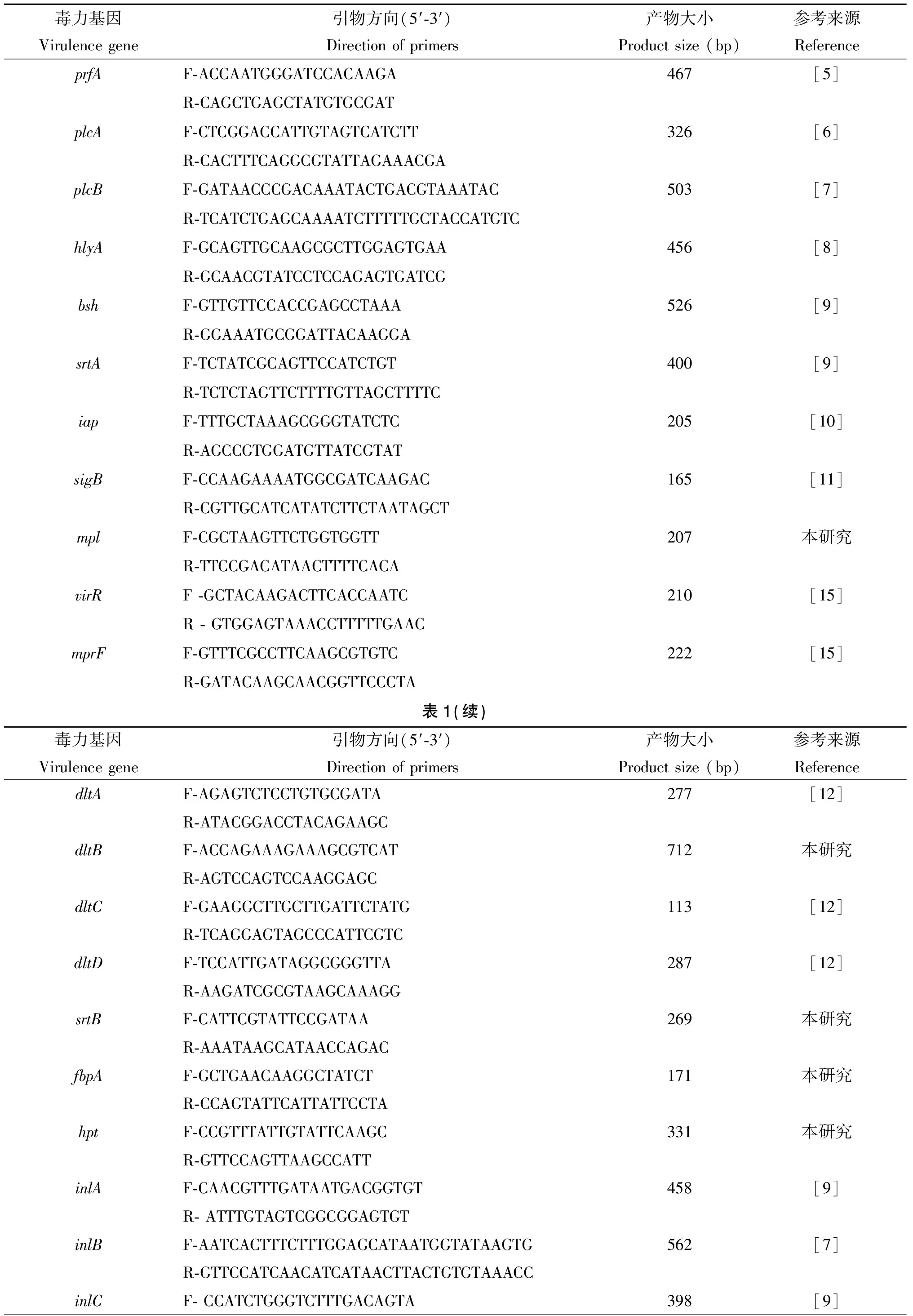
毒力基因Virulencegene引物方向(5′?3′)Directionofprimers產(chǎn)物大小Productsize(bp)參考來(lái)源ReferenceprfAF?ACCAATGGGATCCACAAGA467[5]R?CAGCTGAGCTATGTGCGATplcAF?CTCGGACCATTGTAGTCATCTT326[6]R?CACTTTCAGGCGTATTAGAAACGAplcBF?GATAACCCGACAAATACTGACGTAAATAC503[7]R?TCATCTGAGCAAAATCTTTTTGCTACCATGTChlyAF?GCAGTTGCAAGCGCTTGGAGTGAA456[8]R?GCAACGTATCCTCCAGAGTGATCGbshF?GTTGTTCCACCGAGCCTAAA526[9]R?GGAAATGCGGATTACAAGGAsrtAF?TCTATCGCAGTTCCATCTGT400[9]R?TCTCTAGTTCTTTTGTTAGCTTTTCiapF?TTTGCTAAAGCGGGTATCTC205[10]R?AGCCGTGGATGTTATCGTATsigBF?CCAAGAAAATGGCGATCAAGAC165[11]R?CGTTGCATCATATCTTCTAATAGCTmplF?CGCTAAGTTCTGGTGGTT207本研究R?TTCCGACATAACTTTTCACAvirRF?GCTACAAGACTTCACCAATC210[15]R?GTGGAGTAAACCTTTTTGAACmprFF?GTTTCGCCTTCAAGCGTGTC222[15]R?GATACAAGCAACGGTTCCCTA表1(續(xù))毒力基因Virulencegene引物方向(5′?3′)Directionofprimers產(chǎn)物大小Productsize(bp)參考來(lái)源ReferencedltAF?AGAGTCTCCTGTGCGATA277[12]R?ATACGGACCTACAGAAGCdltBF?ACCAGAAAGAAAGCGTCAT712本研究R?AGTCCAGTCCAAGGAGCdltCF?GAAGGCTTGCTTGATTCTATG113[12]R?TCAGGAGTAGCCCATTCGTCdltDF?TCCATTGATAGGCGGGTTA287[12]R?AAGATCGCGTAAGCAAAGGsrtBF?CATTCGTATTCCGATAA269本研究R?AAATAAGCATAACCAGACfbpAF?GCTGAACAAGGCTATCT171本研究R?CCAGTATTCATTATTCCTAhptF?CCGTTTATTGTATTCAAGC331本研究R?GTTCCAGTTAAGCCATTinlAF?CAACGTTTGATAATGACGGTGT458[9]R?ATTTGTAGTCGGCGGAGTGTinlBF?AATCACTTTCTTTGGAGCATAATGGTATAAGTG562[7]R?GTTCCATCAACATCATAACTTACTGTGTAAACCinlCF?CCATCTGGGTCTTTGACAGTA398[9]

