草地早熟禾干旱脅迫轉錄組差異性分析
2017-12-22 01:52:58冷暖劉曉巍張娜許立新
草業學報 2017年12期
冷暖,劉曉巍,張娜,許立新*
(1.北京林業大學草坪研究所, 北京 100083;2.南京市老山林場, 江蘇 南京 211811)
草地早熟禾干旱脅迫轉錄組差異性分析
冷暖1,劉曉巍2,張娜1,許立新1*
(1.北京林業大學草坪研究所, 北京 100083;2.南京市老山林場, 江蘇 南京 211811)
干旱是影響草地早熟禾生產力的主要因素之一。在沒有參考基因組的情況下, 為了揭示草地早熟禾在干旱處理下基因表達譜的變化, 本文采用了高通量Illumina Hiseq測序平臺對草地早熟禾的對照組(CK)與干旱處理組(D)進行轉錄組測序, 并對測序數據進行了分析研究, 進一步探究了草地早熟禾干旱應答的分子機制。結果表明, 在干旱處理下共檢測到24465個差異表達的基因, 篩選后獲得4143個上調基因和4415個下調基因, 共占差異表達基因總數的34.98%。經富集分析后得, 與蛋白激酶、蛋白磷酸酶、碳代謝以及ABA等相關的基因可以作為研究草地早熟禾干旱響應機制的主要研究對象。qRT-PCR分析表明, 隨機選出的8個差異性表達基因的表達趨勢與高通量測序結果相一致。此外, 還候選了吲哚-3-甘油磷酸合成酶、蛋白磷酸酶、已糖激酶、鈣結合蛋白、葉綠素a/b結合蛋白等基因作為與草地早熟禾干旱脅迫相關的應答候選基因, 為揭示草地早熟禾耐旱分子機制奠定了基礎。
草地早熟禾;干旱脅迫;高通量測序;轉錄組
在我國北方, 草地早熟禾是一種適應性很強的冷季型草, 品質好, 繁殖快, 廣泛應用于公園、居住區、運動場等地綠化。但我國北方大部分地區為干旱半干旱地區, 不同地區面臨著不同程度的缺水問題, 所以干旱成為影響草地早熟禾在北方生長發育的主要因素之一。水分是植物體的重要組成部分, 它參與植物的光合、呼吸、蒸騰等生理作用, 也是影響植物形態結構等重要生物因子, 充足的水分供應是植物正常生存的重要條件。干旱脅迫時, 植物為了適應環境, 不僅在生理上會發生反應, 分子上也會產生一定的變化, 一些功能不同的基因在干旱脅迫時表達量會發生上調或者下調。李偉[1]發現, 草地早熟禾(Poapratensis)PpNAC與水稻(Oryzasativa)的NAC蛋白SNACI高度同源, 水稻的SNACI可使植株抗旱性提高, 因而推測PpNAC與草地早熟禾的抗旱性有密切聯系;Xu[2]發現, 干旱會誘導草地早熟禾兩個品種‘Midnight’和‘Brilliant’的Cyt Cu/ZnSOD、Chl Cu/ZnSOD和APX三個基因上調表達, MR基因下調表達, 且與‘Brilliant’相比, 抗旱性好的‘Midnight’品種這4個基因差異表達更顯著;信金娜[3]通過基因槍轟擊法把抗旱、耐鹽有關外源基因(DREB1A、BADH-CMO、CMO)轉入草地早熟禾, 獲得轉基因植株, 結果表明, 轉基因植株在抗旱性方面強弱程度為轉雙基因植株>轉DREB1A、CMO基因植株>對照。抗旱性是一個多基因控制的數量性狀[4], 因而有很多優良基因還沒有能夠完全被準確探明。
Illumina測序屬于二代測序技術, 它是基于特定限制酶切位點的測序分析, 不需要知道基因組序列, 適于研究基因組不明確的物種的基因表達[5], 并且具有測序通量大、成本低的優勢。目前, 在黑麥草(Loliumspp.)[6]、紫花苜蓿(Medicagosativa)[7]、結縷草(Zoysiajaponica)[8-9]以及草地早熟禾[10]的研究中都有應用, 通過測序技術獲得草坪草不同脅迫環境的響應基因并進行基因表達差異性分析。因此, Illumina測序技術是研究草地早熟禾干旱脅迫基因差異表達較好的選擇。
