堿度對水產養殖絮體生物學特性及氨氮轉化的影響
2017-12-18 02:50:33羅國芝譚洪新
水產科學 2017年4期
關鍵詞:生物
馬 濤,羅國芝,2,3,譚洪新,2,3 ,陳 偉
( 1.上海海洋大學 水產與生命學院,上海 201306; 2.上海水產養殖工程技術研究中心,上海 201306;3.上海高校知識服務平臺 上海海洋大學水產動物育種中心,上海 201306 )
堿度對水產養殖絮體生物學特性及氨氮轉化的影響
馬 濤1,羅國芝1,2,3,譚洪新1,2,3,陳 偉1
( 1.上海海洋大學 水產與生命學院,上海 201306; 2.上海水產養殖工程技術研究中心,上海 201306;3.上海高校知識服務平臺 上海海洋大學水產動物育種中心,上海 201306 )
在異位式生物絮凝反應器中,添加鰻鱺循環水養殖系統中收集的殘餌糞便進行生物絮體培養,用碳酸氫鈉和1 mol/L HCl控制反應器中的堿度水平(以CaCO3計)為:a組≥250 mg/L、b組150~200 mg/L、c組75~100 mg/L,研究堿度對絮體形成過程中3種形態氮含量、絮體的氮素轉化及粗蛋白、粗脂肪和胞外聚合物含量的影響,用IlluminaMiseq測序技術檢測了絮體微生物群落的多樣性。試驗結果表明,堿度150~200 mg/L試驗組絮體氮素轉化率[(70.84±7.67)%]、粗蛋白含量[(36.74±0.59)%]、疏松結合胞外聚合物和緊密結合胞外聚合物中蛋白質與多糖比和絮體原核、真核微生物群落 Shannon多樣性指數均優于其他試驗組。將堿度提高至逾150 mg/L,可促使反應器中的硝化作用。3組絮體中具有顯著性差異的原核微生物優勢菌群為變形菌門和Saccharibacteria門,3個試驗組中變形菌門的比例分別為45.96%、27.81%、80.07%;絮體真核微生物具有顯著性差異的優勢菌群為子囊菌門和纖毛蟲門,3個試驗組中纖毛蟲門的比例分別為7.24%、0.43%、14.85%。異位式生物絮體培養過程中堿度應控制在150 mg/L以上時,絮體微生物多樣性增加,氮素轉化效率增高。
堿度;殘餌糞便;生物絮體;IlluminaMiseq測序;微生物群落
近年來,水產養殖產業迅速發展,高密度集約化養殖規模日益擴大,養殖廢水及水產動物排泄物肆意排放等問題嚴重制約著水產養殖業的健康穩定發展,也污染了生態環境。研究證明,生物絮凝技術可調控水體營養結構,促進異養細菌的繁殖,利用微生物同化無機氮轉化為細菌蛋白,凈化水質、減少換水量;通過細菌絮凝成顆粒,成為部分濾食性養殖對象的食物,降低飼料系數;提高動物免疫和生態防病,是解決水產養殖業發展的環境制約和飼料成本的有效技術。
在水產生物的絮體形成中,異養細菌和硝化細菌消耗無機碳源,使堿度(以CaCO3計)、pH降低[1-2]。從絮體形成的理論方程式:NH4++1.18C6H12O6+HCO3-+2.06O2→C5H7O2N+6.06H2O+3.07CO2,可知異養微生物轉化1 g氨氮需消耗3.57 g堿度(0.86 g無機碳)[3]。自養細菌的硝化作用在生物絮凝系統中很常見[4-5]。理論上完全硝化1 g氨氮需要消耗7.07 g堿度(1.69 g無機碳)[6],主要消耗在氨氮氧化為亞硝態氮過程,因此補充足夠的堿度是保證異養細菌和硝化細菌完成生物絮凝的前提。
Boyd等[7]發現,堿度只要高于75 mg/L就可以形成絮體;Chen等[6]認為,水產生物絮凝系統中堿度應維持在100~150 mg/L,高于200 mg/L時有益于絮體的形成。Furtado等[8]研究了生物絮凝系統中堿度對水質的影響,證實當堿度低于100 mg/L,pH<7時,水體中氨氮和亞硝態氮含量比堿度為150~300 mg/L條件下高。上述研究均未見堿度對生物絮體微生物的影響。本研究利用花鰻鱺(Anguillamarmorata)循環水養殖系統固體廢棄物,通過向異位序批式生物絮凝反應器中添加碳酸氫鈉控制反應器中堿度來培養絮體,監測堿度對生物絮體形成過程中氮素轉化、絮體的生物學特性及微生物群落結構的影響,為生物絮凝技術在水產養殖中的應用提供參考。