R?CAAATAAGTGACCTTAGTCCTTinlDF?CTTGTGGTCCCTAATAACAT449本研究R?GAACGTATAGCCATCCTTCGinlEF?GGCCTTTCTGGTGAGCTT402本研究R?TGGTTCTTCGTTCTCAGTGinlFF?TGACTTATTTGCAGTTGGGGT1119[9]R?TTGGTTCAGGAATAAGCGCGinlGF?GTGAAGACGGAACTTGGAAA668[13]R?GCTTCTACTATCGGTTGAACAinlH/C2F?ATAGCTACTTTATCAGCATTT437[13]R?ATATCACTTATTTTATTATCATCinlIF?GTTTCCAGACGACAATCTTGCTA635[13]R?AATCGGTACAGTTACTCGCATCAinlJF?AAATCCAGCCCCTGAAAAAG545[9]R?CGCCTGTTTTTGGTAGGGTAactAF?TAGCGTATCACGAGGAGG539[14]R?TTTTGAATTTCATATCATTCACC
1.2.2DNA模板制備 從血平板上挑取過(guò)夜培養(yǎng)的Lm新鮮菌落至400 μL 1×TE(pH8.0)緩沖液中,震蕩混勻,煮沸5 min后冰上冷卻,10 000 r/min離心5 min,取上清于-20 ℃保存?zhèn)溆谩?/p>
1.2.3毒力基因檢測(cè) 29個(gè)位點(diǎn)均采用單重PCR檢測(cè),PCR體系為:2×Gotaq○RColorless Master Mix 12.5 μL,上、下游引物(10 μmol/L)各1 μL,DNA模板1 μL,加水補(bǔ)足體積至25 μL。PCR參數(shù)為:95 ℃ 4 min;94 ℃ 30 s,52 ℃ 30 s,72 ℃ 40 s,35個(gè)循環(huán);72 ℃ 7 min。采用QIAxcel DNA Screening Kit卡夾經(jīng)全自動(dòng)毛細(xì)管電泳儀檢測(cè)PCR產(chǎn)物。
1.2.4PCR產(chǎn)物測(cè)序確認(rèn) 29個(gè)毒力基因的部分陽(yáng)性擴(kuò)增產(chǎn)物送大連寶生物公司進(jìn)行雙向測(cè)序,并對(duì)測(cè)序結(jié)果進(jìn)行序列比對(duì),確認(rèn)擴(kuò)增產(chǎn)物為目的基因。
1.2.5統(tǒng)計(jì)分析 利用Bionumerics Version 7.6軟件,采用Categorical UPGMA方法進(jìn)行缺失毒力基因聚類分析,采用SPSS 21.0軟件對(duì)不同地區(qū)分離的菌株毒力基因缺失差異情況進(jìn)行χ2檢驗(yàn),以Plt;0.05為差異有統(tǒng)計(jì)學(xué)意義。
2 結(jié) 果
2.1毒力基因檢測(cè)結(jié)果 在91株Lm中,有23個(gè)毒力基因的檢出率均為100%。其中26株Lm的29種毒力基因全部檢出,65株Lm分別存在inlD、inlF、inlG、inlH/C2、inlJ和mpl等6個(gè)毒力基因的不同缺失。49株Lm菌株出現(xiàn)2種及2種以上毒力基因的缺失。
表2 91株Lm毒力基因檢測(cè)結(jié)果
Tab.2 Results of 29 virulence genes in 91 foodborne Lm

毒力基因Virulencegenes陽(yáng)性菌株數(shù)(株)No.ofpositivestrains檢出率(%)DetectionratesinlD8492.31inlG3639.56inlF4145.05inlH/C28997.80inlJ8492.31mpl7380.22prfA、plcA、plcB、hlyA、actA、inlA、inlB、inlC、inlE、inlI、bsh、srtA、iap、sigB、virR、mprF、dltA、dltB、dltC、dltD、srtB、fbpA、hpt91100.00