本研究中運用Illumina高通量測序平臺的轉錄組測序技術, 分別對正常水分條件和干旱條件下生長的草地早熟禾成熟葉片的轉錄產物mRNA進行測序,測得的數據能夠較全面反映草地早熟禾干旱脅迫下基因表達狀況, 分別對差異表達基因進行GO、KEGG(Kyoto Encyclopedia of Genes and Genomes)注釋和富集分析, 進一步篩選出與草地早熟禾抗旱相關的基因, 為之后草地早熟禾SSR標記開發、抗旱性相關基因的發掘及分子育種提供借鑒。
1 材料與方法
1.1 實驗材料與處理
以草地早熟禾的‘Nuglade’品種為材料, 購自北京綠冠公司。草坪養護在北京林業大學八家村實驗站溫室中進行。試驗于2016年1月開始,設置兩種處理, 對照處理和干旱脅迫處理。對照處理的植株2 d澆一次水, 保證土壤含水量在35%;干旱脅迫處理組不澆水, 直至土壤水分降到4%。每個處理3次生物學重復。以正常澆水的材料為對照樣本(CK), 干旱脅迫為處理樣本(D)。在干旱脅迫后第15天的8:30-9:00之間對對照組(CK)和干旱處理組(D)進行取樣。
1.2 文庫的構建及庫檢
按照TRIzol植物RNA提取試劑盒方法分別提取草地早熟禾對照組(CK)與干旱處理組(D)樣品葉片的RNA。所提取的RNA經電泳檢測合格后送至北京諾禾致源生物有限公司進一步確認質量, 合格后利用 Oligo(dT)磁珠富集真核生物mRNA, 并以 Oligo (dT)引導反轉錄合成雙鏈 cDNA, 通過PCR擴增和AMPure XP beads純化后獲得最終文庫。文庫構建完成后, 使用Qubit2.0進行初步定量, 用Agilent 2100對文庫進行檢測, 符合預期后, 使用Q-PCR方法對文庫的有效濃度進行準確定量(文庫有效濃度>2 nmol/L), 以保證文庫質量。庫檢合格后, 進行Illumina HiSeq測序。
1.3 Illumina測序及測序數據的分析
1.3.1測序原始數據質量評估與轉錄本拼接 高通量測序(如Illumina HiSeqTM)得到的原始圖像數據文件經CASAVA堿基識別(base calling)分析轉化為原始測序序列(sequenced reads), 稱之為 Raw Data或Raw Reads。通過測序錯誤率分布檢查, A/T/G/C含量分布檢查, 進行c測序數據過濾后得到Clean Reads。對于無參考基因組的項目, 獲得Clean Reads后, 需要對Clean Reads進行拼接以獲取后續分析的參考序列。采用Trinity對Clean Reads進行拼接[11]。將Trinity拼接得到的轉錄本序列, 作為后續分析的參考序列。取每條基因中最長的轉錄本作為Unigene, 以此進行后續的分析。
1.3.2基因功能注釋 為獲得全面的基因功能信息, 進行了七大數據庫的基因功能注釋, 包括: Nr, Nt, PFAM, KOG/COG, Swiss Prot, KEGG, GO。得出數據在七大數據庫中的注釋成功率情況。
1.3.3差異表達基因分析 統計好樣本中基因表達量后, 分別對對照組(CK)與干旱處理組(D)之間基因表達情況進行對比, 找出表達量存在差異的基因, 并通過篩選進一步發掘出顯著差異表達的基因。對于有生物學重復的樣品, 采用DESeq[12]進行分析, 篩選閾值為p-adjusted<0.05。
1.3.4差異表達基因富集分析 GO功能顯著性富集分析給出與基因組背景相比, 在差異表達基因中顯著富集的GO功能條目, 從而給出差異表達基因與哪些生物學功能顯著相關。GO富集分析方法為 GOseq[13],此方法基于Wallenius non-central hyper-geometric distribution。在生物體內, 不同基因相互協調行使其生物學功能, 通過Pathway顯著性富集能確定差異表達基因參與的最主要生化代謝途徑和信號轉導途徑。KEGG是有關Pathway的主要公共數據庫[14]。
1.4 qRT-PCR轉錄組數據驗證
隨機選取8個差異性表達的基因進行qRT-PCR分析, 對高通量數據進行驗證。分別提取對照組(CK)與干旱組(D)的葉片RNA, 采用HiScript? Ⅱ Q RT SuperMix for qPCR試劑盒反轉錄合成cDNA, 設計引物(表1)進行qRT-PCR實驗, Actin基因作為內參。各個處理均做3次重復, 計算基因的相對表達量。

