菊芋渣選擇性吸附天然菊粉酶催化制備短鏈低聚果糖
2017-12-15 08:39:13賴晨歡
生物質(zhì)化學(xué)工程 2017年6期
關(guān)鍵詞:質(zhì)量
李 鑫, 周 瑾, 賴晨歡, 勇 強(qiáng)
(1. 南京林業(yè)大學(xué) 江蘇省林業(yè)資源高效加工利用協(xié)同創(chuàng)新中心, 江蘇 南京 210037;2. 南京林業(yè)大學(xué) 化學(xué)工程學(xué)院, 江蘇 南京 210037)
·研究報(bào)告——生物質(zhì)化學(xué)品·
菊芋渣選擇性吸附天然菊粉酶催化制備短鏈低聚果糖
李 鑫1,2, 周 瑾2, 賴晨歡1,2, 勇 強(qiáng)1,2
(1. 南京林業(yè)大學(xué) 江蘇省林業(yè)資源高效加工利用協(xié)同創(chuàng)新中心, 江蘇 南京 210037;2. 南京林業(yè)大學(xué) 化學(xué)工程學(xué)院, 江蘇 南京 210037)
以菊粉加工廢棄物(菊芋渣)選擇性吸附黑曲霉產(chǎn)的天然菊粉酶提高菊粉酶活力(I)與轉(zhuǎn)化酶活力(S)的比值(即I/S比值),并將此純化后的菊粉酶應(yīng)用于后續(xù)的短鏈低聚果糖(FOSs)的生產(chǎn),考察了純化后菊粉酶對(duì)FOSs產(chǎn)率的影響。結(jié)果顯示:選擇的較佳吸附條件為:菊芋渣質(zhì)量濃度為100 g/L,吸附pH值為5.0,吸附溫度為4 ℃。經(jīng)菊芋渣的選擇性吸附,天然菊粉酶酶系中轉(zhuǎn)化酶活力降低80.84%,菊粉酶活力保留67.92%,I/S比值由原來(lái)的1.13上升至4.02。經(jīng)菊芋渣選擇性吸附后的天然菊粉酶用于制備短鏈低聚果糖,可顯著提高短鏈低聚果糖的濃度,酶解條件為:菊粉質(zhì)量濃度為40 g/L,酶解pH值為5.0,溫度為50 ℃,酶用量為20 U/g(以菊粉質(zhì)量計(jì)),短鏈低聚果糖質(zhì)量濃度達(dá)到16.06g/L,是原酶液制備短鏈低聚果糖的2.55倍;短鏈低聚果糖得率為40.16%。
菊粉酶;選擇性吸附;菊芋渣;菊粉;短鏈低聚果糖
低聚果糖(FOSs),是通過(guò)β-2,1-糖苷鍵在果糖殘基的C1位上連接1~3個(gè)果糖分子所形成的混合物,聚合度(Dp)在3~9范圍內(nèi)[1]。短鏈FOSs的聚合度為3~5,主要在近段結(jié)腸位置被利用[1],能夠促進(jìn)腸道內(nèi)有益菌如雙歧桿菌的增殖,還具有防止齲齒和降血糖、膽固醇等生理作用[2]。因此,F(xiàn)OSs作為一種可溶性膳食纖維,是重要的功能性食品原料之一[3]。FOSs的生產(chǎn)主要有兩種方法:一是以蔗糖為原料,在高濃度底物存在的條件下,蔗糖在β-果糖轉(zhuǎn)移酶的作用下發(fā)生轉(zhuǎn)移反應(yīng),所得產(chǎn)物為GFn型FOSs(G為葡萄糖,F(xiàn)為果糖,n為果糖分子數(shù)),但在制備過(guò)程中會(huì)積累大量的副產(chǎn)物葡萄糖,進(jìn)而抑制果糖基轉(zhuǎn)移酶的產(chǎn)生[4],并且 FOSs純度不高[5],工藝繁雜,成本較高;二是以菊粉作為底物,內(nèi)切菊粉酶隨機(jī)切割長(zhǎng)鏈菊粉分子,所得產(chǎn)物為Fm型(F為果糖分子,m為果糖的分子數(shù))與GFn型的混合物,此法的產(chǎn)品純度較高,工藝簡(jiǎn)單,成本較低,是目前主流生產(chǎn)FOSs的方法[6]。國(guó)外以菊粉加工FOSs的公司主要有比利時(shí)的Orafti公司和Cosucra公司,以及美國(guó)的Jarrow Formulas公司[7]。