1 材料與方法
1.1 材料
試驗共用9個相同的圓柱形聚乙烯生物絮凝反應器(內徑15 cm,高15 cm,有效容積11 L),每個反應器裝一個石英沙聚合曝氣石(直徑3 cm,高3.5 cm),3個曝氣石連接到一臺電磁式空氣壓縮機(型號ACO-008,135 W,100 L/min,浙江森森有限公司)。
殘餌糞便取自上海海洋大學花鰻鱺循環水養殖系統的固液分離裝置,投喂鰻魚黑仔配合飼料。飼料水分≤10.0%,粗蛋白質≥48.0%,粗脂肪≥4.0%,粗纖維≤3.0%,粗灰分≤17.0%,總磷1.0%~2.8%(數據由廠家提供)。收集的殘餌糞便經過65 ℃烘干,測得粗蛋白質(34.25±0.20)%,粗脂肪(6.55±0.71)%,粗灰分(61.91±0.4)%。
試驗開始前,向一聚乙烯桶中添加適量殘餌糞便和曝氣自來水,混勻,測得初始總懸浮固體物含量為3500 mg/L,之后轉移到反應器中培養30 d。添加葡萄糖為碳源,維持反應器中碳氮比15以上,以溶解有機碳∶氨氮含量計算,溫度23~25 ℃,溶解氧6.5~8.6 mg/L。
通過碳酸氫鈉和1 mol/L HCl的比例控制反應器中堿度(以CaCO3在3個處理組水平:a組≥250 mg/L; b組150~200 mg/L; c組75~100 mg/L;每組設3個重復)。試驗期間每日向反應器中加純水至10 L,補充蒸發和采樣損失的水。
1.2 測定指標與方法
試驗期間,每日9:00用Multi3430型多參數水質分析儀測定反應器中溫度、溶解氧含量和pH。試驗前7 d每日9:00從系統取水經0.45 μm濾膜過濾后,測氨氮、亞硝態氮、硝態氮、溶解有機碳含量,系統穩定后每2 d測定一次。氨氮含量用納氏試劑分光光度法(型號UV2000,上海優尼科,中國)、亞硝態氮含量采用鹽酸萘乙二胺比色法、硝態氮含量采用N-(1-萘基)-紫外分光光度法、溶解有機碳含量采用總有機碳分析儀測定(N/C?2100,analytikjenamulti,德國)。每日9:00和20:00用酸堿指示劑滴定法測定系統堿度變化并進行調節。
每2 d測定反應器中總懸浮固體顆粒物、揮發性懸浮固體含量、生物絮體沉降性能,觀察絮體形態、提取疏松結合胞外聚合物和緊密結合胞外聚合物;測定絮體粗蛋白、粗脂肪和粗灰分含量。總懸浮固體顆粒物和揮發性懸浮固體含量采用稱量質量法測定。殘餌糞便和絮體經65 ℃干燥后使用元素分析儀測定碳、氮含量。粗灰分通過馬福爐550 ℃灼燒4 h測定。粗脂肪采用氯仿—甲醇溶液(2∶1,體積比)抽提法測定。粗蛋白、粗脂肪和粗灰分含量均按照絮體干質量測定和計量(%)。在奧林帕斯體視鏡(Olympus SZ2-STS)、掃描電子顯微鏡(S3400NII,日本日立公司,日本)下觀察絮體的形態、拍照。參照文獻[9]的方法提取及測定胞外聚合物的含量。
試驗第30 d取生物絮體進行原核及真核微生物高通量測序。使用E.Z.N.A Soil DNA試劑盒(美國OMEGA Biotek公司)提取絮體基因組DNA,針對16S rRNA、18S rRNA基因,合成帶有barcode的特異引物。PCR采用TransGen AP221-02:TransStartFastpfu DNA聚合酶反應體系。每個樣本3個重復,將同一樣本的PCR產物混合后用2%瓊脂糖凝膠電泳檢測,使用AxyPrepDNA凝膠回收試劑盒(AXYGEN公司)切膠回收PCR產物,TrisHCl洗脫;2%瓊脂糖電泳檢測。參照電泳初步定量結果,用QuantiFluorTM-ST藍色熒光定量系統(Promega公司)定量檢測PCR產物,按照每個樣本的測序量要求,進行相應比例的混合,之后采用Illumina公司的Miseq進行測序分析(上海美吉生物醫藥科技有限公司完成)。