表1 干旱脅迫條件下草地早熟禾8個差異性表達基因qRT-PCR引物Table 1 Primers of eight differentially expressed genes used for quantitative real-time PCR in P. pratensis at drought stress
2 結果與分析
2.1 測序數據統計與評估
測序得到的原始測序序列Raw Reads中含有帶接頭的、低質量的Reads, 對照及各處理組的Raw Reads都在4052萬條以上。在去除帶接頭的Reads、去除N的比例大于0.1%的Reads、去除低質量Reads (質量值Qphred≤20的堿基數占整個Reads的50%以上的Reads) 后, 得到Clean Reads, 后續分析都基于Clean Reads。經過濾后, 共得到59.35G的有效Reads (Clean Reads)。由表2可知, 各組得到的Clean Reads的量占原始Reads的比例都達到了96%以上, 說明測序的建庫工作質量良好。
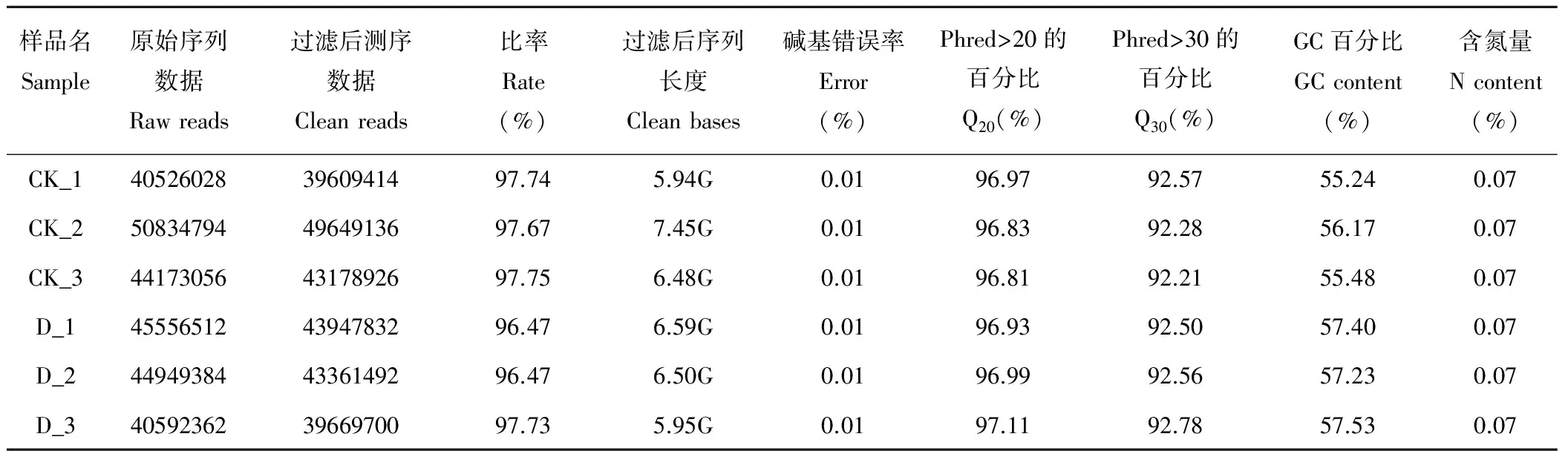
表2 測序數據輸出質量情況Table 2 Sequencing data quality evaluation
2.2 轉錄組的拼接
拼接所得的序列長度用于衡量拼接質量, 如拼接所得的序列長度越長, 則測序質量越好。經Trinity拼接后, 獲得轉錄本(Transcript)435250條, 總長度為324271434 nt,平均長度為745 nt, N50長度為1163 bp;Unigene 254331條, 總長度為147657025 nt,平均長度為581 nt, N50長度為818 bp。具體的組裝統計結果見表3。由表3數據可知, 拼接所得片段有很高的組裝完整性, 拼接所得序列長, 測序質量好。
2.3 基因功能注釋
為了預測Unigene的功能, 對其進行基因注釋, 具體方法為將Unigene序列分別與主要的生物學數據庫, 如GO、KEGG、NR等進行基因信息比對, 最終有254331條Unigene獲得注釋, 其中, NR數據庫注釋的信息最多, 占全部Unigene的30.58%。7842條Unigene同時比對到7個數據庫中, 占總數的3.08%。105852條Unigene至少在一個數據庫中得到功能注釋, 占到總數的41.61%。結果如圖1所示。
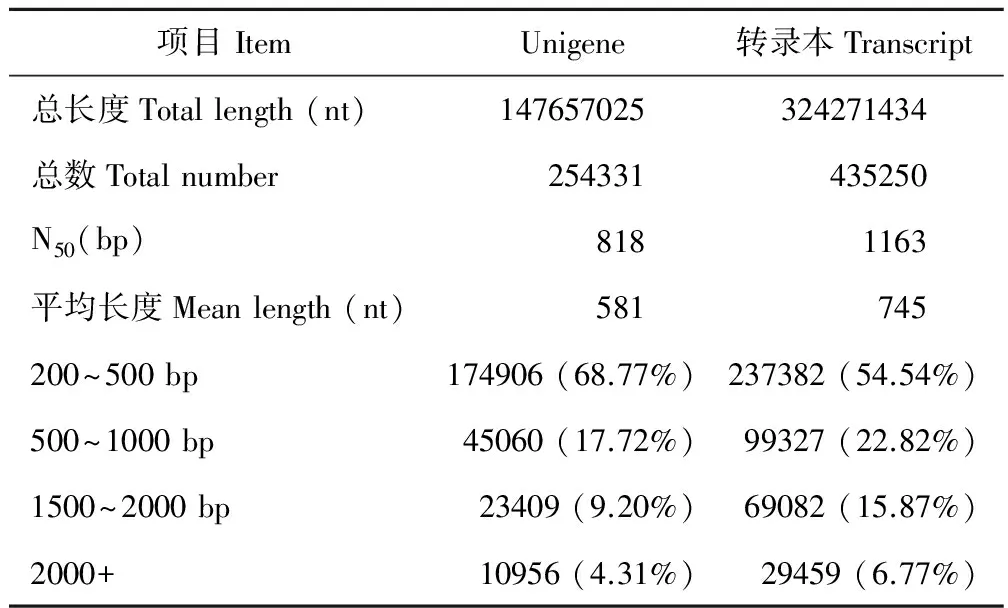
表3 Unigene與轉錄本的長度統計結果表Table 3 Unigene and transcript length statistics
N50: 將組裝片段按從長到短排序, 并進行長度值累加, 當長度值累計至總長度的50%時, 最后累加的片段長度值。Unigene: 取每條基因中最長的轉錄本作為該基因的代表, 稱為Unigene, 以此進行后續的分析。nt: 核苷酸單位。N50: The assembled segments are sorted from long to short and the length values are accumulated. When the length value is accumulated to 50% of the total length. The last accumulated fragment length value. Unigene: The longest transcript of each gene was used as a representative of the gene, called Unigene, followed by subsequent analysis. nt: The unit of nucleotides.
