魚源腐敗希瓦氏菌碳源利用及動力學分析
2018-01-04 05:43:22修艷輝郭全友韓承義陳慶榮
食品科學 2018年2期
關鍵詞:利用
修艷輝,郭全友*,韓承義,陳慶榮
魚源腐敗希瓦氏菌碳源利用及動力學分析
修艷輝1,2,郭全友1,*,韓承義3,陳慶榮4
(1.中國水產科學研究院東海水產研究所,上海 200090;2.上海海洋大學食品學院,上海 201306;3.寧德市益豐水產有限公司,福建 寧德 352100;4.寧德市蕉城區水產技術推廣站,福建 寧德 352100)
為探究溫度對魚源腐敗希瓦氏菌碳源利用的影響,利用多孔平板技術獲得5、15、25 ℃和33 ℃條件下其生長動態,采用修正的Gompertz方程,構建動力學模型,獲取動力學參數,結合孔平均顏色變化率及利用面積,探究其碳源利用效果。結果表明,25 ℃時最大比生長速率(μmax)最大,遲滯期(λ)最小,總體碳源利用能力和活性最強,15 ℃時次之,5 ℃時活性降至約25 ℃的1/6,33 ℃時最弱;5~25 ℃范圍內,溫度和 μ、 呈現良好的線性關系。5~33 ℃時糖類和羧酸類利用率較高,比例分別為30%、29%。糖類(糊精、D-麥芽糖、α-D-葡萄糖、D-阿拉伯醇、水蘇糖)、羧酸類(L-蘋果酸、L-乳酸、乙酰乙酸)、氨基酸類(L-絲氨酸、氨基乙酰-L-脯氨酸、L-天冬氨酸和L-谷氨酸)和胺/酰胺類、脂肪酸/脂類和其他類中(明膠、丙酮酸甲酯、吐溫40、L-組胺)利用較好。通過對腐敗希瓦氏菌碳源利用分析,通過對腐敗希瓦氏菌碳源利用和動力學分析,可為深入其代謝活性與水產品營養相關性等提供理論依據。
腐敗希瓦氏菌;碳源;生長動力學;多孔平板計數
水產品腐敗與微生物代謝活動密切相關,魚體表面、腮和內臟等附著大量的細菌,其中假單胞菌屬(Pseudomonas spp.)、莫拉氏菌(Moraxella spp.)、不動桿菌屬(Acinetobacter spp.)、黃桿菌屬(Flavobacterium spp.)和希瓦氏菌屬(Shewanella spp.)等是導致低溫貯藏魚類腐敗的主要微生物,菌量達到107~109CFU/g時,會產生難聞的氣味,失去食用價值,達到貨架期[1]。腐敗希瓦氏菌(Shewanella putrefaciens)作為低溫貯藏海水魚類的優勢腐敗菌,具有較強腐敗能力[2],同時能黏附在食品表面并形成生物膜[3],增加致病潛力,可能引發腹膜炎、軟組織感染等疾病[4]。