2 結 果
2.1 3組系統堿度的控制及水中3種形態氮含量的變化
將混勻的殘餌糞便添加到反應器中時3組水體堿度相同,初始平均堿度為355.56 mg/L,加酸使3組堿度分別達到試驗設置水平(圖1)。2~6 d 3組系統堿度持續降低,每日分別需補充碳酸氫鈉,第3 d 3組平均降至41.95 mg/L。第4 d,堿度開始緩慢回升,堿度75~100 mg/L組開始穩定,其他兩組還需要加堿,在第6 d后穩定且維持在試驗設置的水平。
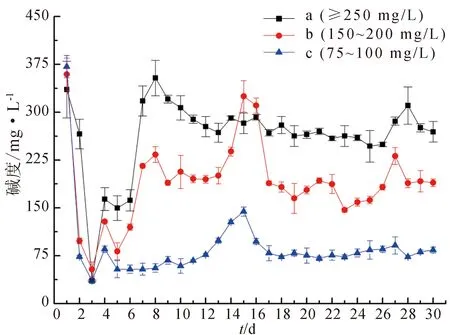
圖1 3組反應器水中堿度的變化
3組反應器運行期間3種形態氮質量濃度變化趨勢相同,但堿度75~100 mg/L組質量濃度與其他兩組間存在明顯差異。微生物對殘餌糞便的降解使得各反應器中氨氮釋放,第2 d堿度≥250 mg/L組和堿度150~200 mg/L組的氨氮質量濃度達最高(24.05 mg/L和31.93 mg/L),第6 d降至0.22 mg/L和0.15 mg/L,之后均保持在3.5 mg/L以下,啟動階段這兩組中氨氮的轉化率分別為88.5%和99.53%;堿度75~100 mg/L組氨氮變化速度較其他兩組緩慢,在第3 d達最高(37.20 mg/L),第7 d降至0.99 mg/L,轉化率為97.34%,至試驗結束時質量濃度維持在6 mg/L以下。各組反應器均發生了明顯的硝化作用,第5 d堿度≥250 mg/L組和堿度150~200 mg/L組亞硝態氮質量濃度達最高,為40.68 mg/L、43.50 mg/L,明顯高于堿度75~100 mg/L組(18.28 mg/L),之后前兩組亞硝態氮質量濃度快速下降,至第14 d 后趨于穩定,質量濃度均低于0.05 mg/L。第14、15 d堿度75~100 mg/L組堿度異常升高,亞硝態氮質量濃度升至23.20 mg/L,第18 d降低并穩定。試驗中硝態氮含量不斷變化,3個試驗組的最高質量濃度分別為38.30 mg/L、48.43 mg/L和30.37 mg/L,在第16 d開始發生了明顯的反硝化作用,未發生積累,至第20 d 3組硝態氮質量濃度均降至0.05 mg/L以下。

圖2 3個試驗組反應器水中3種形態氮質量濃度的變化
2.2 絮體氮素轉化率和營養成分含量
隨著糞便分解和氨化作用的進行,試驗第2 d 3個試驗組分別有氮(884.70±17.01) mg、(906.28±18.21) mg和(915.60±30.9) mg釋放在反應器中,試驗結束時,3個試驗組絮體中每克揮發性懸浮物分別含有氮(563.66±24.17) mg、(642.01±12.37 ) mg和(542.45±11.89) mg。氮素轉化率[10](終末絮體中每克揮發性懸浮物所含氮/初始總氮)分別為(63.71±4.98)%、(70.84±7.67)%、(59.25±6.36)%,說明生物絮凝可將水產養殖固體廢棄物中氮素有效轉化,堿度150~200 mg/L組高于其他兩組。
培養第14 d時絮體的粗蛋白、粗脂肪和粗灰分含量如表1。堿度≥250 mg/L組和堿度150~200 mg/L組粗蛋白含量高于培養初期殘餌糞便中的粗蛋白含量(34.25%),堿度150~200 mg/L組粗蛋白含量顯著高于其他兩組。3組絮體粗脂肪含量均比培養初期殘餌糞便粗脂肪含量高。3組絮體粗灰分含量明顯低于培養初期殘餌糞便粗灰分含量。
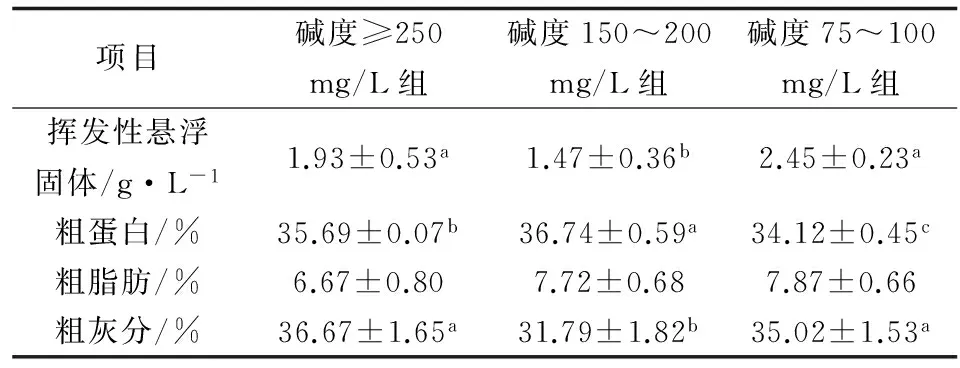
表1 第14 d不同堿度組絮體的粗蛋白、粗脂肪和粗灰分含量
注:表中同一列數據上標不同小寫字母的平均值間差異顯著(P<0.05),余同.