2.2毒力基因分型分析 根據(jù)不同毒力基因缺失情況進(jìn)行聚類分析(圖1),91株Lm可分為10個(gè)毒力基因型別,其中型別最多的Ⅰ型,由具有全部29個(gè)毒力基因的26株菌株組成,占28.57%,其次為同時(shí)缺失InlF和InlG毒力基因的Ⅵ型(23株)以及不僅缺失InlF和InlG毒力基因,還缺失mpl毒力基因的Ⅸ型(17株),分別占25.27%和18.68%。
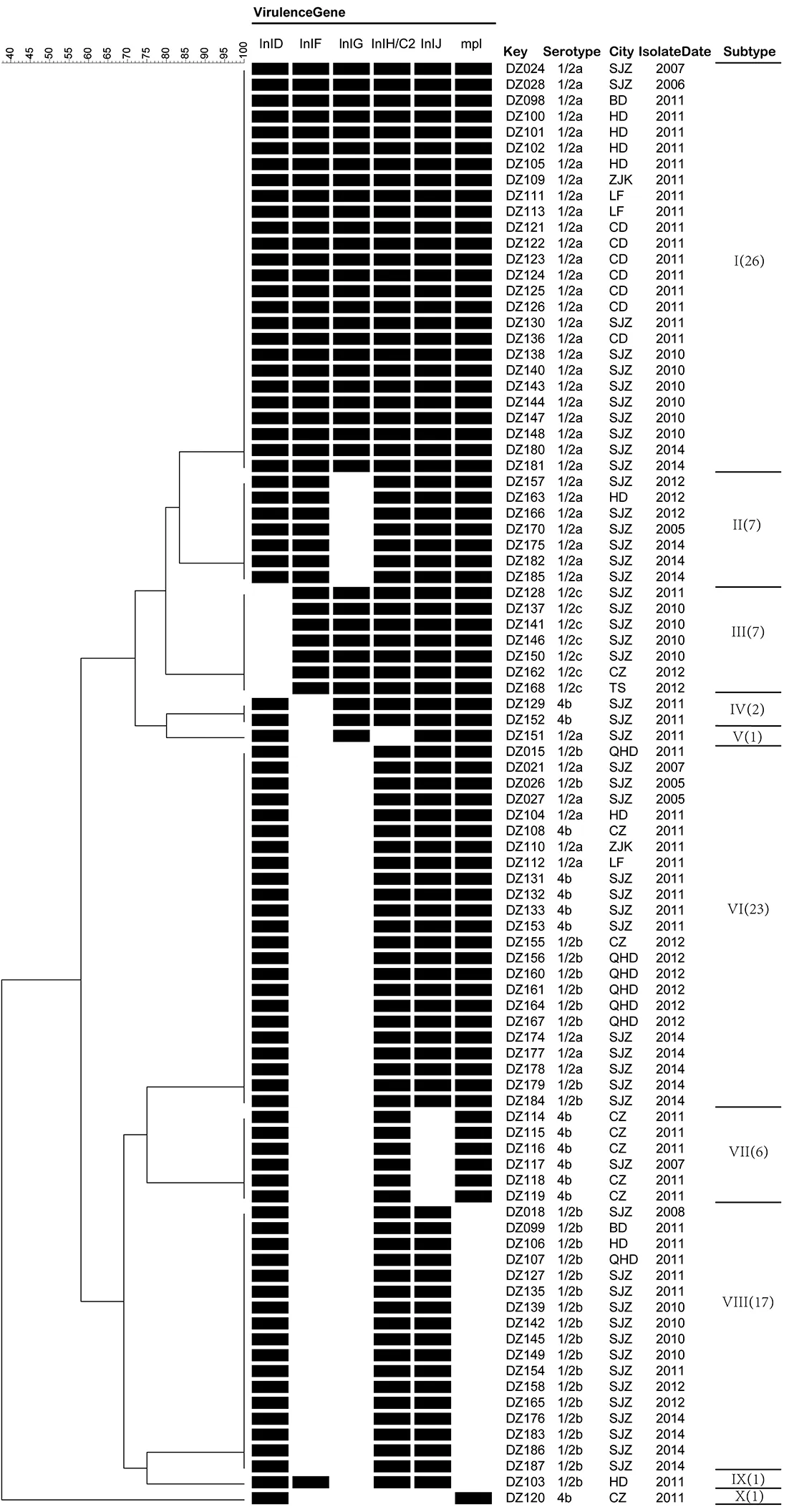
圖1 缺失毒力基因聚類分析圖Fig.1 Clustering analysis of deletion virulence genes
2.3毒力基因缺失與分子血清型關(guān)系 29種毒力基因全部檢出的26株菌株均為1/2a血清型。在出現(xiàn)缺失的6個(gè)毒力基因中,inlD缺失菌株全部為1/2c型(7/7);inlJ缺失菌株全部為4b型(7/13);mpl缺失菌株全部為1/2b型(18/30)。1/2a型菌株主要缺失inG和/inlF基因;1/2b型菌株主要為inG、inlF和mpl基因的缺失;1/2c型菌株主要缺失inlD;4b型菌株主要為inlF、inlG和inlJ3種毒力基因的缺失。在10個(gè)毒力基因型中,除了Ⅵ型中包含有1/2a、1/2b和4b血清型外,其余各型均只有1種血清型(圖1、表3)。
2.465株毒力基因缺失菌株分離地區(qū)分析 為方便計(jì)算,本研究將65株毒力基因缺失菌株按地區(qū)來(lái)源不同分為3大地區(qū),其中石家莊地區(qū)41株,河北南部地區(qū)(邯鄲、衡水、滄州)13株,河北北部地區(qū)(保定、承德、廊坊、秦皇島、唐山、張家口)11株,不同地區(qū)分離菌株毒力基因缺失差異有統(tǒng)計(jì)差異(χ2=6.125,P=0.043),石家莊地區(qū)分離的Lm菌株毒力基因缺失率(78.85%)高于河北北部地區(qū)(50%)(χ2=6.158,P=0.013)。河北北部地區(qū)和石家莊地區(qū)的主要分離株為1/2b型, 河北南部地區(qū)主要分離株為4b型。在河北北部和河北南部地區(qū)Lm分離株中,毒力基因型主要為Ⅶ(8)型,分別占8株和5株,主要缺失毒力基因?yàn)閕nlF和inlG。石家莊地區(qū)由于菌株最多,表現(xiàn)出更多的毒力基因型,主要毒力基因型為Ⅷ(14)、Ⅵ(12)、Ⅱ(6)和Ⅲ(5)型,缺失主要毒力基因inlF、inlG和mpl(表4)。
表3 65株毒力基因缺失菌株血清型
Tab.3 Serotype of 65 Lm strains with virulence genes deletion

缺失毒力基因Deletionvirulencegenes菌株數(shù)(株)No.ofstrains分子血清型(菌株數(shù))Serotype(No.ofstrains)inlD71/2c(7)inlF501/2a(9);1/2b(28);4b(13)inlG551/2a(14);1/2b(30);4b(11)inlH/C224b(1);1/2a(1)inlJ74b(7)mpl181/2b(18)
表4 不同地區(qū)分離Lm菌株的分子血清型與毒力基因型別分布