改變外界因素(pH值、水分活度、NaCl、溫度和保鮮劑等)可抑制腐敗希瓦氏菌生長,例如高溫(37 ℃)、低pH值(pH 4)或高鹽分(12%)可抑制腐敗希瓦氏菌生長[5],茶多酚等保鮮劑能較好抑制其生長[6]。同時,微生物在受到環境因子脅迫作用下,進行自我修復過程中,需要不斷消耗能量來維持內部平衡,當能量耗盡時促使細菌代謝衰竭而失活甚至死亡,產生“自動殺菌”效應,這為微生物從生長狀態轉變為非生長狀態的客觀出現提供一種能量學解釋[7],但目前鮮見關于腐敗希瓦氏菌碳源利用及動力學代謝等相關研究。
基于氧化還原反應的多孔平板技術多用于微生物菌株鑒定和微生物群落分析,根據微生物種類和固有性質分析微生物對不同單一碳源的利用能力,如周麗英等[8]通過分析碳源平均顏色變化率(average well color development,AWCD)隨時間的變化分析3 種假單胞菌碳源利用差異,柏中中等[9]利用Biolog分析兩株凝結芽孢桿菌的乳酸發酵相關代謝能力,陳玉梅等[10]分析以培養基為基礎的乳酸鈉和葡萄糖對薛氏丙酸桿菌代謝物的抑制作用,劉文等[11]采用單碳源純培養等技術研究純菌株對碳源利用的選擇性。分析微生物代謝情況常采用AWCD、曲線擬合面積、Shannon指數和Shannon均勻度等多樣性指數[12],這些指標在微生物群落分析中均有較廣泛應用,在比較不同碳源利用情況時具有一定局限性。
常采用Logistic、Gompertz或Richard等模型分析環境因素對菌株生長的影響[2],如Speranza等[13]采用修正的Gompertz模型擬合腐敗希瓦氏菌等混合菌株生長數據,探究脅迫效應(如氣調包裝和天然抗菌劑)對其生長的影響。目前預測模型運用主要用于生長數據的擬合,亦可對其代謝產物等進行分析,如構建的大黃魚腐敗細菌PLFAs初級模型具有一定實際價值[14],但已有研究發現水解效應使得腐敗希瓦氏菌等菌生長和酶數據建模無法實現[15]。
本實驗以低溫貯藏大黃魚優勢腐敗菌(腐敗希瓦氏菌)為對象,研究溫度(5 、15、25 ℃和33 ℃)對腐敗希瓦氏菌碳源利用的影響,采用修正的Gompertz模型進行數據擬合,求出動力學參數,并結合AWCD和積分面積等分析其利用能力及差異性,為高效抑制低溫貯藏魚類優勢腐敗菌生長,延長貨架期等提供理論依據。
1 材料與方法
1.1 材料與試劑
從腐敗冷藏大黃魚中分離出優勢菌并經16S rRNA鑒定(KY859796),凍干保藏備用。