2.3 絮體的外觀形態
在顯微鏡和電鏡下觀察,培養初期的固體廢棄物是粒徑較大、緊致的顆粒,充滿了有機碎屑,隨著微生物的分解逐漸發展成為以絲狀菌、菌膠團為骨架的團狀物,結構松散,大小有幾微米到幾百微米,甚至超過1000 μm。堿度≥250 mg/L組和堿度150~200 mg/L組絮體結構松散、邊緣模糊,含大量絲狀菌;堿度75~100 mg/L組絮體粒徑較小、結構緊密、邊緣整齊,以菌膠團為主。對絮體形成動態進行觀察時發現,堿度75~100 mg/L組微生物的量明顯減少,但纖毛蟲類的數量增多,與真核域的檢測結果一致,堿度≥250 mg/L組和堿度150~200 mg/L組中含有原生動物的卵、藻類、輪蟲、砂殼蟲、線蟲較堿度75~100 mg/L組多(圖3,圖4)。
2.4 絮體胞外聚合物含量
利用超聲波—高速離心法提取生物絮體的疏松結合胞外聚合物和緊密結合胞外聚合物。第6 d 3組中氨氮含量降低時的胞外聚合物含量見表2。絮體胞外聚合物的主要組成部分是蛋白質和多糖,DNA 所占比例最少,蛋白質主要存在于緊密結合胞外聚合物中。包裹著疏松結合胞外聚合物的異養活性污泥表面負電性強且疏水性差,不利于生物絮凝,緊密結合胞外聚合物通過疏水性與菌細胞緊密結合在一起,絮凝性能和吸附污染物質的能力更強[11-12]。目前,較多研究發現,胞外聚合物的蛋白質與多糖質量濃度比與污泥表面性質有相關性,比值越高,越有利于絮狀污泥絮凝,其脫水性能越差[13]。堿度150~200 mg/L組絮體胞外聚合物蛋白質/多糖比值最高,且與其他兩組有顯著性差異。

圖3 第24 d時3組絮體在顯微鏡下的形態(×200倍)
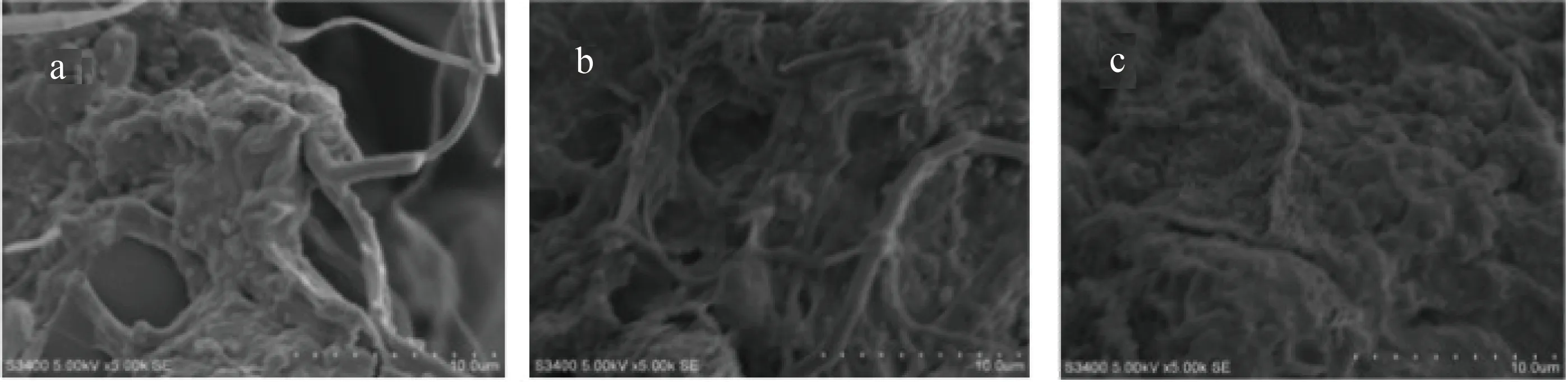
圖4 第24 d 時3組絮體在電子顯微鏡下的形態(×5000倍)
2.5 絮體微生物群落
2.5.1 樣品序列數目和多樣性
絮體細菌16S序列數目和平均長度分別為:堿度≥250 mg/L組,40 956條、434.38 bp;堿度150~200 mg/L組,32 623條、434.58 bp;堿度75~100 mg/L組,38 450條、443.76 bp。真核18S 序列數目和平均長度分別為:堿度≥250 mg/L組,3214條、400.85 bp;堿度150~200 mg/L組,29 155條、401.0 bp;堿度75~100 mg/L組,40 319條、400.59 bp。用Usearch對序列進行歸類操作,按照97%相似性得到OTU的代表序列,利用Mothur軟件計指數分析,通過單樣品的多樣性分析可以反映微生物的豐度和多樣性(表3)。細菌16S OTU數量和微生物豐度排序為:堿度≥250 mg/L組>堿度150~200 mg/L組>堿度75~100 mg/L組;真菌18S OTU數量和微生物豐度排序為:堿度75~100 mg/L組>堿度≥250 mg/L組>堿度150~200 mg/L組。Shannon值是估算樣品中微生物多樣性指數之一,其值越大,說明群落多樣性越高。利用3組絮體的測序量在不同測序深度時的微生物多樣性指數構建曲線,當曲線趨向平坦時,說明測序數據量足夠大,可以反映絮體中絕大多數的微生物物種信息。Shannon指數曲線表明(圖5),堿度150~200 mg/L組細菌16S多樣性指數最優,Shannon值最高4.35,Simpson值最低0.03;3組絮體真菌18S多樣性整體較低,相對于其他兩組,堿度150~200 mg/L組Shannon值最高1.4,Simpson值最低0.33,可知堿度150~200 mg/L組絮體原核、真核微生物多樣性最高。