2.4 基因差異性表達分析
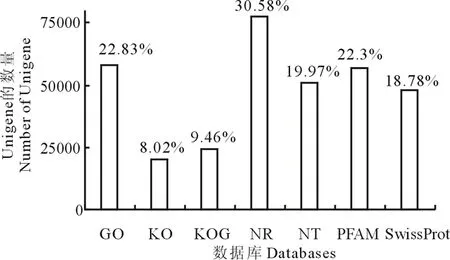
圖1 Unigene在各大數據庫中的功能注釋分布柱狀圖Fig.1 Distribution histogram of Unigene functional annotation according to the major databases
對對照組(CK)與干旱組(D)進行差異基因篩選(DEGs), 經篩選, 共鑒定出24465個差異表達的基因, 其中上調基因有12783個, 占差異表達基因總數的52.25%;下調基因共11682個, 占差異表達基因總數的47.75%。結果如圖2所示。
2.5 差異表達基因的GO分析
GO 分析(gene ontology)是一個常用的基因功能分類體系, 它按照生物途徑(biology process), 分子功能(molecular function)和細胞組分(cellular component)對基因進行注釋和分類。通過對差異表達基因進行GO terms富集度統計學的分析, 計算出差異基因GO term的p-value和p-value的FDR值(q-value), 定位差異基因最可能相關的GO term。GO 分析對實驗結果有提示的作用, 通過差異基因的GO 分析, 可以找到富集差異基因的GO分類條目, 尋找不同樣品的差異基因可能和哪些基因功能的改變有關。
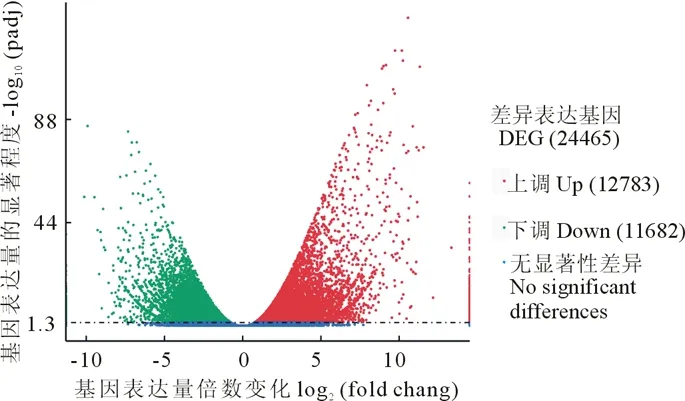
圖2 干旱處理(D)/對照組(CK)基因差異表達分析火山圖Fig.2 Volcanic map of differential gene expression analysis between drought treatment (D)/control group (CK)
對干旱組(D)與對照組(CK)之間的差異表達基因(DEGs)進行GO功能分析得到圖3, 經分析, 代謝過程(metabolic process)、蛋白磷酸化(protein phosphorylation)、泛素連接酶復合物(ubiquitin ligase complex)、催化活性(catalytic activity)和蛋白激酶活性(protein kinase activity)等與干旱脅迫緊密相關的生物學途徑、分子功能、細胞組成等都有明顯的響應變化。
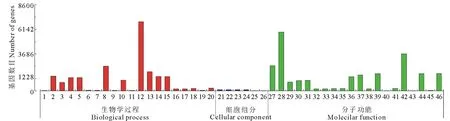
圖3 差異表達基因GO富集分類Fig.3 GO enrichment classification of differentially expressed genes 1: 響應水 Response to water;2: 大分子修飾 Macromolecule modification;3: 蛋白磷酸化 Protein phosphorylation;4: 細胞蛋白修飾過程 Cellular protein modification proess;5: 蛋白修飾過程 Protein modification process;6: 響應非生物刺激 Response to abiotic stimulus;7: 響應無機物質 Response to inorganic substance;8: 蛋白代謝過程 Protein metabolic process;9: 響應含氧化合物 Response to oxygen-containing compound;10: 磷酸化 Phosphorylation;11: 響應酸性化學品 Response to acid chemical;12: 代謝過程 Metabolic process;13: 細胞蛋白質代謝過程 Cellular protein metabolic process;14: 磷酸代謝過程 Phosphorus metabolic process;15: 含磷酸鹽的化合物代謝 Phosphate-containing compound metabolic;16: 蛋白泛素化 Protein ubiquitination;17: 小蛋白質共軛修飾 Protein modification by small protein conjugation;18: 小蛋白質共軛或去除修飾 Protein modification by small protein conjugation or removal;19: 過期過氧化酶反應 Obsolete peroxidase reaction;20: 脂質生物合成過程 Lipid biosynthesis process;21: 泛素連接酶復合物 Ubiquitin ligase complex;22: Cullin-RING泛素連接酶復合物 Cullin-RING ubiquitin ligase complex;23: 核泛素連接酶復合物 Nuclear ubiquitin ligase complex;24: 后期促進復合物 Anaphase-promoting complex;25: 脂肪酸合酶復合物 Fatty acid synthase complex;26: 光系統Ⅱ氧氣演變復合體 Photosystem Ⅱ oxygen evolving complex;27: 轉移酶活性 Transferase activity;28: 催化活性 Catalytic activity;29: 蛋白激酶活性 Protein kinase activity;30: 磷酸轉移酶活性, 酒精組作為受體 Phosphotransferase activity, alcohol group as acceptor;31: 激酶活性 Kinase activity;32: 泛素蛋白轉移酶活性 Ubiquitin-protein transferase activity;33: 泛素類似蛋白轉移酶活性 Ubiquitin-like protein transferase activity;34: 血紅素結合 Heme binding;35: 四吡咯結合 Tetrapyrrole binding;36: 轉移酶活性, 轉移含磷基因 Transferase activity, transferring phosphorus-containing groups;37: ATP結合 ATP binding;38: 轉移酶活性, 轉移除氨基酰基以外的酰基 Transferase activity, transferring acyl groups other than amino-acyl groups;39: 嘌呤核糖核苷三磷酸結合 Purine ribonucleoside triphosphate binding;40: 脂肪酸合酶活性 Fatty acid synthase activity;41: 轉移酶活性, 轉移己糖基 Transferase activity, transferring hexosyl groups;42: 離子結合 Ion binding;43: 3-氧代酰基-[酰基-載體-蛋白]合酶活性 3-oxoacyl-[acyl-carrier-protein] synthase activity;44: 腺苷酸核糖核酸結合 Adenyl ribonucleotide binding;45: 過氧化物酶活性 Peroxidase activity;46: 腺苷酸核苷酸結合 Adenyl nucleotide binding.