HCl標準溶液(0.01 mol/L) 深圳市博材達科技有限公司;營養瓊脂、營養肉湯 國藥集團化學試劑上海有限公司。
1.2 儀器與設備
MIR-153/253低溫培養箱 日本三洋公司;微生物生長自動鑒定儀 美國Biolog公司;SW-CJ-1FB型潔凈工作臺 上海博訊實業有限公司醫療設備廠。
1.3 方法
1.3.1 Biolog實驗
凍干腐敗希瓦氏菌在營養肉湯中活化后,取一株菌于營養瓊脂平板進行兩代劃線,然后從平板中取適量菌株接種到Biolog專業接種液(IF-A)中,調節透光率為95%~98%之間,使其光密度(OD)約為0.01。然后使用Biolog專用移液器,分別吸取200 μL菌懸液到GenⅢ 96 孔板中,分別置于5、15、25 ℃和33 ℃的培養箱中培養96 h,間隔適當時間經Biolog儀中的Microstation讀數儀測定OD值。
1.3.2 腐敗希瓦氏菌對碳源利用能力分析
將GenⅢ板中的71 種碳源分為6 類,分別為糖類(25 種)、羧酸類(17 種)、氨基酸類(10 種)、胺/酰胺類(6 種)、脂肪酸/脂類(6 種)和其他類(7 種)[16]。當腐敗希瓦氏菌V穩(穩定時OD值,即C-R值)大于0.25時[17],判為可利用,V穩不大于0時,判為完全不能利用。
碳源利用能力可用孔AWCD、利用面積S和動力學參數(μmax和λ)進行描述,AWCD和S計算如式(1)和(2)所示:

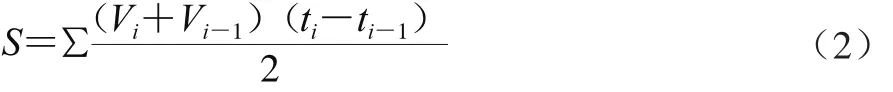
式中:R為對照孔OD值;C為含有碳源孔的OD值;n為碳源孔數;Vi、Vi-1分別為i、i-1時刻的(C-R)值。
1.3.3 模型建立及評價
初級模型采用修正的Gompertz方程擬合Biolog數據[18],方程見式(3),平方根方程[19]可描述環境因子對動力學參數的影響,方程見(4)和(5):
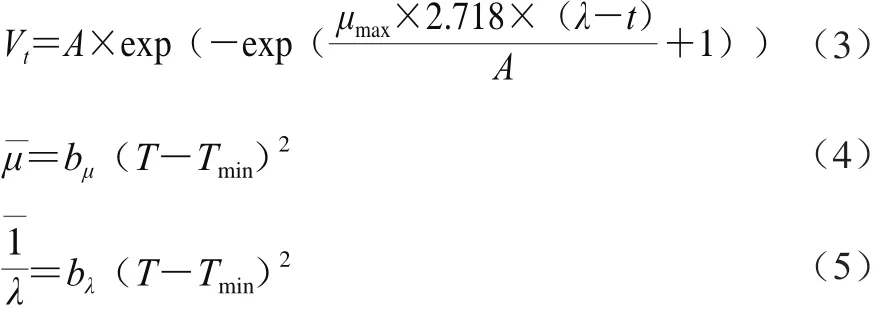
式中:t為時間/h;Vt為t時的(C-R)值;μmax為顏色變化最大速率/h-1;λ為遲滯期/h;A為顏色變化最大水平;T為溫度/℃;Tmin為理論最小溫度/℃;bμ、bλ為方程常數。
采用決定系數R2、準確度Af、精確度Bf對模型進行評價[20-21],R2接近1表明擬合較好,Af和Bf值越接近1,準確性或精確性較好[22]。
1.4 數據分析