其中Orafti公司所生產(chǎn)的低聚果糖純度達(dá)到93%~97%[8],現(xiàn)有的研究報(bào)道中基于菊粉的酶法制備FOSs的得率達(dá)到60%~86%[1]。以菊粉為原料生產(chǎn)FOSs,關(guān)鍵在于內(nèi)切菊粉酶的制備。然而,天然菊粉酶是由內(nèi)切菊粉酶和外切菊粉酶組成的復(fù)合酶系,菊粉在內(nèi)切菊粉酶的作用下主要產(chǎn)物為低聚果糖,而低聚果糖則在外切菊粉酶的作用下最終會(huì)水解成為果糖和葡萄糖[9]。因此,內(nèi)、外切菊粉酶的拆分對(duì)生產(chǎn)FOSs非常重要。天然菊粉酶酶系中內(nèi)切菊粉酶和外切菊粉酶均表現(xiàn)一定的菊粉酶活力和轉(zhuǎn)化酶活力,多數(shù)外切菊粉酶比內(nèi)切菊粉酶表現(xiàn)出更高的轉(zhuǎn)化酶活力[10]。因此,天然菊粉酶活力的高低常以菊粉酶活力(I)與轉(zhuǎn)化酶活力(S)的比值來(lái)表示,即I/S比值。拆分內(nèi)、外切菊粉酶的傳統(tǒng)方法主要有超濾、鹽析、柱層析、電泳等[11],這些方法都存在效率低、成本高、不易于工業(yè)化生產(chǎn)等缺點(diǎn)。本研究以黑曲霉產(chǎn)的天然菊粉酶為研究對(duì)象,采用菊粉加工廢棄物(菊芋渣)為吸附劑來(lái)選擇性吸附天然菊粉酶,降低天然菊粉酶酶系中的轉(zhuǎn)化酶活力,保留較多的菊粉酶活力,即提高I/S比值,以期通過(guò)低成本的選擇性吸附方法改變天然菊粉酶的酶系結(jié)構(gòu),提高低聚果糖的產(chǎn)率,為低聚果糖的工業(yè)化生產(chǎn)提供理論基礎(chǔ)。
1 材料與方法
1.1 材料
黑曲霉NLX-H164,南京林業(yè)大學(xué)生物化工研究所提供。純菊粉,購(gòu)自Beneo Orafit公司,菊芋渣由青海威德生物技術(shù)有限公司提供。其他化學(xué)品均為市售分析純。
1.2 方法
1.2.1天然菊粉酶的制備 在250 mL錐形瓶中裝入50 mL 產(chǎn)酶培養(yǎng)基,產(chǎn)酶培養(yǎng)基由20 g/L菊粉和10 g/L酵母粉構(gòu)成。接入5 mL黑曲霉NLX-H164,在170 r/min、30 ℃條件下培養(yǎng)。發(fā)酵72 h后取出發(fā)酵液,5 000 r/min下離心10 min,上清液即為天然菊粉酶酶液。
1.2.2菊芋渣選擇性吸附天然菊粉酶 菊芋渣于60 ℃烘干至質(zhì)量恒定,再經(jīng)粉碎至粒徑≤0.85 mm,所得固形物于4℃冷藏待用。取一定質(zhì)量的菊芋渣于錐形瓶中,加入0.1 mL的天然菊粉酶酶液,用不同pH值的乙酸-乙酸鈉緩沖溶液定容至20 mL。攪拌至均勻后,在80 r/min條件下恒溫吸附。吸附30 min后取樣,樣品于5 000 r/min離心10 min,收集上清液,測(cè)定上清液中菊粉酶活力(I)和轉(zhuǎn)化酶活力(S),計(jì)算I/S比值。菊芋渣對(duì)菊粉酶活力或轉(zhuǎn)化酶活力的選擇性吸附效果,以酶活力回收率表示,酶活力回收率=上清液中的酶活力(U)/起始加入的酶活力(U)×100 %。其中,酶活力單位(U)的定義為每分鐘生成1 μmol還原糖所需要的酶量為一個(gè)酶活力單位。
1.2.3菊粉酶活力(I)的測(cè)定 用0.1 mol/L、pH值4.6的乙酸-乙酸鈉緩沖溶液配制5 %菊粉溶液450 μL,加入50 μL的天然菊粉酶酶液,混勻后,60 ℃水浴中反應(yīng)10 min,反應(yīng)結(jié)束后,即刻取出并用沸水浴滅活5 min,從反應(yīng)液中取出200 μL,加入1.5 mL DNS試劑和1.8 mL水,混勻后,沸水浴反應(yīng)5 min,冷水冷卻后,用蒸餾水在試管中定容至25 mL,測(cè)定520 nm處的吸光度值。根據(jù)果糖標(biāo)準(zhǔn)曲線計(jì)算得出樣品溶液中的還原糖質(zhì)量(mg),進(jìn)而計(jì)算酶活力。