Tab.4 Regional distribution of serotype and subtype of virulence genes in 91 Lm strains

地區(qū)District菌株總數(shù)No.ofstrains毒力基因缺失菌株數(shù)No.ofstrainswithvirulencegenesdeletion缺失毒力基因菌株分子血清類型Serotypeofstrainswithvirulencegenedeletion1/2a1/2b4b1/2c河北北部NorthernHebei2211Ⅵ(2)Ⅵ(6)Ⅷ(2)lt;FounderNodename=""value=""/gt;Ⅲ(1)河北南部SouthernHebei1713Ⅱ(1)Ⅵ(1)Ⅸ(1)Ⅷ(2)Ⅵ(1)Ⅶ(5)Ⅹ(1)Ⅲ(1)石家莊Shijiazhuang5241Ⅱ(5)Ⅴ(1)Ⅵ(5)Ⅱ(1)Ⅵ(4)Ⅷ(14)Ⅳ(2)Ⅵ(3)Ⅶ(1)Ⅲ(5)合計(jì)Total9165Ⅱ(6)Ⅴ(1)Ⅵ(8)Ⅱ(1)Ⅸ(1)Ⅵ(10)Ⅷ(18)Ⅳ(2)Ⅵ(4)Ⅶ(6)Ⅹ(1)Ⅲ(7)
3 討 論
Lm一般經(jīng)胃腸道感染,侵入腸上皮細(xì)胞后被單核巨噬細(xì)胞吞噬,并隨其擴(kuò)散到局部淋巴結(jié),最后到達(dá)內(nèi)臟器官,引起全身性感染。整個(gè)感染過(guò)程由一系列的步驟組成,包括內(nèi)化、逃離吞噬泡、Lm在宿主細(xì)胞內(nèi)的極向運(yùn)動(dòng)與細(xì)胞間傳播,需要許多毒力蛋白因子和酶參與其中。Lm的致病性與其毒力基因密切相關(guān),缺失毒力基因?qū)?dǎo)致其致病性的消失或下降。通過(guò)檢測(cè)毒力基因,可掌握這些基因在Lm中的分布,從而為評(píng)估Lm分離株的致病性強(qiáng)弱,進(jìn)行相關(guān)監(jiān)測(cè)提供實(shí)驗(yàn)室支持。
本研究對(duì)91株河北省食源性Lm進(jìn)行了29個(gè)毒力基因的檢測(cè),包括毒力島Ⅰ(Listeriamonocytogenespathogenicity islands,LIPI-1)在內(nèi)的6個(gè)基因、內(nèi)化素家族的10個(gè)基因位點(diǎn)inlA、inlB、inlC、inlD、inlE、inlF、inlG、inlH/C2、inlI和inlJ,以及其他目前研究較多的13個(gè)毒力相關(guān)位點(diǎn):bsh、srtA、iap、sigB、virR、mprF、dltA、dltB、dltC、dltD、srtB、fbpA和hpt。研究結(jié)果顯示河北省食源性Lm具有很高的毒力基因攜帶率,29個(gè)毒力基因中有23個(gè)位點(diǎn)檢出率為100%,僅6個(gè)毒力基因出現(xiàn)不同程度的缺失,具有較高的潛在致病性,該結(jié)果與目前報(bào)道一致[1,3,10,16],因此我國(guó)應(yīng)加強(qiáng)食品中該病菌的監(jiān)測(cè),臨床治療中應(yīng)增強(qiáng)對(duì)李斯特菌病的重視和培訓(xùn),從而更好的發(fā)現(xiàn)和防控李斯特菌病。