數據采用SPSS 19.0軟件(美國IBM公司),采用最小二乘算法,進行非線性回歸,求出動力學參數,采用Origin 8.0軟件(美國OriginLab公司)作圖。
2 結果與分析
2.1 腐敗希瓦氏菌碳源利用情況及模型評價
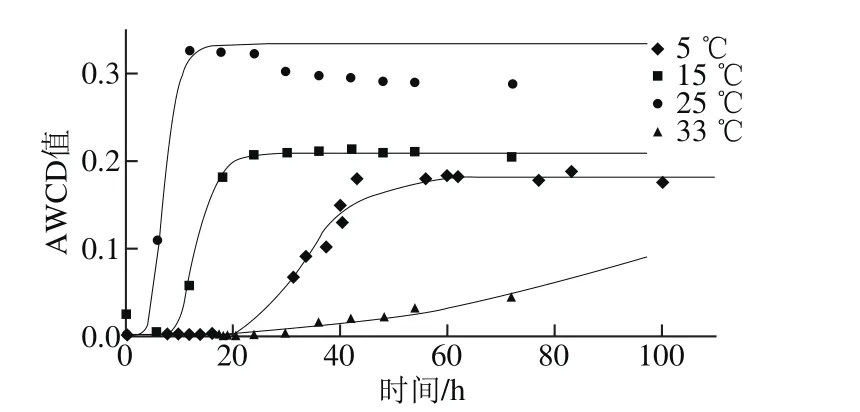
圖1 不同溫度條件下腐敗希瓦氏菌總體碳源利用狀況Fig. 1 Carbon utilization of S. putrefaciens at different temperatures
由圖1可知,5~25 ℃時總體碳源利用曲線呈S型,25 ℃時μmax最大(0.064 h-1),λ最小(4.5 h),AWCD最大值為0.329,表明總體碳源利用能力和活性最強,15 ℃時次之,μmax降至25 ℃時的約1/2,λ增大約2 倍,AWCD最大值為0.216;5 ℃時μmax降至25 ℃時的約1/6,λ增至約5.6 倍,最大AWCD值為0.183;33 ℃時最弱,μmax接近0,λ長達61.4 h,AWCD最大值為0.091。結果表明,溫度對腐敗希瓦氏菌碳源利用影響較大,最大AWCD值隨著溫度升高而變小;λ隨著溫度升高先變小后變大,μmax則先變大后變小(表1)。
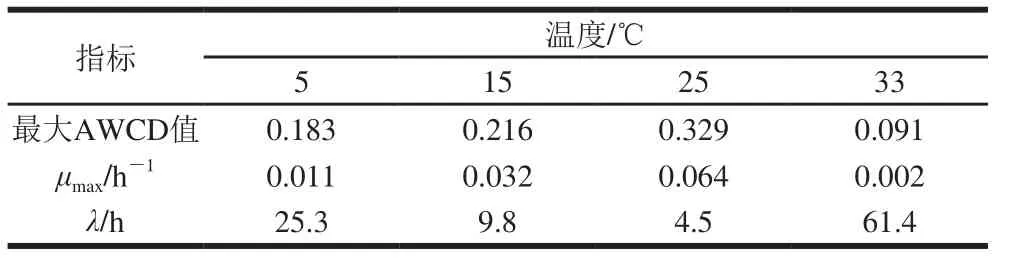
表1 不同溫度條件下腐敗希瓦氏菌動力學參數及AWCD值Table 1 Growth kinetic parameters and AWCD values of S. putrefaciens at different temperatures
桂萌等[23]對0~20 ℃托盤包裝冰藏鱘魚中的優勢腐敗菌——腐敗希瓦氏菌用修正的Gompertz方程擬合得到動力學參數,結果顯示0~20 ℃條件下μmax逐漸變大,λ逐漸變小,最大菌數變化不大,在8.57~9.8(lg(CFU/mL))之間,與本研究中5~25 ℃時碳源μmax和λ變化規律相似。動力學參數(μmax和λ)通過擬合平方根模型如下:
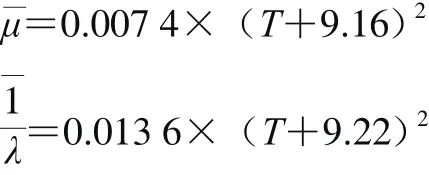
其中R2分別為1和0.996,表明5~25 ℃范圍內,溫度和、呈現良好的線性關系,但5~33 ℃時擬合效果較差,表明腐敗希瓦氏菌最適溫度在25 ℃,故在5~25 ℃內溫度可與、呈現良好的線性關系。本研究中擬合的模型R2為0.991±0.004,Af為0.998±0.025,Bf為1.013±0.033,表明模型能較好擬合5~33 ℃范圍內腐敗希瓦氏菌碳源利用情況。