其中, 4143個上調Unigenes, 有2416個(58%)Unigenes注釋到生物學過程(BP), 其中富集較顯著的分布在蛋白質改性過程(protein modification process)與單生物體運輸(single-organism transport);543個(13%)Unigenes注釋到細胞組分(CC), 其中富集較顯著的為nucleus;1181個(29%)Unigenes注釋到分子功能(MF), 其中富集較顯著的為轉移酶活性(transferase activity)和陽離子活性(cation activity)。
4415個下調Unigene, 有2643個(60%)Unigenes注釋到生物過程(BP), 其中富集最顯著的為代謝過程(metabolic process)和大分子代謝過程(macromolecule metabolic process);550個(12%)Unigenes注釋到細胞組分(CC), 1217個(28%)Unigenes注釋到分子功能(MF), 均勻分布在20類基因功能中。
2.6 差異表達基因KEGG富集分析
KEGG是系統分析基因功能、基因組信息的數據庫, 可用于進行生物體內代謝分析與代謝網絡研究。為對樣本進行全面的代謝通路分析, 將測序所得Unigene序列比對到KEGG數據庫。不同數據庫注釋基因數目的統計結果顯示, 共有20398條Unigenes在KEGG數據庫中得到注釋, 占總數的8.02%, 參與到129個代謝途徑中, 其中注釋到植物-病原體相互作用(plant-pathogen interaction)中的最多, 為848個。
對樣本對照組(CK)與干旱組(D)之間的差異表達基因(DEGs)進行KEGG代謝通路分析后, 共獲得差異表達基因(DEGs)8764個, 對應在307個代謝通路中。
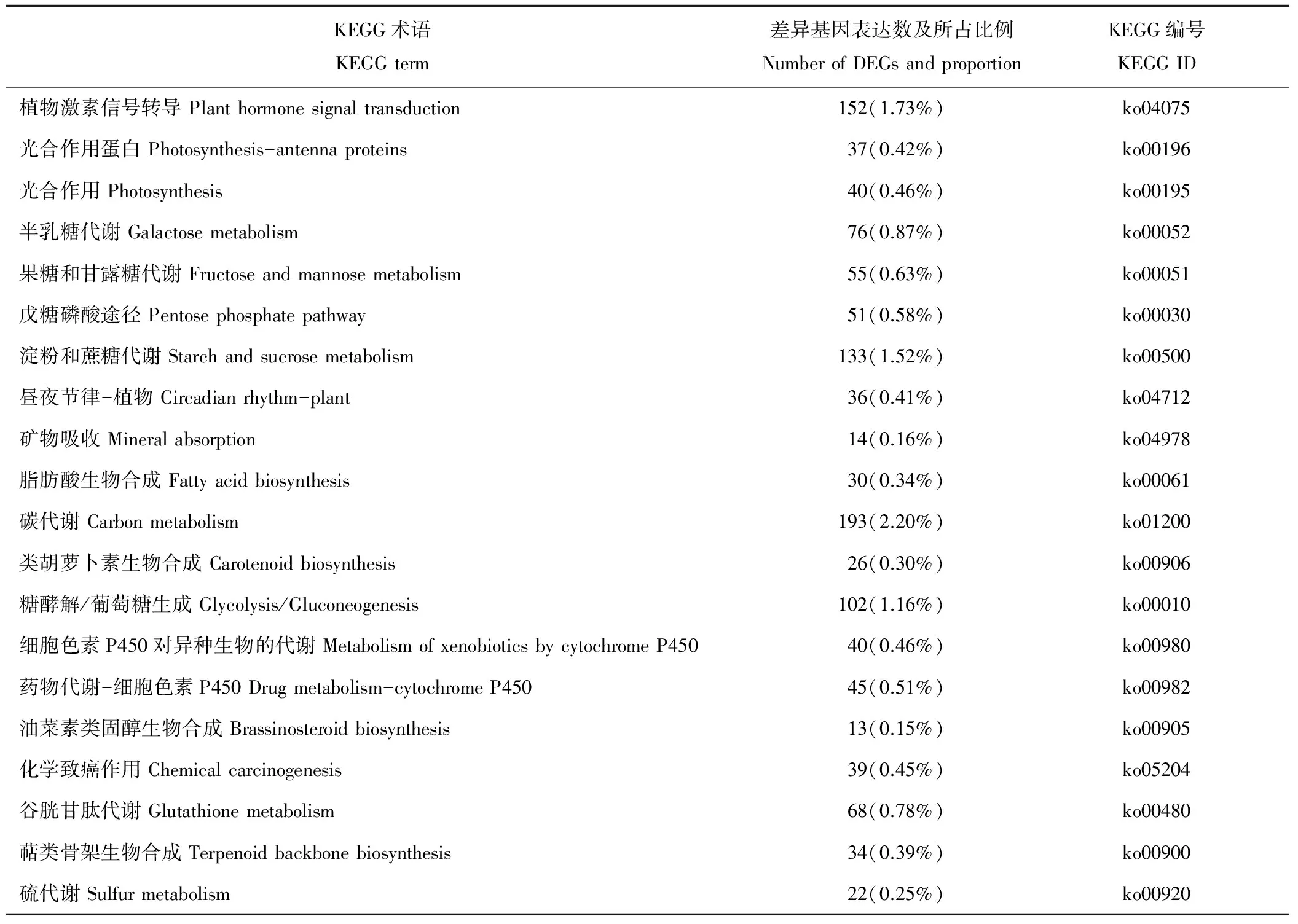
表4 草地早熟禾干旱脅迫下差異基因富集程度排名前20的Pathway條目Table 4 Top 20 enrichment pathway in responses to drought in Poa pratensis
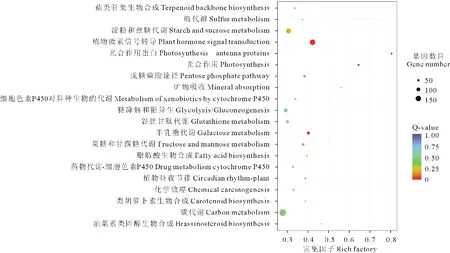
圖4 差異表達基因KEGG富集分析Fig.4 KEGG enrichment analysis of differentially expressed genes
進一步統計后發現, 在307個KEGG Pathway中最顯著的4個Pathway的是植物激素信號轉導(plant hormone signal transduction)、光合作用蛋白(photosynthesis-antenna proteins)、光合作用(photosynthesis)以及半乳糖代謝(galactose metabolism)。 由表4和圖4可知, 排名前20顯著性富集差異基因占差異表達基因總量的13.77%, 主要涉及代謝途徑、信號轉導以及次生代謝等。其中, 與碳代謝相關的基因占差異表達基因總量的2.20%, 與植物激素信號轉導相關的基因占1.73%, 淀粉和蔗糖代謝相關的基因占1.52%, 糖酵解相關的基因占1.16%, 其他相關基因所占比例相對較低。差異性表達基因在植物激素信號轉導和碳代謝的通道上富集程度較高。
從植物激素信號轉導(plant hormone signal transduction)、光合作用蛋白(photosynthesis-antenna proteins)、果糖和甘露糖代謝(fructose and mannose metabolism)等Pathway中候選出部分與吲哚-3-甘油磷酸合成酶、蛋白磷酸酶、己糖激酶、鈣結合蛋白、葉綠素a/b結合蛋白有關的基因作為草地早熟禾干旱響應的候選基因。主要有c120656_g1、c122301_g3、c145664_g1、c117236_g1 、c119413_g2、c137695_g1、c123772_g1、c135567_g1等。
2.7 差異表達基因的qPT-PCR熒光定量驗證
隨機選取8個差異表達基因, 其中含有4個上調表達的差異基因(c111268_g1、c145664_g1、c145507_g1、c128115_g1)和4個下調表達的差異基因(c117236_g1、c119413_g2、c135104_g1、c93924_g1), 以Actin為內參進行qRT-PCR驗證。由圖5可知, 8個基因在干旱脅迫下的表達程度不同, 但表達趨勢與高通量測序結果基本一致, 即表明測序結果真實可靠。

圖5 差異表達基因的qRT-PCR驗證Fig.5 Validation of DEGs data by qRT-PCR
3 討論
植物在干旱脅迫下, 細胞感受干旱脅迫信號, 通過信號轉導, 調控相關基因表達和生理反應, 同時從轉錄和翻譯等不同水平做出響應。目前, 草地早熟禾尚未完成基因組測序, 沒有完整的基因組序列可供參考, 因而不依賴基因組序列信息的高通量測序成為研究草地早熟禾干旱響應基因差異表達更好的選擇。通過數據的統計評估可知數據質量良好。