1.2.4轉(zhuǎn)化酶活力(S)的測(cè)定 用0.1 mol/L、pH值4.6的乙酸-乙酸鈉緩沖液配制5 %蔗糖溶液450 μL,取50 μL酶液加入混勻后,60 ℃水浴中反應(yīng)10 min,反應(yīng)結(jié)束后,即刻取出并用沸水浴滅活5 min,從反應(yīng)液中取出200 μL,加入1.5 mL DNS試劑和1.8 mL水,混勻后,沸水浴反應(yīng)5 min,冷水冷卻后,用蒸餾水在試管中定容至25 mL,測(cè)定520 nm處的吸光值。根據(jù)果糖標(biāo)準(zhǔn)曲線計(jì)算得出樣品溶液中的還原糖質(zhì)量(mg),進(jìn)而計(jì)算酶活力。
1.2.5酶法制備短鏈低聚果糖 稱取2.0 g菊粉于100 mL酶解瓶中,加入適量吸附處理的天然菊粉酶酶液,補(bǔ)加乙酸-乙酸鈉緩沖液(pH值5.0)至50 mL,溫度為50 ℃,酶用量為20 U/g(以菊粉質(zhì)量計(jì))在150 r/min條件下進(jìn)行酶水解。間隔2 h取樣,樣品于10 000 r/min離心10 min,收集上清液,測(cè)定上清液中短鏈FOSs的質(zhì)量濃度。酶解效果以短鏈FOSs得率表示,短鏈FOSs得率=總短鏈FOSs質(zhì)量/底物菊粉質(zhì)量×100%。
1.2.6分析方法 短鏈FOSs質(zhì)量濃度分析參考文獻(xiàn)[12],并有一定的改進(jìn)。短鏈FOSs的聚合度一般為3~5[1]。采用戴安離子色譜系統(tǒng)測(cè)定上清液中的短鏈FOSs質(zhì)量濃度,離子色譜條件為:色譜柱為CarboPac PA10,柱溫30 ℃,流速0.3 mL/min。以超純水、200 mmol/L NaOH 和 500 mmol/L NaAc為流動(dòng)相,洗脫梯度:0~13 min,80 %水和20 % 200 mmol/L NaOH;13~30 min,90 %~85 % 200 mmol/L NaOH 和10 %~15 % 500 mmol/L NaAc;30~55 min,80 %水和 20 % 200 mmol/L NaOH。
2 結(jié)果與討論
2.1 菊芋渣吸附條件對(duì)菊粉酶活力的影響
2.1.1菊芋渣質(zhì)量濃度 菊芋渣作為吸附劑,其濃度的高低會(huì)影響天然菊粉酶酶系的選擇性吸附。隨著菊芋渣質(zhì)量濃度的增加,體系中的固形物含量隨之提高,能夠增加吸附底物與酶組分的接觸,提高吸附效率。在吸附溫度4 ℃、pH值4.6條件下,研究菊芋渣質(zhì)量濃度對(duì)菊芋渣選擇性吸附菊粉酶的影響,如圖1(a)所示。
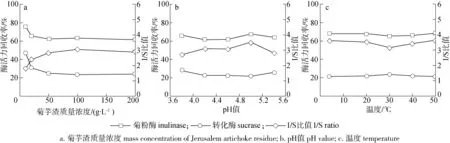
圖1 菊芋渣吸附條件對(duì)菊芋渣吸附菊粉酶的影響Fig. 1 Effect of adsorption conditions on the adsorption of inulinase by Jerusalem artichoke residue