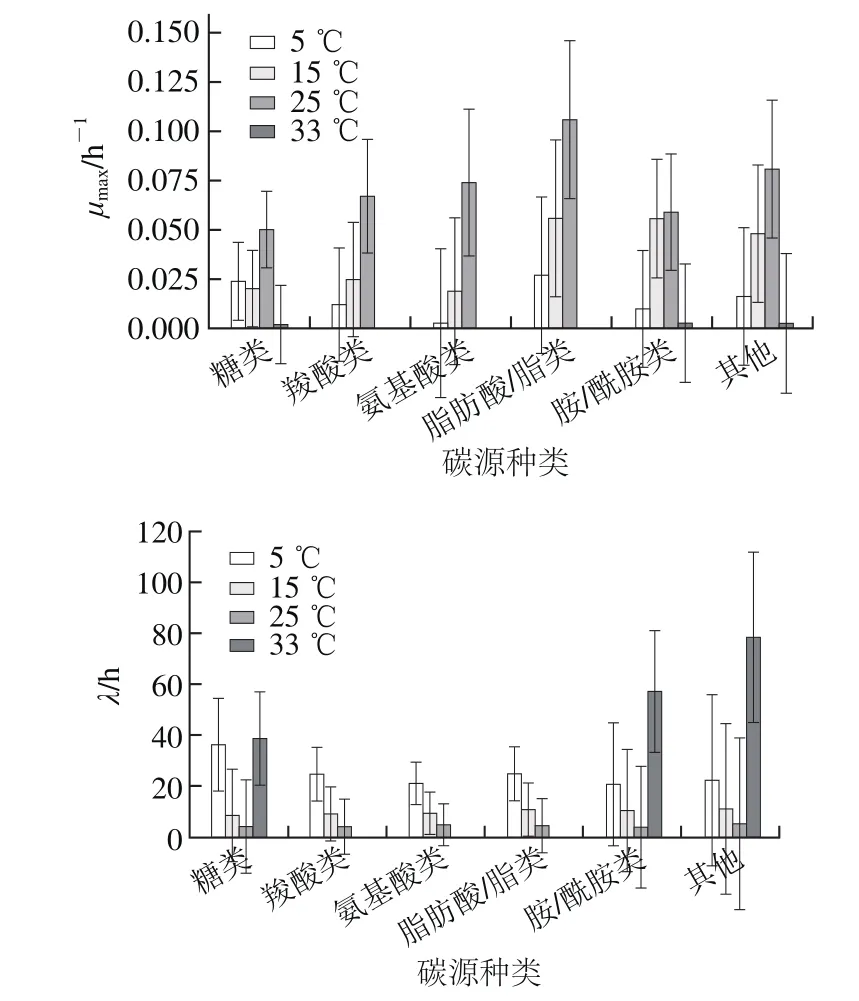
圖2 溫度對腐敗希瓦氏菌各類碳源利用的影響Fig. 2 Effect of temperature on the utilization of different carbon sources by S. putrefaciens
由圖2可知,5~25 ℃內,腐敗希瓦氏菌對各類碳源(除糖類)的μmax隨著溫度升高而增大,λ呈現相反的規律,如5~25 ℃時羧酸類μmax分別為0.012、0.025 h-1和0.067 h-1,λ分別為24.6、9.3 h和4.3 h。5~25 ℃時各類碳源中脂肪酸/脂類利用速率最快,如25 ℃時脂肪酸/脂類利用速率為0.106 h-1,糖類速率為0.05 h-1。33℃時羧酸類、氨基酸類和脂肪酸類基本不利用。相同溫度下各類碳源λ變化不明顯,但μmax變化明顯。
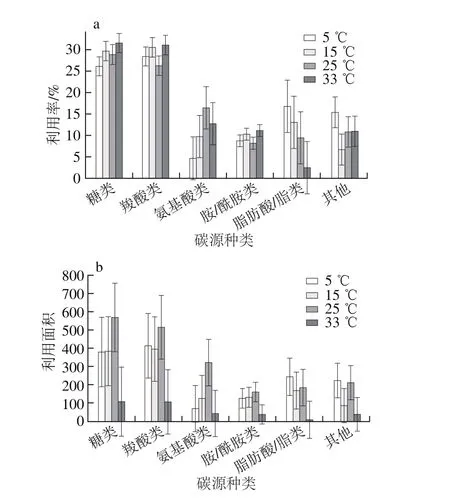
圖3 溫度對腐敗希瓦氏菌6類碳源利用率(a)及利用面積(b)的影響Fig. 3 Utilization rates and areas of six carbon sources of S. putrefaciens at different temperatures
由圖3a可知,5~33 ℃時糖類和羧酸類利用率較高,平均值分別為(30±1.4)%、(29±2.7)%,且溫度對其影響不大;氨基酸類、脂肪酸/脂類和其他碳源利用比例受溫度影響較大,如5 ℃時氨基酸類利用率為4.7%,25 ℃時增至16.4 %,而33 ℃降至12.7%。由圖3b可知,溫度對腐敗希瓦氏菌利用不同類型碳源的能力有較大影響,如5 ℃和15 ℃時糖類以及羧酸類利用面積變化不大,分別約為380和400,但33 ℃時分別降至107和106。溫度對碳源利用情況受微生物種類、外界環境及碳源種類的影響,如有研究表明小麥及玉米秸稈腐解微生物對供試碳源的利用隨著溫度升高而降低,而且秸稈腐解微生物利用糖類、脂類和醇類能力較強,利用氨基酸和酸類能力較弱[24]。也有研究表明15 ℃時腐敗希瓦氏菌的脫色能力明顯低于在25 ℃和35 ℃[25]。劉文等[11]研究表明相似碳鏈長度碳源所培養出的微生物種類組成相似,潛在地說明碳鏈長度可能影響微生物對有機碳的利用,而微生物對碳鏈長度選擇的主要原因可能是由于其生理代謝功能限制。腐敗希瓦氏菌生長的細胞內代謝物在污水中的組成比較復雜,不同生長基質中代謝產物差異較大,作為限制碳源的甲酸鹽在污水中濃度降低[26]。