在91株Lm中,有65株Lm有1-4種毒力基因的缺失,毒力基因缺失菌株比例較高,總?cè)笔蕿?1.43%。根據(jù)缺失毒力基因的聚類分析,91株菌株分為10個(gè)毒力基因型,具有較高的多態(tài)性。除毒力基因Ⅵ型含有1/2a、1/2b和4b血清型以外,其余9個(gè)型別都僅有一種血清型,提示了毒力基因型別與血清型具有較高的相關(guān)性,其內(nèi)在關(guān)系還需進(jìn)一步研究。我國(guó)食品中Lm分離株主要為1/2a血清型[17],而引起食源性疾病最多的血清型也是1/2a血清型[18],本次檢測(cè)結(jié)果也顯示63.4%的1/2a菌株具有全部29個(gè)毒力基因,菌株的致病力較強(qiáng),較易引發(fā)食源性疾病。
LIPL-1毒力島與Lm的致病性密切相關(guān),是細(xì)菌在細(xì)胞內(nèi)的生存必不可少的,具有高度保守性[19]。本次檢測(cè)的91株Lm中,IPL-1中的6個(gè)毒力基因,除了mpl基因以外,均具有較高的穩(wěn)定性。mpl基因編碼鋅依賴金屬蛋白酶(Zinc metalloprotease),參與Lm在細(xì)胞間的直接傳遞,可避免自身被體液免疫系統(tǒng)清除,此外,該酶還介導(dǎo)毒力因子卵磷脂磷脂酶C(PC-PLC)(由plcB基因編碼)的前體在吞噬體中的活性,是LIPL-1中的一個(gè)重要的毒力因子。本次研究顯示在60%的1/2b血清類型菌株(18/30)中出現(xiàn)缺失或變異,提示在該血清類型菌株具有較大的毒力缺陷,致病性較弱。
內(nèi)化素是Lm毒力相關(guān)基因編碼的一個(gè)蛋白質(zhì)家族,介導(dǎo)細(xì)菌侵入無(wú)吞噬能力的上皮細(xì)胞,其N端都含有富含亮氨酸的串聯(lián)重復(fù)區(qū)域,該區(qū)域被認(rèn)為與蛋白質(zhì)-蛋白質(zhì)相互作用及多種功能相關(guān),參與黏附、配體-受體相互作用以及信號(hào)傳導(dǎo)等過(guò)程。InlA、InlB以及InlC是研究較多的內(nèi)化素蛋白,分別在黏附、侵襲以及胞內(nèi)生存和擴(kuò)散方面具有重要作用[20-23]。內(nèi)化素基因容易發(fā)生堿基突變,具有多態(tài)性,還容易發(fā)生水平基因轉(zhuǎn)移和重組[24]。本研究檢測(cè)的10個(gè)內(nèi)化素家族成員中有5個(gè)基因發(fā)生了變異和缺失,inlG和inlF2個(gè)基因的缺失率均高于50%,inlD、inlH/C2和inlJ等3個(gè)基因也發(fā)生了不同程度的缺失。本研究還發(fā)現(xiàn),除了1/2a血清型外,其余3種血清類型,每一種血清類型均出現(xiàn)了一種特定內(nèi)化素基因的缺失,如所有1/2b血清型菌株均缺失inlG基因,所有4b菌株均缺失inlF基因,所有1/2c菌株均缺失inlD基因;另外還有一些基因的缺失僅出現(xiàn)在某一種特定血清型的菌株中,如inlD缺失僅見(jiàn)于1/2c型菌株;inlJ基因缺失也是只在部分4b型菌株中出現(xiàn)。這兩種不同形式的內(nèi)化素基因缺失狀況,顯示出內(nèi)化素基因的缺失與血清型具有一定的相關(guān)性,可能是該血清型中一些菌株的毒力基因發(fā)生了變異,還需要進(jìn)一步研究。