在GO功能分析中, 生物學過程的差異表達基因差異在蛋白磷酸化、蛋白質修飾過程、磷代謝過程等;細胞組分顯著差異在泛素連接酶復合物、后期促進復合物、細胞膜等;分子功能顯著差異在轉移酶、蛋白激酶、過氧化物酶、脂肪酸合酶等。這說明草地早熟禾干旱脅迫響應是一個復雜的生理變化過程, 要提高其抗旱性也不是單個基因可以完成的。許多研究表明, Ca2+參與細胞內信號轉導過程。當植物受到脅迫時, 細胞質Ca2+濃度瞬時增加, 細胞質Ca2+可以與各種鈣結合蛋白相互作用, 發揮相應的生物學功能。目前, 人們在植物中鑒定出鈣調素、CDPKs(蛋白激酶)和CBLs(鈣調磷酸酶類蛋白)這三類Ca2+感受體參與了脅迫信號轉導[15-16]。有研究顯示, 玉米絲氨酸/蘇氨酸蛋白磷酸酶2C基因ZmPP2Ca與玉米對干旱脅迫的應答有關[17];紫花苜蓿促分裂原活化蛋白激酶MAPK基因與抗旱相關, 其中MMK4可能作為信號傳遞體參與磷酸化與去磷酸化的過程, 來調控紫花苜蓿對干旱的適應性反應[18];水稻OsDISI基因通過在轉錄水平上抑制一系列干旱正調控因子和誘導一系列干旱負調控因子的表達而負調控水稻的干旱脅迫響應過程[19]。本研究發現草地早熟禾干旱脅迫中蛋白激酶活性(GO: 0004672)上調基因394個與蛋白磷酸酶活性(GO: 0004721)上調基因14個都參與了草地早熟禾對干旱脅迫的響應。
在干旱脅迫下, 植物會產生內源激素來提高植物水分的利用效率。在這些激素中, 以ABA的研究最為廣泛。ABA作為信號分子, 通過活化保衛細胞質膜上的PLC和Ca2+離子內流通道, 引起保衛細胞細胞質內Ca2+濃度的升高, 并激活K+外流通道, 最終使保衛細胞水勢升高而失水, 引起氣孔關閉, 抑制蒸騰作用, 從而增強植物對干旱的防御性反應。研究發現, 在植物體中許多基因的表達受干旱脅迫調控, 其中相當一部分基因響應ABA[20]。另外, 在對野生小麥根干旱響應蛋白的篩選與鑒定中發現, 表達差異蛋白主要涉及碳代謝等[21]。本研究中, 通過對草地早熟禾差異表達基因進行KEGG富集分析發現, 在前20顯著性富集差異中, 與碳代謝(ko01200)、植物激素信號轉導(ko04075)相關的基因相對較多。
在對擬南芥的耐受脅迫的研究中發現, 由茉莉酮酸介導的代謝途徑的協調激活提供了對環境脅迫的抵抗力, 這些代謝途徑中包含有吲哚-3-甘油磷酸合成酶的基因[22]。大麥穗干旱脅迫下發現, 它的芒中吲哚-3-甘油磷酸合成酶和葉綠素a/b結合蛋白基因下調表達[23]。在擬南芥中, 葉綠素a/b結合蛋白(LHCB)基因的下調表達會降低氣孔運動對ABA的反應度, 從而導致擬南芥在干旱脅迫下的耐受性降低, 反之則增強[24]。在黑麥草的干旱脅迫研究中發現, 碳代謝相關基因有助于提高耐旱性, 其中己糖激酶(HxK)編碼基因在抗性系中顯示上調[25]。
使用qRT-PCR對隨機選出的8個基因的表達量檢測結果與高通量測序的表達結果總體趨勢吻合, 但在具體表達倍數的數值上存在差異, 這可能與樣本送測時間, 數據分析方法及檢測靈敏度不同有關。
4 結論
本研究以草地早熟禾‘Nuglade’為材料, 以正常水分和干旱脅迫條件下的2個樣本葉片進行轉錄組分析, 在轉錄水平上獲得254331個基因的真實序列信息。檢出17110個SSR位點, 通過差異基因篩選, 共鑒定出24465個差異表達基因。其中有12783個DGEs在干旱脅迫響應中表達上調, 11682個表達下調。分析得出, 與蛋白激酶、蛋白磷酸酶、碳代謝以及ABA等相關的基因可以作為研究草地早熟禾干旱響應機制的主要研究對象, 吲哚-3-甘油磷酸合成酶、蛋白磷酸酶、己糖激酶、鈣結合蛋白、葉綠素a/b結合蛋白的基因或轉錄因子可作為與草地早熟禾干旱脅迫相關的應答候選基因, 從而揭示草地早熟禾葉片響應干旱脅迫的分子機制。
References:
[1] Li W. Cloning and Expression Analysis of Transcription Factors GenePpNACinPoapratensisL. Beijing: Chinese Academy of Forestry, 2011.
李偉. 草地早熟禾轉錄因子基因PpNAC的克隆和表達分析. 北京: 中國林業科學研究院, 2011.
[2] Xu L X, Han L B, Huang B R. Antioxidant enzyme qctivities and gene expression patterns in leaves of kentucky bluegrass in response to drought and post-drought recovery. Journal of The American Society for Horticultural Science, 2011, 136(4): 247-255.
[3] Xin J N. Genetic Transformation of Drought-resistance and Salt-resistance Gene in Kentucky Bluegrass (PoapratensisL.). Beijing: Beijing Forestry University, 2006.
信金娜. 草地早熟禾(PoapratensisL.)抗旱耐鹽基因遺傳轉化. 北京: 北京林業大學, 2006.