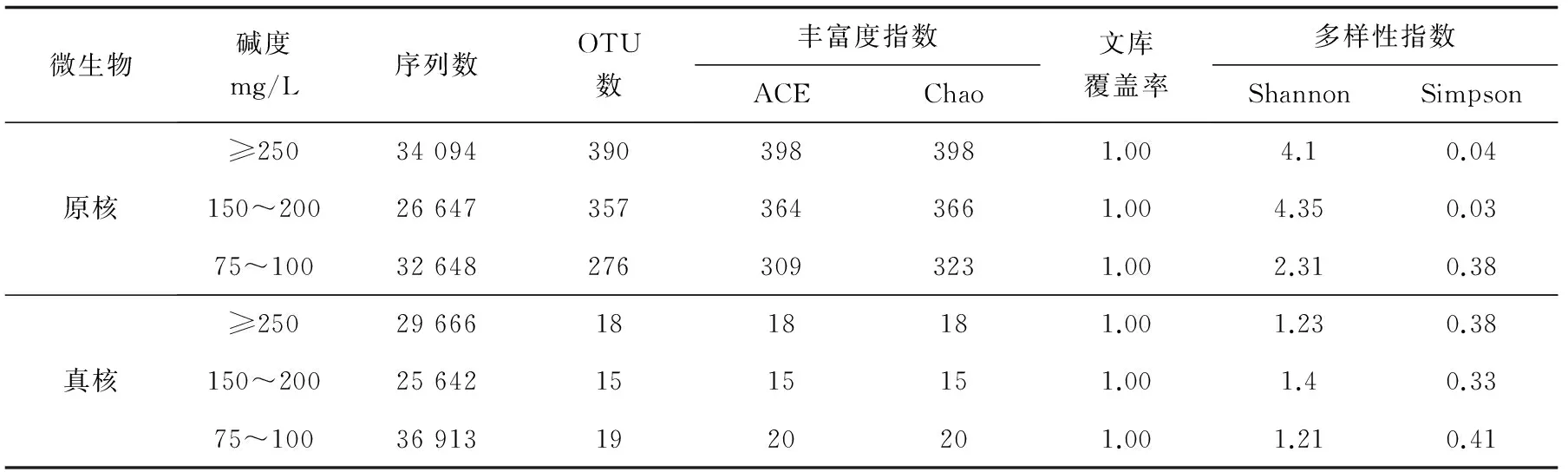
表3 不同堿度組絮體原核16S、真核18S的 OTU 數量及 alpha 多樣性
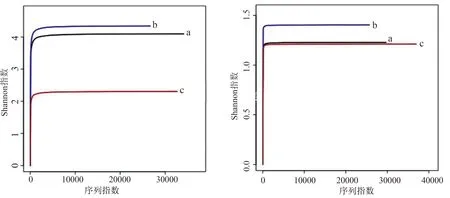
圖5 原核16S(左)、真核18S(右)多樣性Shannon 指數曲線
2.5.2 微生物群落組成分析
絮體中原核、真核微生物各門的分布情況見表4。由表4可見,3組絮體原核微生物主要隸屬于8個菌門,分別為變形菌門、Saccharibacteria門、擬桿菌門、綠彎菌門、放線菌門、厚壁菌門、疣微菌門、浮霉菌門,不同堿度的3組絮體所含微生物在門水平上相似,但堿度75~100 mg/L組所占比例與其他兩差異顯著。
A組生物絮體原核微生物包括205個菌屬,相對豐度大于1% 的有18個屬,其中相對豐度大于10%的2個菌屬,分別是Saccharibacteria_norank占21.2%和突柄微菌屬(Prosthecomicrobium)占12.28%;堿度150~200 mg/L組生物絮體原核微生物包括187個菌屬,相對豐度大于1% 的有19個屬,其中相對豐度大于10%的2個菌屬,分別為Saccharibacteria_norank占21.3%和Caldilineaceae_uncultured占12.36%;堿度75~100 mg/L組生物絮體原核微生物包括162個菌屬,相對豐度大于1%的有11個屬,其中相對豐度大于10%的菌屬有1個,是克雷伯氏菌屬(Klebsiella)占61.19%。3組生物絮體真核微生物所包含的屬分別為: 堿度≥250 mg/L組,18個;堿度150~200 mg/L組,15個;堿度75~100 mg/L組,16個。3組中優勢菌群及所占比例分別為瓶霉屬(Phialophora)43.56%、47.58%、60.16%和糞殼菌綱—未分類43.06%、28.98%、15.89%。原核、真核微生物屬的分布情況見圖6,其中相對豐度低于1%的菌屬合并為其他。
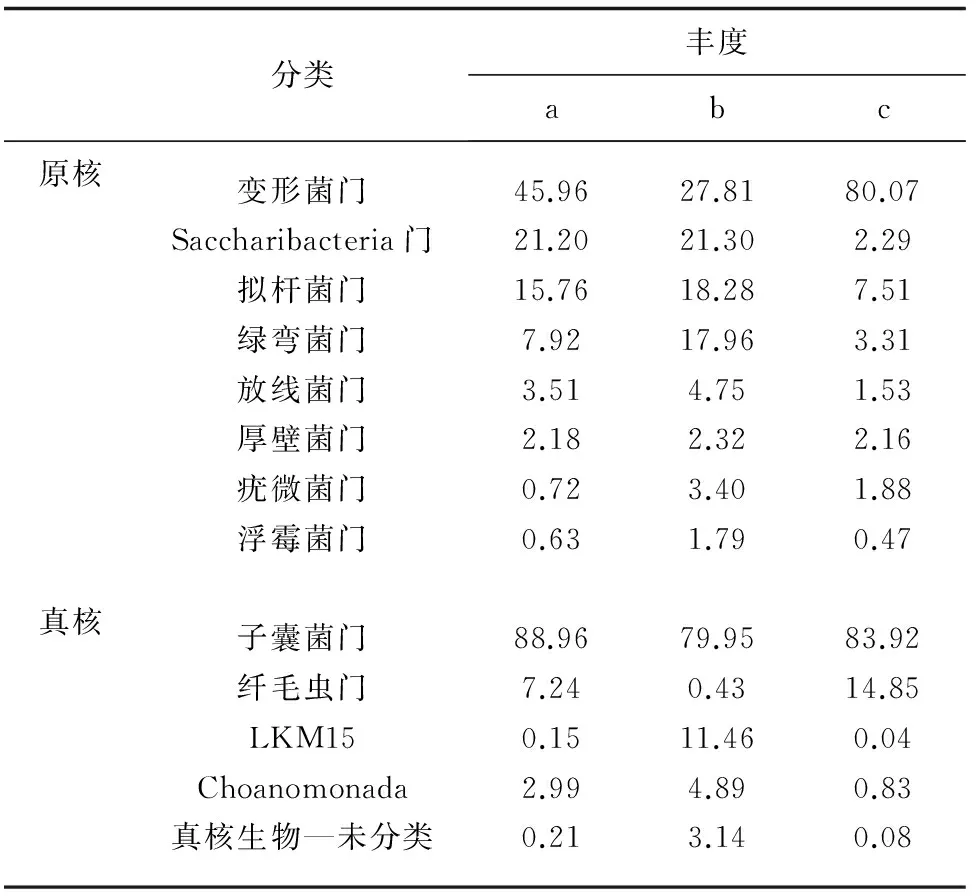
表4 不同堿度組絮體原核和真核微生物各門的豐度 %
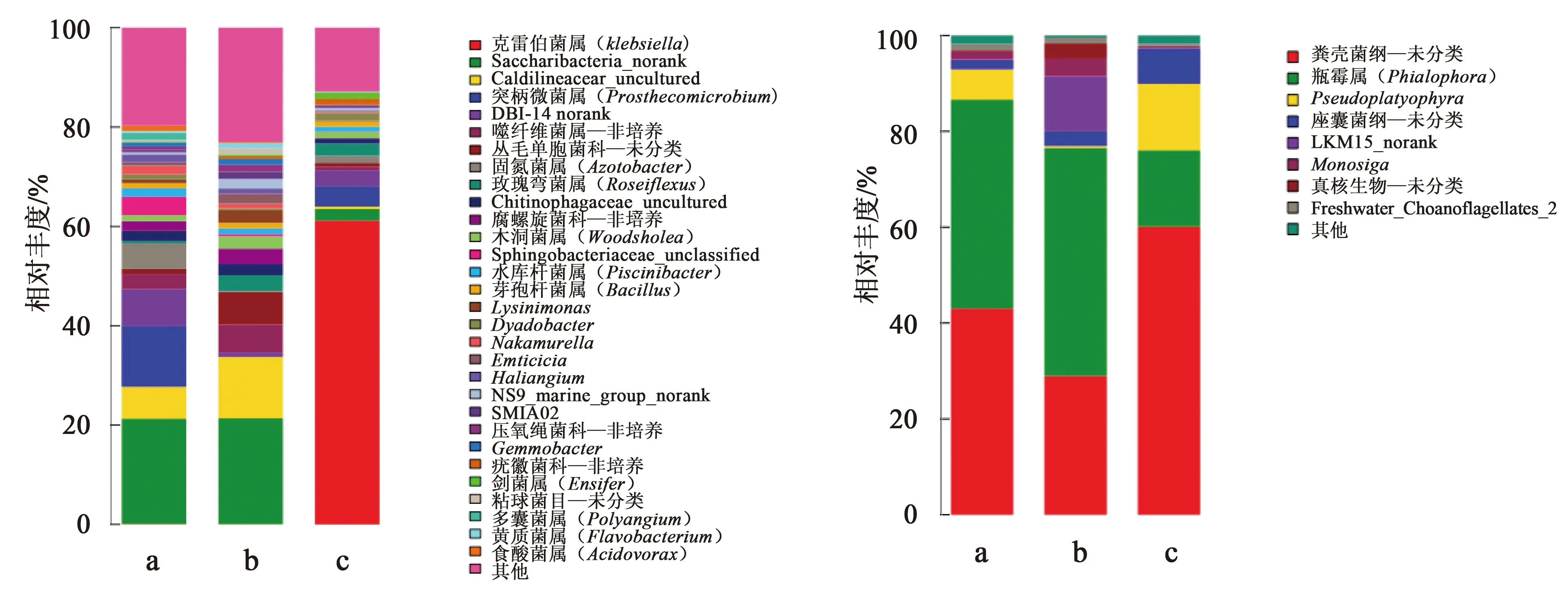
圖6 3組樣品微生物中不同屬的原核16S(左)和真核18S(右)的分布
3 討 論
3.1 堿度對生物絮體培養水質參數的影響
通常認為水產養殖生物絮凝系統中碳氮比、溫度、溶解氧、pH 等決定了水體中微生物分解作用的強弱。本試驗中,反應器中懸浮固體顆粒物含量高,隨著其分解氨氮快速增加,堿度降低,要維持較高堿度,則需要頻繁大量加堿。提高堿度可促進氨化與硝化作用,緩沖水體pH,為微生物快速增長提供了穩定環境。王大鵬等[14]在零換水生物絮凝養殖凡納濱對蝦(Litopenaeusvannamei)時發現,將堿度提高至100 mg/L以上,細菌數量明顯增加,能有效提高水處理效果。當系統啟動階段結束,氨氮含量降低并穩定,細菌將水中氮素轉化為菌體蛋白,隨之系統的堿度也維持穩定狀態。