由圖1(a)可知,隨著菊芋渣質(zhì)量濃度由10 g/L上升至200 g/L,菊粉酶和轉(zhuǎn)化酶的酶活力回收率變化趨勢(shì)一致,即隨著吸附劑質(zhì)量濃度的增加,酶活力回收率也逐漸降低,并趨于平衡。當(dāng)菊芋渣質(zhì)量濃度由10 g/L上升至100 g/L時(shí),菊粉酶和轉(zhuǎn)化酶酶活力回收率下降較快,I/S比值隨菊芋渣質(zhì)量濃度的增加而增加;當(dāng)菊芋渣質(zhì)量濃度大于100 g/L時(shí),菊粉酶和轉(zhuǎn)化酶的酶活力回收率趨于平穩(wěn),I/S比值也趨于平穩(wěn)。在體系內(nèi)酶蛋白一定的條件下,增加吸附劑用量,酶蛋白吸附量增加,當(dāng)酶蛋白吸附飽和后,進(jìn)一步增加吸附劑用量,酶蛋白吸附量不再增加[13]。同時(shí),菊粉酶酶活力回收率明顯高于轉(zhuǎn)化酶的酶活力回收率,說(shuō)明菊芋渣對(duì)菊粉酶和轉(zhuǎn)化酶具有選擇性吸附,菊芋渣優(yōu)先吸附轉(zhuǎn)化酶。當(dāng)菊芋渣質(zhì)量濃度達(dá)100 g/L時(shí),I/S比值達(dá)到最大值,為3.06。因此,選擇的最佳吸附劑質(zhì)量濃度為100 g/L。
2.1.2pH值 pH值不僅影響吸附劑的表面性質(zhì),也影響吸附質(zhì)所帶電荷,特別是蛋白質(zhì)等吸附質(zhì)所帶電荷受pH值影響較大,過(guò)高和過(guò)低的pH值均對(duì)吸附不利[12]。因此,選擇合適的吸附pH值對(duì)菊芋渣吸附菊粉酶具有重要的作用。文獻(xiàn)表明黑曲霉產(chǎn)菊粉酶的最適pH值在4.0~5.0之間[11]。因此,在吸附溫度4 ℃、菊芋渣質(zhì)量濃度100 g/L條件下,研究pH值對(duì)菊芋渣吸附菊粉酶的影響,如圖1(b)所示。由圖1(b)可知,在pH值3.8~5.4范圍內(nèi),菊粉酶與轉(zhuǎn)化酶的酶活力回收率呈現(xiàn)不同的趨勢(shì),菊粉酶的酶活力回收率表現(xiàn)為先降低后升高再降低的趨勢(shì),而轉(zhuǎn)化酶的酶活力回收率表現(xiàn)為先降低后升高的趨勢(shì),說(shuō)明菊粉酶與轉(zhuǎn)化酶因蛋白質(zhì)組成的不同,在不同的pH值條件下,所帶電荷存在差異,進(jìn)而影響吸附效果,表現(xiàn)為選擇性吸附。在pH值為5.0時(shí),菊粉酶的酶活力回收率達(dá)到最高為67.62 %,而轉(zhuǎn)化酶的酶活力回收率最低為21.69 %,I/S比值達(dá)到最大值為3.53。因此,菊芋渣吸附菊粉酶的最佳pH 值為5.0,此時(shí)菊粉酶中的轉(zhuǎn)化酶活力被最大限度的去除。
2.1.3溫度 吸附溫度對(duì)于吸附的影響和吸附過(guò)程的熱效應(yīng)相關(guān)[14]。同時(shí),溫度也對(duì)酶活力產(chǎn)生影響。在菊芋渣質(zhì)量濃度100 g/L、pH值5.0條件下,研究溫度對(duì)菊芋渣吸附菊粉酶的影響,如圖1(c)所示。由圖1(c)可以看出,在4~50 ℃范圍內(nèi),吸附溫度的升高對(duì)菊粉酶與轉(zhuǎn)化酶的酶活力回收率影響較小。在4℃時(shí),菊粉酶的酶活力回收率為67.91 %,轉(zhuǎn)化酶的酶活力回收率為21.33 %,I/S比值為3.61;而在30 ℃時(shí),菊粉酶的酶活力回收率為65.32 %,轉(zhuǎn)化酶的酶活力回收率為23.19 %,I/S比值為3.18。吸附過(guò)程一般為放熱過(guò)程,溫度對(duì)吸附的影響為負(fù)效應(yīng),即低溫有利于吸附[15]。同時(shí),溫度過(guò)高影響菊粉酶的生物活性。因此,選擇吸附溫度為4 ℃。
2.2 菊芋渣選擇性吸附前后菊粉酶酶活力的變化
經(jīng)優(yōu)化后確定菊芋渣選擇性吸附天然菊粉酶的較佳吸附條件為:菊芋渣質(zhì)量濃度100 g/L,吸附pH值5.0,吸附溫度4 ℃。在此條件下,對(duì)天然菊粉酶吸附前后的性質(zhì)進(jìn)行比較,結(jié)果如表1所示。