[4] Zhang Z B, Shan L. Advances in inheritance of crop drought resistance physiological traits. Chinese Science Bulletin, 1998, (17): 1812-1817.
張正斌, 山侖. 作物抗旱生理性狀遺傳研究進展. 科學通報, 1998, (17): 1812-1817.
[5] Wang X C, Yang Z R, Wang M,etal. High-throughput sequencing technology and its application. China Biotechnology, 2012, (1): 109-114.
王興春, 楊致榮, 王敏, 等. 高通量測序技術及其應用. 中國生物工程雜志, 2012, (1): 109-114.
[6] Duhoux A, Carrere S, Gouzy J,etal. RNA-Seq analysis of rye-grass transcriptomic response to an herbicide inhibiting acetolactate-synthase identifies transcripts linked to non-target-site-based resistance. Plant Molecular Biology, 2015, 87(4/5): 473-487.
[7] Zhou Q, Luo D, Ma L C,etal. Development and cross-species transferability of EST-SSR markers in siberian wildrye (ElymussibiricusL.) using illumina sequencing. Scientific Reports, 2016, 6: 20549.
[8] Zhu C, Ai L, Wang L,etal. De Novo Transcriptome Analysis of Rhizoctonia Solani AG1 IA Strain Early Invasion inZoysiajaponicaRoot. Frontiers in Microbiology, 2016, 7: 708.
[9] Ahn J H, Kim J, Kim S,etal. De novo transcriptome analysis to identify anthocyanin biosynthesis genes responsible for tissue-specific pigmentation inZoysiagrass (ZoysiajaponicaSteud.). Plos One, 2015, 10: e01379439.
[10] Zhao S, Zhang N, Liu D Y,etal. The impact of fenarimol application on the magnaporthe poae and fungi in the rhizosphere soil of Kentucky Bluegrass (PoapratensisL.). Journal of Nanjing Agricultural University, 2015, (4): 590-595.
趙爽, 張寧, 劉東陽, 等. 氯苯嘧啶醇施用對草坪斑枯病致病菌及根際土壤真菌的影響. 南京農業大學學報, 2015, (4): 590-595.
[11] Grabherr M G, Haas B J, Yassour M,etal. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nature Biotechnology, 2011, 29(7): 130-644.
[12] Anders S, Wolfgang H. Differential expression analysis for sequence count data. Genome Biology, 2010, 11(10): 106.
[13] Young M D, Wakefield M J, Smyth G K,etal. Gene ontology analysis for RNA-seq: accounting for selection bias. Genome Biology, 2010, 11: 142.
[14] Kanehisa M, Araki M, Goto S,etal. KEGG for linking genomes to life and the environment. Nucleic Acids Research, 2008, 36(SI): 480-484.
[15] Zhang J J, Li J J, Nian H J. The role of calcium/calmodulin signaling pathways in the stresses: progress in researches. Chinese Journal of Microecology, 2013, (7): 858-860.
張晶晶, 李金金, 年洪娟. 鈣/鈣調素信號途徑在脅迫中的作用研究進展. 中國微生態學雜志, 2013, (7): 858-860.