3.2 堿度對生物絮體生物學特性的影響
生物絮體含有大量細菌、原生動物、藻類等微型生物群體,其中活的生物體占10%~90%[15],而原生動物、輪蟲是生物絮體系統中的捕食者,在活性污泥凈化廢水中被用作“指示生物”,一方面通過捕食生物絮體中的細菌,促使細菌保持生長期,延緩衰老,優化菌群結構,同時捕食水體中不能自由沉降去除的小于10 μm粒徑的懸浮顆粒物質,增強除氮凈水能力。另一方面能分泌黏性代謝物質,吸附水中的懸浮顆粒,促使絮狀凝結[16]。本試驗發現,堿度為75~100 mg/L下纖毛蟲類原生動物增多,形成的絮體較密實,堿度大于150 mg/L下輪蟲數量增多,因此可通過調節堿度來控制理想微型動物的種類和數量,解決生物絮體老化等問題。堿度不同使生物絮凝系統產生了不同的菌群結構,不同微生物分泌的胞外聚合物含量不同,堿度為150~200 mg/L這種較溫和的環境下,微生物分泌較多的胞外聚合物,且蛋白與多糖比較高,有利于絮體形成。絮體粗蛋白和粗脂肪含量達到了羅非魚(Oreochromis)的生長需求。可見循環水高密度養殖鰻鱺固體廢棄物基質含量高,生物絮體資源化可利用度佳。
3.3 堿度對生物絮體原核與真核微生物的影響
本試驗中,水產生物絮體中變形細菌門的數量占主導地位,與夏耘等[5, 17]的結果一致,但不同堿度系統中變形細菌門所占比例有較大差異,尤其是堿度為75~100 mg/L下培養的絮體,變形菌屬在細菌組成上占支配地位,其中克雷伯氏菌屬占 61.19%。該屬具有固氮作用[18],此門細菌是污水處理系統中去除污染物的優勢菌種,說明生物絮體能高效調節水產養殖的水質。Kindaichi等[19]研究了活性污泥中Saccharibacterium門細菌系統發育多樣性和生理機能。結果表明是一個系統發育不同的群體,之前劃分在TM7門,Saccharibacteria在有機化合物的降解中發揮作用,在有氧、缺氧和以硝酸鹽作電子受體條件下能吸收葡萄糖,一些種屬還可以利用N-乙酰葡糖胺、油酸、氨基酸和丁酸,一些絲狀Saccharibacteria表現出β-半乳糖苷酶和脂肪酸的活性,但是Saccharibacteria不吸收乙酸、丙酮酸、丙酸、甘油和乙醇。本試驗以葡糖糖為碳源培養絮體,堿度為150~200 mg/L和大于250 mg/L組Saccharibacteria所占比例分別為21.2%和21.3%,75~100 mg/L組中僅含2.29%, 說明高堿度下有利于Saccharibacteria生長,充分利碳源—葡萄糖,同時堿度大于250 mg/L組和150~200 mg/L組絮體中絲狀菌較多,也是此細菌所占比例較多所致。厚壁菌門以芽孢桿菌(Bacillus)為主,本試驗發現堿度對此門細菌的含量影響差異不顯著。芽孢桿菌分泌的蛋白酶和淀粉酶可以分解污水中的大分子有機物,在特性條件下產生的細菌素對某些致病菌有抑制作用。本試驗中不同堿度條件下均未檢測到弧菌屬(Vibrio)等病原菌。細菌介導參與厭氧氨氧化,使得亞硝酸鹽和銨可以直接轉化成氮氣,因此可以去除廢水處理中的氨氮,執行此過程的細菌均屬于浮霉菌門[20]。本試驗中,堿度為150~200 mg/L組浮霉菌門細菌所占比例最大超過1%,真菌作為生物絮體重要組成部分。大量研究表明,許多淡水子囊菌的子囊孢子具有膠狀的鞘、附著絲,在水里會伸展的很長,成為水體中動物的食物,構成生態系統的食物鏈[21]。本試驗檢測到絮體中超過80%的真菌均屬于子囊菌門,其中分類位置未定的糞殼菌屬(Sordaria)占主導地位,能夠分解纖維素和木質素等固體基質,促進殘餌糞便分解形成生物絮體。