表 1 菊芋渣選擇性吸附前后天然菊粉酶性質(zhì)的對(duì)比Table 1 Changes of native inulinase by the selective adsorption of Jerusalem artichoke residue
由表1可知,經(jīng)菊芋渣吸附后,菊粉酶活力由11.78 U/mL下降至8.00 U/mL,而轉(zhuǎn)化酶活力從10.39 U/mL降至1.99 U/mL;相應(yīng),I/S比值由1.13上升至4.02,增加了255.75 %。菊芋渣吸附天然菊粉酶表現(xiàn)為選擇性吸附,優(yōu)先吸附轉(zhuǎn)化酶活力,吸附80.84 %的轉(zhuǎn)化酶活力,對(duì)應(yīng)的菊粉酶活力僅被吸附32.08 %,使得吸附后的天然菊粉酶酶系中的轉(zhuǎn)化酶活力顯著降低,保留了較多的菊粉酶活力。
2.3 酶法制備短鏈低聚果糖
利用菊芋渣選擇性吸附處理前后的天然菊粉酶來(lái)水解菊粉制備短鏈低聚果糖(FOSs),按1.2.5節(jié)操作,如圖2所示。由圖2可知,40 g/L菊粉溶液本身含有少量的短鏈FOSs,質(zhì)量濃度為5.77 g/L。吸附后的天然菊粉酶水解菊粉過(guò)程中,隨著時(shí)間的延長(zhǎng),生成的短鏈FOSs不斷增加,酶解10 h后,短鏈FOSs質(zhì)量濃度和得率達(dá)到最大值,分別為16.06 g/L和40.16 %;進(jìn)一步增加酶解時(shí)間,短鏈FOSs質(zhì)量濃度和得率開(kāi)始下降。原酶液的酶水解過(guò)程中,酶解10 h,短鏈FOSs質(zhì)量濃度從5.77 g/L增加至6.31 g/L,僅增加9.36 %,選擇性吸附的天然菊粉酶制備的短鏈FOSs質(zhì)量濃度是原酶液的2.55倍。天然菊粉酶經(jīng)菊芋渣選擇性吸附后,多數(shù)轉(zhuǎn)化酶活力被吸附,說(shuō)明從天然菊粉酶酶系中去除較多的外切菊粉酶,而內(nèi)切菊粉酶保留較多。利用菊芋渣吸附的天然菊粉酶制備短鏈FOSs的結(jié)果進(jìn)一步證明吸附后的天然菊粉酶酶系中存在較多的內(nèi)切菊粉酶,能夠隨機(jī)切割長(zhǎng)鏈菊粉分子,生成短鏈FOSs,增加了短鏈FOSs質(zhì)量濃度,提高了短鏈FOSs的得率。因此,采用菊粉加工產(chǎn)生的廢棄物(菊芋渣),經(jīng)一步法選擇性吸附天然菊粉酶,用以制備短鏈FOSs,具有步驟少、成本低、食品安全性高等優(yōu)點(diǎn)。同時(shí),短鏈FOSs得率仍然存在較大的提升空間,可通過(guò)對(duì)菊芋渣修飾提高對(duì)天然菊粉酶的選擇性吸附,或結(jié)合蛋白質(zhì)沉淀方法,進(jìn)一步處理吸附后的天然菊粉酶,提高I/S比值,從而提高短鏈FOSs質(zhì)量濃度和得率。
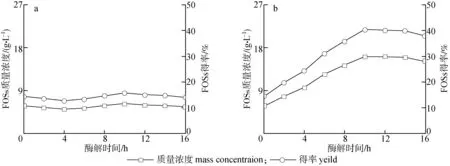
圖 2 菊芋渣吸附前(a)和吸附后(b)的天然菊粉酶制備短鏈低聚果糖(FOSs)Fig. 2 Preparation of short-chain fructooligosaccharides(FOSs) by the native inulinase before(a) and after(b) adsorbed by Jerusalem artichoke residue
3 結(jié) 論
3.1利用菊粉加工廢棄物(菊芋渣)選擇性吸附黑曲霉產(chǎn)天然菊粉酶,制備低轉(zhuǎn)化酶活力的菊粉酶,較佳吸附條件為:菊芋渣質(zhì)量濃度為100 g/L,吸附pH值為5.0,吸附溫度為4 ℃。經(jīng)菊芋渣吸附后,天然菊粉酶酶系中的轉(zhuǎn)化酶活力顯著降低,菊粉酶活力保留較多。利用吸附的天然菊粉酶制備短鏈低聚果糖,酶解條件為菊粉質(zhì)量濃度為40 g/L,酶解pH值為5.0,溫度為50 ℃,酶用量為20 U/g(以菊粉質(zhì)量計(jì)),短鏈低聚果糖質(zhì)量濃度較之原酶液高2.55倍,菊芋渣選擇性吸附天然菊粉酶,去除較多的外切菊粉酶活力,保留較多的內(nèi)切菊粉酶活力,大幅度提高短鏈低聚果糖質(zhì)量濃度和得率。