2.2 溫度對腐敗希瓦氏菌碳源利用影響
2.2.1 不同溫度條件下腐敗希瓦氏菌糖類的代謝利用分析
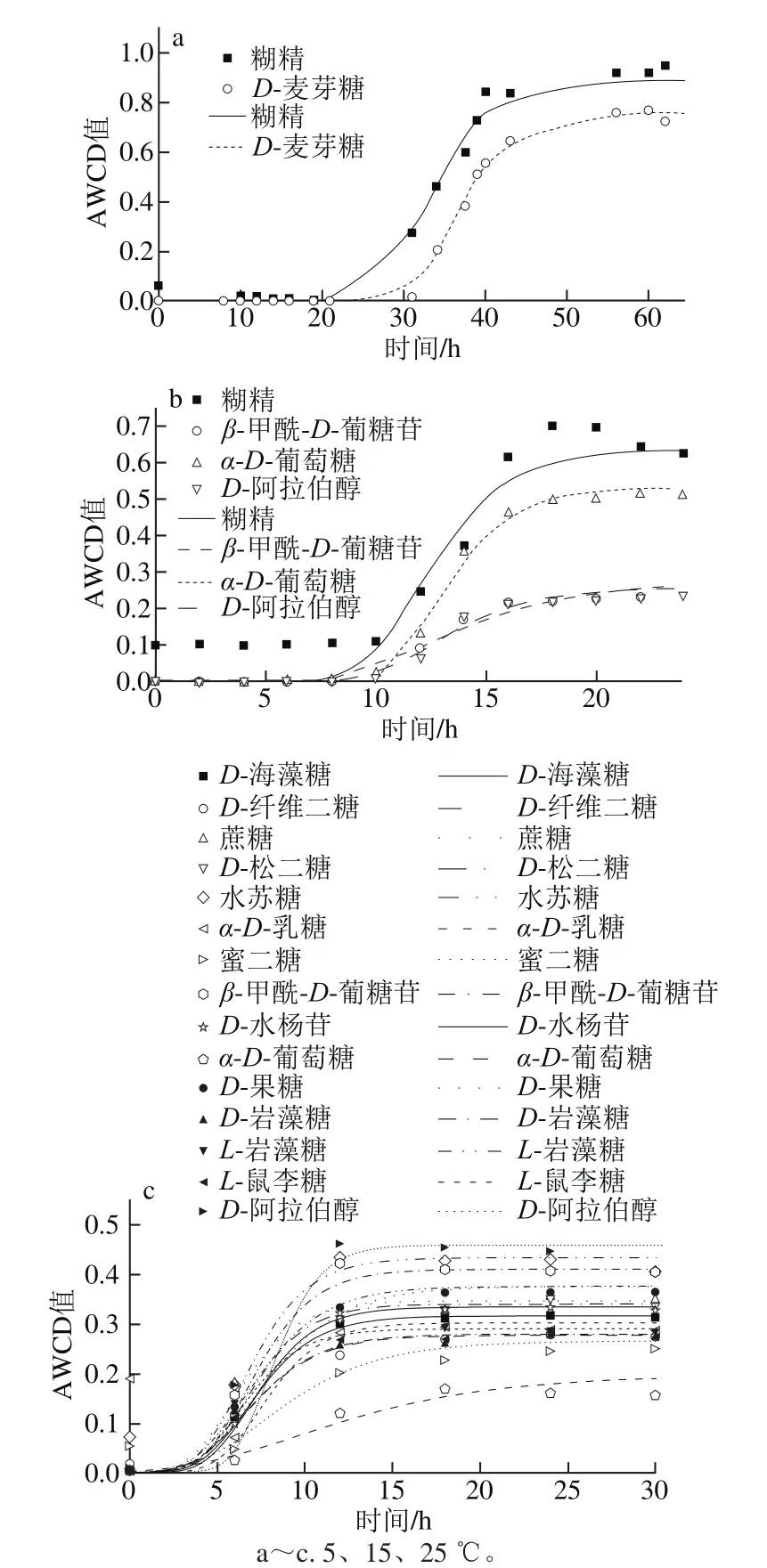
圖4 不同溫度條件下糖類AWCD隨培養時間的變化Fig. 4 Changes in AWCD of carbohydrates during incubation at different temperatures
如圖4所示,5~25 ℃內腐敗希瓦氏菌對糖類的利用整體上呈“S”型,且溫度越高,代謝反應越強烈,延滯期越短,達到穩定期的時間越短。5 ℃時μmax為(0.051±0.061)h-1,ˉλ為(24.5±9.8)h;15 ℃時μmax為(0.071±0.043)h-1,ˉλ為(9.1±0.8)h;25 ℃時μmax為(0.084±0.024)h-1,ˉλ為(5.0±0.7)h;33 ℃時皆不可利用。不同溫度達到穩定期的時間也不同,順序依次為25 ℃(12 h)<15 ℃(15 h)<5 ℃(17 h以上)。5℃時利用糊精和D-麥芽糖較好,μmax分別為0.032 h-1和0.069 h-1,λ分別為17.6 h和31.4 h。15 ℃時糊精和α-D-葡萄糖利用較好,μmax分別為0.111、0.106 h-1,β-甲酰-D-葡糖苷和D-阿拉伯醇利用較差,μmax分別為0.028、0.040 h-1;其λ在8.7~10.7 h之間,差異不大(P>0.05)。25 ℃時D-阿拉伯醇、水蘇糖利用較好,μmax分別為0.111 h-1和0.08 h-1,其次是β-甲酰-D-葡糖苷、D-松二糖和蔗糖,μmax分別為0.075、0.066 h-1和0.065 h-1;而這15 種糖類λ與ˉλ((5.0±0.6)h)之間沒有顯著差異(P>0.05)。說明腐敗希瓦氏菌對糖類的代謝能力受培養溫度的影響較大。
本研究中糖類共有25 種(糊精、D-麥芽糖、D-海藻糖、D-纖維二糖、龍膽二糖、蔗糖、D-松二糖、水蘇糖、蜜三糖、α-D-乳糖、蜜二糖、β-甲酰-D-葡糖苷、D-水楊苷、α-D-葡糖糖、D-甘露糖、D-果糖、D-半乳糖、3-甲酰葡糖、D-巖藻糖、L-巖藻糖、L-鼠李糖、D-山梨醇、D-甘露醇、D-阿拉伯醇和果膠),溫度對糖類μmax影響較大,進而影響可利用碳源種類,如25 ℃可利用糖類種類遠大于15 ℃(15/4),而33 ℃時可利用碳源種類為0 種,可見25 ℃糖類碳源利用種類更好。