[1] Wasielesky W, Atwood H, Stokes A, et al. Effect of natural production in a zero exchange suspended microbial floc based super-intensive culture system for white shrimpLitopenaeusvannamei[J]. Aquaculture, 2006, 258(1/4):396-403.
[2] Furtado P S, Gaona C A P, Poersch L H, et al. Application of different doses of calcium hydroxide in the farming shrimpLitopenaeusvannameiwith the biofloc technology (BFT)[J]. Aquaculture International, 2014, 22(3):1009-1023.
[3] 阮贇杰. 水產養殖與加工廢水生物絮體資源化技術研究[D].杭州:浙江大學,2013.
[4] Luo G Z, Avnimelech Y, Pan Y F, et al. Inorganic nitrogen dynamics in sequencing batch reactors using biofloc technology to treat aquaculture sludge[J]. Aquacultural Engineering, 2013, 52(1):73-79.
[5] 夏耘, 邱立疆, 郁二蒙,等.生物絮團培養過程中養殖水體水質因子及原核與真核微生物的動態變化[J].中國水產科學, 2014,20(1):75-83.
[6] Chen S, Jian L, Blancheton J P. Nitrification kinetics of biofilm as affected by water quality factors[J]. Aquacultural Engineering, 2006, 34(3):179-197.
[7] Boyd C E, Tucker C S. Pond Aquaculture Water Quality Management [M].US:Springer,1998:106.
[8] Furtado P S, Poersch L H, Wasielesky W.The effect of different alkalinity levels onLitopenaeusvannameireared with biofloc technology (BFT) [J].Aquaculture International, 2015,23(1):345-358.