3.2利用菊粉加工廢棄物建立的選擇性吸附天然菊粉酶的方法,改變天然菊粉酶的酶系結(jié)構(gòu),有效提高短鏈低聚果糖的質(zhì)量濃度,為高效、低成本生產(chǎn)短鏈低聚果糖提供一種綠色、安全的新思路。
[1]SINGH R S,SINGH R P,KENNEDY J F. Recent insights in enzymatic synthesis of fructooligosaccharides from inulin[J]. International Journal of Biological Macromolecules,2016,85:565-572.
[2]MUTANDA T,MOKOENA M P,OLANIRAN A O,et al. Microbial enzymatic production and applications of short-chain fructooligosaccharides and inulooligosaccharides:Recent advances and current perspectives[J]. Journal of Industrial Microbiology and Biotechnology,2014,41(6):893-906.
[3]李娟,曹澤虹,李超,等. 響應(yīng)面法優(yōu)化黑曲霉深層發(fā)酵產(chǎn)內(nèi)切型菊粉酶工藝[J]. 食品科學(xué),2014,35(9):207-212.
[4]覃益民,唐江濤,蘇加坤,等. 米曲霉 GX0011β-果糖基轉(zhuǎn)移酶的性質(zhì)研究[J]. 食品工業(yè)科技,2007,28(5):123-126.
[5]JONY W Y,SEUNG K S. The production of high-content fructo-oligosaccharides from sucrose by the mixed-enzyme system of fructosyltransferase and glucose oxidase[J]. Biotechnology Letters,1993,15(6):573-576.
[6]PARK S,JEONG H Y,KIM H S,et al. Enhanced production ofAspergillusficuumendoinulinase inSaccharomycescerevisiaeby using the SUC2-deletion mutation[J]. Enzyme and Microbial Technology,2001,29(2):107-110.
[7]SINGH R S, SINGH R P. Production of fructooligosaccharides from inulin by endoinulinases and their prebiotic potential [J]. Food Technology and Biotechnology,2010,48(4):435-450.
[8]任瑋. 產(chǎn)菊粉酶微生物水解菊芋粉的應(yīng)用研究[D]. 無(wú)錫:江南大學(xué)碩士學(xué)位論文,2008.
[9]鄭重. 內(nèi)切菊粉酶的制備,純化及應(yīng)用研究[D]. 南京:南京林業(yè)大學(xué)碩士學(xué)位論文,2010.