在本研究檢測(cè)的13種毒力相關(guān)基因中,bsh和sigB與細(xì)菌耐受環(huán)境的壓力應(yīng)答有關(guān),srtB與逃避巨噬細(xì)胞吞噬有關(guān),srtA、fbpA與細(xì)菌的侵襲、粘附相關(guān),iap、mprF與抵抗宿主防御有關(guān),hpt與細(xì)胞內(nèi)增值有關(guān),dltA、dltB、dltC、dltD與細(xì)胞感染相關(guān),virR具有調(diào)控功能,是一種應(yīng)答調(diào)控因子,調(diào)控包括mprF、dltA、dltB等在內(nèi)的12 個(gè)毒力基因。這13種毒力基因是細(xì)菌耐受環(huán)境壓力、細(xì)胞內(nèi)生存、逃逸以及細(xì)胞感染有關(guān)的基礎(chǔ)基因,具有一定的保守型。本研究結(jié)果也顯示該13種毒力相關(guān)基因在91株菌株中均存在,進(jìn)一步證明該13種基因在Lm菌株中的高度保守性,其缺失將可能會(huì)明顯影響菌株的生存。
本研究通過(guò)對(duì)91株食源性Lm中29個(gè)毒力基因的檢測(cè),證明了一些毒力基因?qū)τ贚m生存感染是必須的,具有高度的保守性;另有一些毒力基因如內(nèi)化素家族成員基因和mpl基因則具有一定的多態(tài)性,容易發(fā)生變異和缺失。曾有文獻(xiàn)報(bào)道hlyA、inlA和plcB3個(gè)毒力基因的檢出情況與Lm的MLST分型聚類有較一致的關(guān)系,可為分析Lm毒力強(qiáng)弱提供參考意義[25],本研究發(fā)現(xiàn)毒力基因缺失菌株具有一定的地域差異,毒力基因缺失變異與菌株的血清類型也具有一定聯(lián)系,因此進(jìn)一步研究毒力基因缺失變異的地域特點(diǎn)以及與血清類型的關(guān)系對(duì)于制定防控策略以及分析Lm的毒力強(qiáng)弱同樣具有重要的參考意義。
[1] He CY, Wang Y, Wang TS, et al. Distribution of virulence-related genes of selected Listeria monocytogenes strains isolated from food samples in China[J]. Dis Surveill, 2013, 28(5): 384-388. (in Chinese)
賀春月,王艷,王天姝,等.中國(guó)部分食品來(lái)源單增李斯特菌中致病相關(guān)基因的分布研究[J]. 疾病監(jiān)測(cè), 2013, 28(5): 384-388.
[2] Yang Y, Fu P, Guo YC, et al. Virulent gene prevalence of foodborneListeriamonocytogenesin China in 2005[J]. Chin J Prev Med, 2010, 44(12): 1097-1101. (in Chinese)
楊洋,付萍,郭云昌,等. 2005年中國(guó)食源性單核細(xì)胞增生李斯特菌毒力基因分布[J].中華預(yù)防醫(yī)學(xué)雜志, 2010, 44(12): 1097-1101.
[3] Zhang LR, Wang Y, Zhang Y, et al. Biological and molecular epidemiological characteristics ofListeriamonocytogenesisolated in Tongzhou, Beijing[J]. Dis Surveill, 2007, 27(7): 562-564. (in Chinese)
張?zhí)m榮,王艷,張揚(yáng),等. 2007-2011年北京市通州區(qū)分離單增李斯特菌的生物學(xué)及分子流行病學(xué)特征[J].疾病監(jiān)測(cè), 2007, 27(7): 562-564.