但也有部分碳源反而隨著溫度升高利用變差,如糊精在5~33 ℃范圍內V穩值依次為0.795、0.534、0.247和0。此外,15 ℃時只有3-甲酰葡糖完全不能利用,25 ℃時D-麥芽糖和D-山梨醇完全不能利用,33 ℃糊精和D-麥芽糖完全不能利用。
有研究表明腐敗希瓦氏菌能利用葡萄糖、蔗糖作為碳源,而無法利用乳糖和山梨醇,利用甘露醇時利用速度較慢[25],與本研究中腐敗希瓦氏菌可利用葡萄糖和蔗糖作為碳源、不能利用山梨醇及利用甘露醇較慢(V穩=0.18)相一致,但本研究中腐敗希瓦氏菌可利用α-D-乳糖,且在25 ℃時μmax達到0.06 h-1,λ為5.3 h,這可能是由于33 ℃時溫度過高導致分解乳糖的相關酶失活。有人研究中與腐敗希瓦氏菌ATCC8071T的16S rDNA序列相似值為97%的中國希瓦氏菌(Shewanella cinica)D14T,可利用D-半乳糖、D-葡萄糖、蔗糖等多種有機物為碳源[27],與本研究中結果類似,只是對半乳糖的利用較差(V穩<0.25)。從能量角度考慮,小分子碳源物質更容易被利用且能耗低,但對于某些微生物類群來說,二糖或低聚糖比單糖更容易利用[11]。
2.2.2 不同溫度條件下腐敗希瓦氏菌羧酸類的代謝利用分析
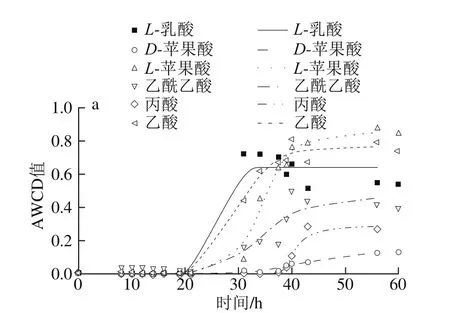
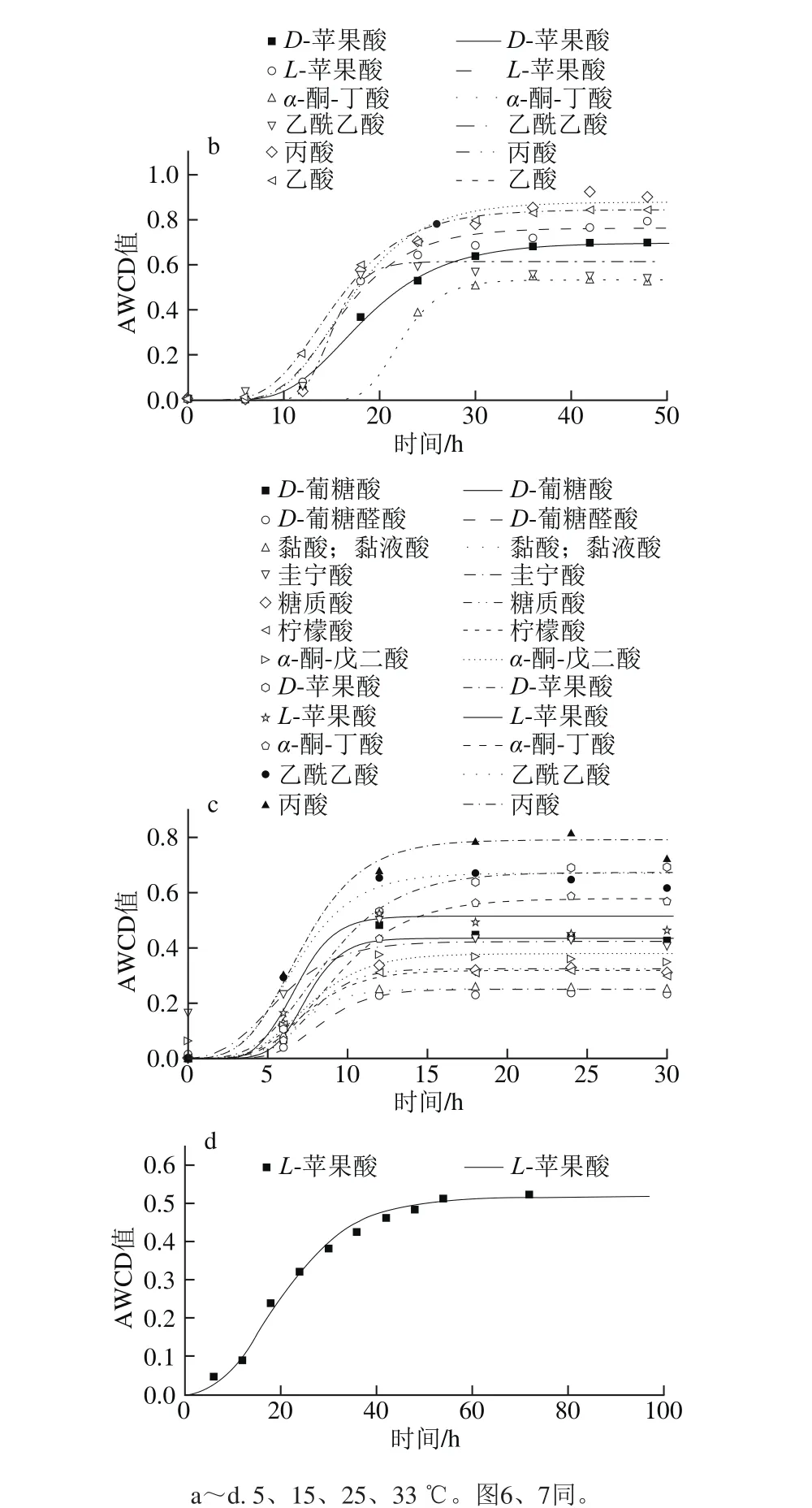
圖5 不同溫度條件下羧酸類AWCD隨培養時間的變化Fig. 5 Changes in AWCD of carboxylic acids during incubation at different temperatures
由圖5可知,曲線皆呈現“S”型,5 ℃時μmax為(0.067±0.038)h-1,λ為(29.1±6.0)h;15 ℃時μmax為(0.072±0.020)h-1,λ為(11.6±3.7)h;25 ℃時μmax為(0.087±0.033)h-1,λ為(4.5±1.0)h;33 ℃時只能利用L-蘋果酸,μmax為0.02 h-1,λ為7.2 h,5~25 ℃下達到穩定期的時間順序為25 ℃(15 h)<15 ℃(30 h)<5 ℃(40 h以上)。