[9] 陳家捷, 譚洪新, 羅國芝,等.羅非魚糞便在分解過程中形態和營養成分變化[J].水產科學, 2015, 34(10):634-939.
[10] Luo G, Liang W, Tan H, et al. Effects of calcium and magnesium addition on the start-up of sequencing batch reactor using biofloc technology treating solid aquaculture waste[J]. Aquacultural Engineering, 2013, 57(6):32-37.
[11] 龍騰銳, 龍向宇, 唐然,等. 胞外聚合物對生物絮凝影響的研究[J]. 中國給水排水, 2009, 25(7):30-34.
[12] 夏志紅, 任勇翔, 楊壘, 等.自養菌和異養菌胞外聚合物對活性污泥絮凝特性的影響[J].環境科學學報, 2015,35(2):468-475.
[13] 王淑瑩, 何岳蘭, 李夕耀,等.不同活性污泥胞外聚合物提取方法優化[J].北京工業大學學報, 2016,42(4):569-76.
[14] 王大鵬, 何安尤, 韓耀全,等.堿度調節對凡納濱對蝦室內高密度養殖固定化微生物處理效果的影響[J].中國水產科學, 2014,20(2):330-339.
[15] 趙培. 生物絮團技術在海水養殖中的研究與應用[D]. 上海:上海海洋大學,2011.
[16] 沈成媛, 萬小娟. 附著生活型輪蟲的懸浮固體去除特性[J]. 環境工程學報, 2013, 7(12):5047-5050.
[17] 楊章武, 楊鏗, 張哲,等.基于宏基因組測序技術分析凡納濱對蝦育苗中生物絮團細菌群落結構[J].福建水產,2015,37(2):91-97.
[18] Chen W M, Tang Y Q, Mori K, et al. Distribution of culturable endophytic bacteria in aquatic plants and their potential for bioremediation in polluted waters[J]. Aquatic Biology, 2012(15):99-110.
[19] Kindaichi T, Yamaoka S, Uehara R, et al.Phylogenetic diversity and ecophysiology of candidate phylum Saccharibacteria in activated sludge[J].Fems Microbiology Ecology, 2016, 92(6):fiw078.
[20] 田美, 劉漢湖, 申欣,等. 百樂克(BIOLAK)活性污泥宏基因組的生物多樣性及功能分析[J]. 環境科學, 2015,36(5):1739-1748.
[21] 胡殿明. 淡水糞殼綱及無性型真菌系統學及人為干擾下的淡水真菌多樣性研究[D]: 北京:中國林業科學研究院,2011.
EffectofAlkalinityonNitrogenConversionandBiologicalCharacteriztionofBioflcsinBioflocculationProcessinAquaculture
MA Tao1, LUO Guozhi1,2,3, TAN Hongxin1,2,3,CHEN Wei1
( 1. College of Fisheries and Life Science, Shanghai Ocean University, Shanghai 201306, China;2. Shanghai Aquacultural Engeering Research Center, Shanghai 201306, China;3. Aquatic Animal Breeding Center, Shanghai University Knowledge Service Platform,Shanghai 201306, China )
Sodium bicarbonate and 1 mol/L hydrochloric acid (HCl) were regularly added into a reactor containing solid wastes including feed remnants and feces collected from a system of eelAnguillamarmorataculture at various alkalinities (CaCO3/L) of ≥250 in Group a, 150—200 in Group B and 75—100 mg in Group c with three replicates to cultivate bioflocs to evaluate changes in levels of three types of nitrogen and conversion rate of nitrogen by bioflocs, and contents of crude protein, crude fat, and extracelluar polymeric substances (EPS) in the bioflocs. The diversity of microbial communities in the bioflocs was detected by Illumina Miseqsequencing technology. The results showed that there were higher nitrogen conversion rate(70.84±7.67)%, crude protein content (36.74±0.59)% and the ratio of protein and polysaccharide in loosely bound-EPS (LB-EPS) and tightly bound-EPS (TB-EPS) and higher Shannon diversity index in prokaryotic and eukaryotic microbial community in Group b than those in Group a and Group c. The nitrification was promoted in the reactor where the alkalinity was increased to above 150 mg/L (CaCO3).ProteobacteriaandSaccharibacteriawere of the dominant bacteria with significant differences in prokaryotic community in flocs of the three groups,accounting for 45.96% in Group a, 27.81% in Group b, and 80.07% in Group c. In eukaryotic community, however,AscomycotaandCiliophorawere the dominant bacteria with significant differences,Ciliophorarepresenting 7.24% in Group a, 0.43% in Group b, and 14.85% in Group c. In conclusion,the high microbial diversity, and efficiency of nitrogen conversion in bioflocculation are found in the reactor at alkalinity of over 150 mg/L (CaCO3).
alkalinity; feces; biofloc; Illumina Miseq; microbial community
10.16378/j.cnki.1003-1111.2017.04.004
S912
A
1003-1111(2017)04-0421-08
2016-07-08;
2016-09-12.
國家自然科學基金資助項目(31202033);上海市科技委員會資助項目 (14320501900).
馬濤(1990-),女,碩士研究生;研究方向:水產養殖重復利用. E-mail:mt1011207@163.com.通訊作者:羅國芝(1974-),女,副教授,博士;研究方向:水產養殖重復利用和循環水養殖系統與工程.E-mail: gzhluo@shou.edu.cn.
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