[4] Doumith M, Buchrieser C, Glaser P, et al. Differentiation of the majorListeriamonocytogenesserovars by multiplex PCR[J]. J Clin Microbiol, 2004, 42(8): 3819-3822. DOI: 10.1128/JCM.42.8.3819-3822.2004
[5] Bubert A, Sokolovic Z, Chun SK, et al. Differential expression ofListeriamonocytogenesvirulence genes in mammalian host cells[J]. Mol Gen Genet, 1999, 261(2): 323-336. DOI: 10.1007/PL00008633
[6] Lomonaco S, Patti R, Knabel SJ,et al. Detection of virulence-associated genes and epidemic clone markers inListeriamonocytogenesisolates from PDO Gorgonzola cheese[J]. Int J Food Microbiol, 2012, 160(1): 76-79. DOI: 10.1016/j.ijfoodmicro.2012.09.011
[7] Volokhov D, Rasooly A, Chumakov K, et al. Identification ofListeriaspecies by microarray-based assay[J]. J Clin Microbiol, 2002, 40(12): 4720-4728. DOI: 10.1128/JCM.40.12.4720-4728.2002
[8] Paziak-Domańska B, Bogusawska E, Wieckowska-Szakiel M, et al. Evaluation of the API test, phosphatidylinositol- specific phospholipase C activity and PCR method in identification ofListeriamonocytogenesin meat foods[J]. FEMS Microbiol Lett, 1999, 171(2): 209-214. DOI: 10.1111/j.1574-6968.1999.tb13434.x
[9] Lomonaco S, Chen Y, Knabel SJ. Analysis of additional virulence genes and virulence gene regions inListeriamonocytogenesconfirms the epidemiologic relevance of multi-virulence-locus sequence typing[J]. J Food Prot, 2008, 71(12): 2559-2566. DOI: 10.4315/0362-028X-71.12.2559
[10] Gong ZL, Zhu RF, Ye CY, et al. Detection of virulence genes in 118 strains of ChinaListeriamonocytogenes[J]. Dis Surveill, 2007, 22(5): 299-301. (in Chinese)
宮照龍,祝仁發(fā),葉長(zhǎng)蕓,等. 118株單核細(xì)胞增生李斯特菌的毒力基因檢測(cè)[J]. 疾病監(jiān)測(cè),2007, 22(5): 299-301.
[11] Kalliopi R, Marios M, Valentina A, et al. Expression of virulence genes ofListeriamonocytogenesin food[J]. J Food Safety, 2012, 32 (2): 161-168. DOI: 10.1111/j.1745-4565.2011.00363.x
[12] Li XH. Distribution and pathogenicity to virulence genes of foodborneListeriamonocytogenes[D]. Shijiazhuang: Agricultural University of Hebei, 2012: 1-45. (in Chinese)
李欣華.食源性單核增生性李斯特氏菌毒力基因分布及致病力的研究[D].石家莊:河北農(nóng)業(yè)大學(xué), 2012:1-45.
[13] Chen J, Luo X, Jiang L, et al. Molecular characteristics and virulence potential ofListeriamonocytogenesisolates from Chinese food systems[J]. Food Microbiol, 2009, 26(1): 103-111. DOI: 10.1016/j.fm.2008.08.003
[14] Wiedmann M, Bruce JL, Keating C, et al. Ribotypes and virulence gene polymorphisms suggest three distinctListeriamonocytogeneslineages with differences in pathogenic potential[J]. Infect Immun, 1997, 65 (7): 2707-2716.
[15] Li XH, Han J, Yu HW, et al. Distribution of virR and mprF genes inListeriamonocytogenesfrom different food sources[J]. J Agr Univ Hebei, 2012, 35(3): 86-89. (in Chinese)
李欣華,韓軍,于宏偉, 等.單核細(xì)胞增生性李斯特氏菌virR和mprF基因分布[J].河北農(nóng)業(yè)大學(xué)學(xué)報(bào), 2012, 35(3):86-89.
[16] Chen JS, Jiang LL, lü YH, et al. Molecular epidemiology and pathogenic potential ofListeriamonocytogenesin aquatic products[J]. J Chin Institute Food Sci Technol, 2013, 13(9): 182-189. (in Chinese)
陳建舜,江玲麗,呂永輝,等. 水產(chǎn)品中單增李斯特菌的分子流行病學(xué)特征與致病力研究[J]. 中國(guó)食品學(xué)報(bào), 2013, 13(9):182-189.