[16] Sanchez-Barrena M J, Fujii H, Angulo I,etal. The structure of the C-terminal domain of the protein kinase AtSOS2 bound to the calcium sensor AtSOS3. Molecular Cell, 2007, 26(3): 427-435.
[17] He L., Li F H, Sha L N,etal. Activity of serine/threonine protein phosphatase type-2C (PP2C) and its relationships to drought tolerance in maize. Acta Agronomica Sinica, 2008, (5): 899-903.
何亮, 李富華, 沙莉娜, 等. 玉米2C型絲氨酸/蘇氨酸蛋白磷酸酶(PP2C)活性與耐旱性的關系. 作物學報, 2008, (5): 899-903.
[18] Bai X M. Cloning of MAPK gene which was activated by drought inMedicagosativa. Inner Mongolia Petrochemical Industry, 2011, (5): 18-20.
白雪梅. 紫花苜蓿(Medicagosativa)抗旱相關促分裂原活化蛋白激酶MAPK基因的克隆. 內蒙古石油化工, 2011, (5): 18-20.
[19] Ning Y S. Functional and Mechanistic Analysis of the SINA E3 Ligase OsDIS1 in Rice. Changsha: Hunan Agricultural University, 2011.
寧約瑟. 水稻SINA泛素連接酶OsDIS1的功能分析和作用機制研究. 長沙: 湖南農業大學, 2011.
[20] Kang Z L, Yang Y H, Zhang L J. Molecular mechanism of responsing to drought stress in plants. Journal of Maize Sciences, 2006, (2): 96-100.
康宗利, 楊玉紅, 張立軍. 植物響應干旱脅迫的分子機制. 玉米科學, 2006, (2): 96-100.
[21] Wang Y N, Liu C, Meng K,etal. Effects of exogenous carbon source on carbon and nitrogen metabolism of wheat under drought stress. Journal of Henan Agricultural Sciences, 2015, (10): 29-34.
王雅楠, 劉存, 孟珂, 等. 外源性碳源對干旱脅迫下小麥幼苗碳氮代謝的影響. 河南農業科學, 2015, (10): 29-34.
[22] Sasaki-Sekimoto Y, Taki N, Obayashi T,etal. Coordinated activation of metabolic pathways for antioxidants and defence compounds by jasmonates and their roles in stress tolerance inArabidopsis. Plant and Cell Physiology, 2006, 47S: 233.
[23] Abebe T, Melmaiee K, Berg V,etal. Drought response in the spikes of barley: gene expression in the lemma, palea, awn, and seed. Functional & Integrative Genomics, 2010, 10(2): 191-205.
[24] Xu Y, Liu R, Yan L,etal. Light-harvesting chlorophyll a/b-binding proteins are required for stomatal response to abscisic acid inArabidopsis. Journal of Experimental Botany, 2012, 63(3); 1095-1106.
[25] Pan L, Zhang X Q, Wang J P,etal. Transcriptional profiles of drought-related genes in modulating metabolic processes and antioxidant defenses inLoliummultiflorum. Frontiers in Plant Science, 2016, 7: 519.
DifferentialgeneanalysisofPoapratensisinresponsetodroughtstress
LENG Nuan1, LIU Xiao-Wei2, ZHANG Na1, XU Li-Xin1*
1.InstituteofTurfgrassScience,BeijingForestryUniversity,Beijing100083,China; 2.NanjingOldMountainForestFarm,Nanjing211811,China
Drought is one of the main factors affecting the productivity ofPoapratensis. This study was undertaken to reveal the change of gene expression profile ofP.pratensisin the absence of a reference genome. The high-throughput Illumina Hiseq sequencing platform was used to investigate the transcriptome of a control group (CK) and a drought treated group (D) ofP.pratensis. The sequencing data were subsequently analyzed to help reveal the molecular mechanisms of drought response inP.pratensis. The results showed that 24465 differentially expressed genes were detected under drought treatment. After screening 4143 up-regulated genes and 4415 down-regulated genes were obtained, accounting for 34.98% of the total number of differentially expressed genes. The genes related to protein kinase, protein phosphatase, carbon metabolism and ABA can be used as the main research object to study the drought response mechanism ofP.pratensis. qRT-PCR analysis showed that the expression of 8 randomly differentially expressed genes was consistent with the high-throughput sequencing results. In addition, a series of genes for indole-3-glycerophosphate synthase, protein phosphatase, carbohydrate kinase, calcium binding protein and chlorophyll a/b binding protein were selected as candidate genes for drought stress related toP.pratensis. This laid the foundation for potentially revealing the molecular mechanism of drought response inP.pratensis.
Poapratensis; drought stress; high-throughput sequencing; transcription group
10.11686/cyxb2017130http//cyxb.lzu.edu.cn
冷暖, 劉曉巍, 張娜, 許立新. 草地早熟禾干旱脅迫轉錄組差異性分析. 草業學報, 2017, 26(12): 128-137.
LENG Nuan, LIU Xiao-Wei, ZHANG Na, XU Li-Xin. Differential gene analysis ofPoapratensisin response to drought stress. Acta Prataculturae Sinica, 2017, 26(12): 128-137.
2017-03-21;改回日期:2017-06-14
中國林學會——青年人才托舉工程項目資助。
冷暖(1992-),女,山東威海人,在讀碩士。E-mail:lengnuan99@163.com*通信作者Corresponding author. E-mail:lixinxu@bjfu.edu.cn
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
電子制作(2018年18期)2018-11-14 01:48:24
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
山東工業技術(2016年15期)2016-12-01 05:31:22
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
山東女子學院學報(2014年6期)2014-03-01 02:24:55