[10]ETTALIBI M,BARATTI J C. Purification,properties and comparison of invertase,exoinulinases and endoinulinases ofAspergillusficuum[J]. Applied Microbiology and Biotechnology,1987,26(1):13-20.
[11]陳曉明,陳寒青,陳天祥,等. 菊粉酶的酶學(xué)特性與分子生物學(xué)[J]. 食品與生物技術(shù)學(xué)報(bào),2009,28(5):577-588.
[12]XU Y B,ZHENG Z J,XU Q Q,et al. Efficient conversion of inulin to inulooligosaccharides through endoinulinase fromAspergillusniger[J]. Journal of Agricultural and Food Chemistry,2016,64:2612-2618.
[13]徐威,黃海華,梁麗莉. 吸附法純化青霉素酰化酶[J]. 沈陽(yáng)藥科大學(xué)學(xué)報(bào),2002,19(6):443-447.
[14]吳志堅(jiān),劉海寧,張慧芳. 離子強(qiáng)度對(duì)吸附影響機(jī)理的研究進(jìn)展[J]. 環(huán)境化學(xué),2010,29(6):997-1003.
[15]謝祖芳,何星存,夏金虹,等. 苦味酸在聚酰胺樹(shù)脂上的吸附熱力學(xué)及動(dòng)力學(xué)[J]. 化學(xué)研究,2003,14(4):53-56.
Selective Adsorption of Native Inulinase by Jerusalem Artichoke Residue andPreparation of Short-chain Fructooligosaccharides
LI Xin1, 2, ZHOU Jin2, LAI Chenhuan1, 2, YONG Qiang1, 2
(1. Jiangsu Co-Innovation Center of Efficient Processing and Utilization of Forest Resources,Nanjing Forestry University, Nanjing 210037, China; 2. College of Chemical Engineering,Nanjing Forestry University, Nanjing 210037, China)
A selective adsorption method was developed to adsorb native inulinase fromAspergillusnigerto reduce sucrase activity by Jerusalem artichoke residue. The inulinase activity(I)/sucrase activity(S) ratio was generally used to characterize the inulinases. Afterward, the native inulinase, selectively adsorbed by Jerusalem artichoke residue, was applied for production of short-chain fructooligosaccharides(FOSs). The results showed that the optimal conditions of the selective adsorption were as follows: Jerusalem artichoke residue loading 100 g/L, pH value 5.0, adsorption temperature 4 ℃. 67.92% Inulinase activity was retained and 80.84% sucrase activity was decreased in the native inulinase system after the selective adsorption. The I/S ratio increased from 1.13 to 4.02. The selective adsorption of native inulinase by Jerusalem artichoke residue improved the production of short-chain FOSs. The conditions of enzymatic hydrolysis were inulin 40 g/L, pH value 5.0, 50 ℃, inulinase loading 20 U/g(based on the mass of inulin) . The mass concentration of short-chain FOSs was 16.06 g/L, which was 2.55 times than that of the original inulinase. The yield of short-chain FOSs was 40.16%.
inulinase;selective adsorption;Jerusalem artichoke residue;inulin;short-chain fructooligosaccharides
TQ35;Q815
A
1673-5854(2017)06- 0033- 05
10.3969/j.issn.1673-5854.2017.06.006
2017- 10-16
江蘇省重點(diǎn)研發(fā)計(jì)劃項(xiàng)目(BF2015007);青海省重點(diǎn)研發(fā)與轉(zhuǎn)化計(jì)劃項(xiàng)目(2016-HZ-819);江蘇省高校優(yōu)勢(shì)學(xué)科建設(shè)工程資助項(xiàng)目(無(wú)編號(hào))
李 鑫 (1975— ),男,遼寧大連人,副教授,博士,主要從事生物質(zhì)資源生物利用研究工作;E-mailxli@njfu.edu.cn。
猜你喜歡
中學(xué)生數(shù)理化·中考版(2022年10期)2022-11-10 09:37:42
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2022年12期)2022-02-14 07:08:42
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2021年12期)2021-12-31 03:23:08
中學(xué)生數(shù)理化·中考版(2020年10期)2020-11-27 01:59:48
中國(guó)生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(shè)(2018年6期)2018-04-22 03:16:54
產(chǎn)品可靠性報(bào)告(2017年7期)2017-09-05 09:49:12
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