[17] Shen J, Rump L, Zhang Y, et al. Molecular subtyping and virulence gene analysis ofListeriamonocytogenesisolates from food[J]. Food Microbiol, 2013, 35(1): 58-64. DOI: 10.1016/j.fm.2013.02.014
[18] Kathariou S.Listeriamonocytogenesvirulence and pathogenicity, a food safety perspective[J]. J Food Prot, 2002, 65(11): 1811-1829. DOI: 10.4315/0362-028X-65.11.1811
[19] Vázquez-Boland JA, Kuhn M, Berche P, et al.Listeriapathogenesisand molecular virulence determinants[J]. Clin Microbiol Rev, 2001, 14(3): 584-640. DOI: 10.1128/CMR.14.3.584-640.2001
[20] Mengaud J, Ohayon H, Gounon P, et al. E-cadherin is the receptor for internalin, a surface protein required for entry of L. monocytogenes into epithelial cells[J]. Cell, 1996, 84(6):923-932. DOI: 10.1016/S0092-8674(00)81070-3
[21] Braun L, Dramsi S, Dehoux P, et al. InlB: an invasion protein ofListeriamonocytogeneswith a novel type of surface association[J]. Mol Microbiol, 1997, 25(2): 285-294. DOI: 10.1046/j.1365-2958.1997.4621825.x
[22] Dussurget O. New insights into determinants ofListeriamonocytogenesvirulence[J]. Int Rev Cell Mol Biol, 2008, 270: 1-38. DOI: 10.1016/S1937-6448(08)01401-9
[23] Ren JJ, Yang MW, Chen YF, et al. Biological characters and pathogenicity ofListeriamonocytogenesΔinlA/ΔinlB double gene deletion strains[J]. Chin J Zoonoses, 2016, 32 (6): 529-534. (in Chinese)
任靜靜,楊銘偉,陳云飛,等.單增李斯特菌inl A/ inl B雙基因缺失株生長(zhǎng)特性觀察及對(duì)小鼠致病性研究[J].中國(guó)人獸共患病學(xué)報(bào), 2016, 32 (6): 529-534.
[24] Tsai YH, Orsi RH, Nightingale KK, et al.Listeriamonocytogenesinternalins are highly diverse and evolved by recombination and positive selection[J]. Infect Genet Evol, 2006, 6(5): 378-389. DOI: 10.1016/j.meegid.2006.01.004
[25] Liu EL, Yuan MY, Lü YZ, et al. Development of triplex TaqMan MGB-probe real-time PCR assay for detection ofListeriamonocytogenesand the distribution of virulence gene in isolates[J]. Chin J Zoonoses, 2016,32 (5): 451-456. (in Chinese)
劉二龍,袁慕云,呂英姿,等.單增李斯特菌三重實(shí)時(shí)熒光PCR檢測(cè)的建立及其毒力基因在分離菌株中分布[J].中國(guó)人獸共患病學(xué)報(bào), 2016, 32 (5): 451-456.
Distributioncharacterizationof29virulencegenesin91foodborneListeriamonocytogenesstrains
LI Xiu-juan, ZHAO Dong, PAN Zhuo, GAO Wei-li, XU Bao-hong, QIN Li-yun
(DepartmentofMicrobiology,ShijiazhuangCenterforDiseaseControlandPrevention,Shijiazhuang050011,China)
In order to study the distribution of virulence genes ofListeriamonocytogenes(Lm) in Hebei Province, 29 virulence genes ofLm, includingListeriamonocytogenespathogenicity islands Ⅰ(LIPI-Ⅰ:prfA,plcA,plcB,hlyA,mplandactA), 10 internalins genes (inlA,inlB,inlC,inlD,inlE,inlF,inlG,inlH/C2,inlIandinlJ) and the other 13 virulence-associated genes (bsh,srtA,iap,sigB,virR,mprF,dltA,dltB,dltC,dltD,srtB,fbpAandhpt) were detected by PCR. Results showed that in the 91 Lm strains, the detection rate of 23 virulence genes were 100%. The 29 virulence genes of 26 Lm strains were all detected, and 65Lmstrains had different deletion of 6 virulence genesinlD,inlF,inlG,inlH/C2,inlJandmpl. The deletion rate ofinlGandinlFwere 60.44% and 54.95%, respectively, following by mpl gene, with a deletion rate of 19.78%. According to the absence of virulence genes, 91 strains could be divided into 10 subtypes, and the dominant virulence subtypes was type Ⅰwith all 23 virulence genes.The deletion rate of virulent genes in Shijiazhuang was higher than that in northern Hebei. It is suggested that the rate of virulence gene of food-borneLmin Hebei Province is high, and the virulence gene deletion patterns has diversity and regional differences.
Listeriamonocytogenes; virulence genes; distribution characteristics
10.3969/j.issn.1002-2694.2017.11.005
河北省醫(yī)學(xué)科學(xué)研究重點(diǎn)課題(No. ZL20140244)
石家莊市疾病預(yù)防控制中心,石家莊 050011
Email:wsws1120@126.com
R378.99
A
1002-2694(2017)11-0972-07
Supported by the Hebei Medical Scientific Research Fund Project(No. ZL20140244)
2017-02-20編輯張智芳
猜你喜歡
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
幼兒園(2019年7期)2019-09-05 17:49:18
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
海峽科技與產(chǎn)業(yè)(2016年3期)2016-05-17 04:32:12
小小說(shuō)月刊(2013年6期)2013-05-14 14:55:19