5 ℃時能較好利用L-蘋果酸和L-乳酸,μmax分別為0.092 h-1和0.085 h-1,λ分別為29.8 h和21.2 h;其次為乙酸和丙酸,μmax分別為0.079 h-1和0.082 h-1,λ為25.2 h和38.6 h;D-蘋果酸利用較差,μmax為0.007 h-1,λ為32.5 h。15 ℃時丙酸和乙酸利用較好,μmax分別為0.066 h-1和0.063 h-1,λ為10.1 h和8.5 h;此外,α-酮-丁酸和乙酰乙酸μmax最大(0.121、0.076 h-1),但是其最大AWCD值最小,λ長(18.7、12.4 h),其利用情況不如丙酸和乙酸。25 ℃時,能利用較好丙酸、乙酰乙酸和D-蘋果酸,μmax分別為0.121、0.108 h-1和0.085 h-1,λ差異不大,均值為(3.9±0.5)h,最大AWCD值為0.65~0.8;此外利用較好的有α-酮-丁酸和L-蘋果酸,μmax分別為0.073 h-1和0.146 h-1,λ為5.1 h和4.9 h;雖然L-蘋果酸μmax最大,但其最大AWCD值較小,利用面積較小,所以丙酸等利用情況更好,葡糖酸和糖質酸(0.139、0.077 h-1)與之類似;D-糖醛酸和黏酸利用較差,μmax最大,AWCD值皆較小,導致利用面積較小,利用情況較差。33 ℃時只有L-蘋果酸利用較好,μmax為0.02 h-1,λ為7.5 h,雖然其最大AWCD值與25 ℃時L-蘋果酸接近,但是其對數期明顯較長,導致其利用速率降低。
對17 種羧酸類(D-半乳糖醛酸、D-葡糖酸、D-葡糖醛酸、黏酸、奎寧酸、糖質酸、p-羥基苯乙酸、L-乳酸、檸檬酸、α-酮-戊二酸、D-蘋果酸、L-蘋果酸、α-酮-丁酸、乙酰乙酸、丙酸、乙酸和甲酸)利用進行分析。5 ℃時p-羥基-苯乙酸和甲酸完全不能利用,15 ℃時黏酸、p-羥基-苯乙酸、檸檬酸和甲酸完全不能利用,25 ℃時D-半乳糖醛酸、p-羥基-苯乙酸、L-乳酸和甲酸完全不能利用,33 ℃時p-羥基-苯乙酸、L-乳酸、α-酮-丁酸、丙酸和甲酸完全不能利用。樂毅全等[25]關于腐敗希瓦氏菌的脫色實驗表明乙酸、丙酸和丁酸是該菌較理想的碳源,而草酸、檸檬酸和戊酸則不能被該菌用于降解染料,本研究中乙酸和丙酸在5~25 ℃利用較好,而檸檬酸僅在25 ℃時利用較好。本研究結果與上述研究中報道的腐敗希瓦氏菌對碳源的利用類似。
2.2.3 不同溫度條件下腐敗希瓦氏菌氨基酸類的代謝利用分析
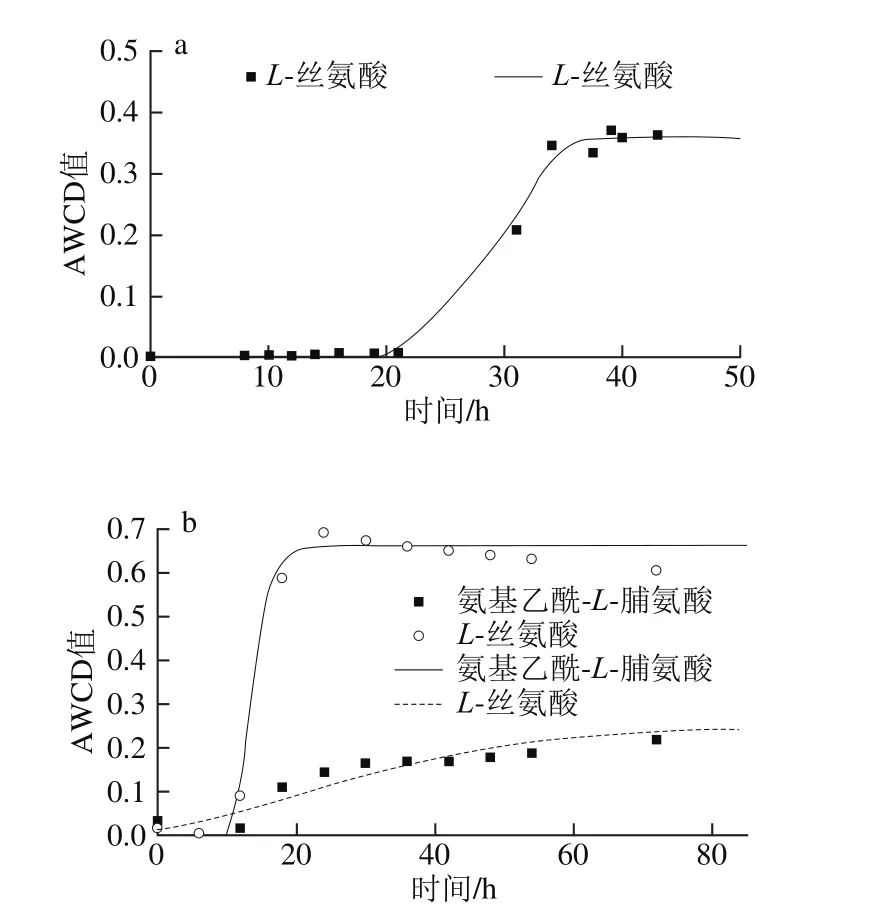
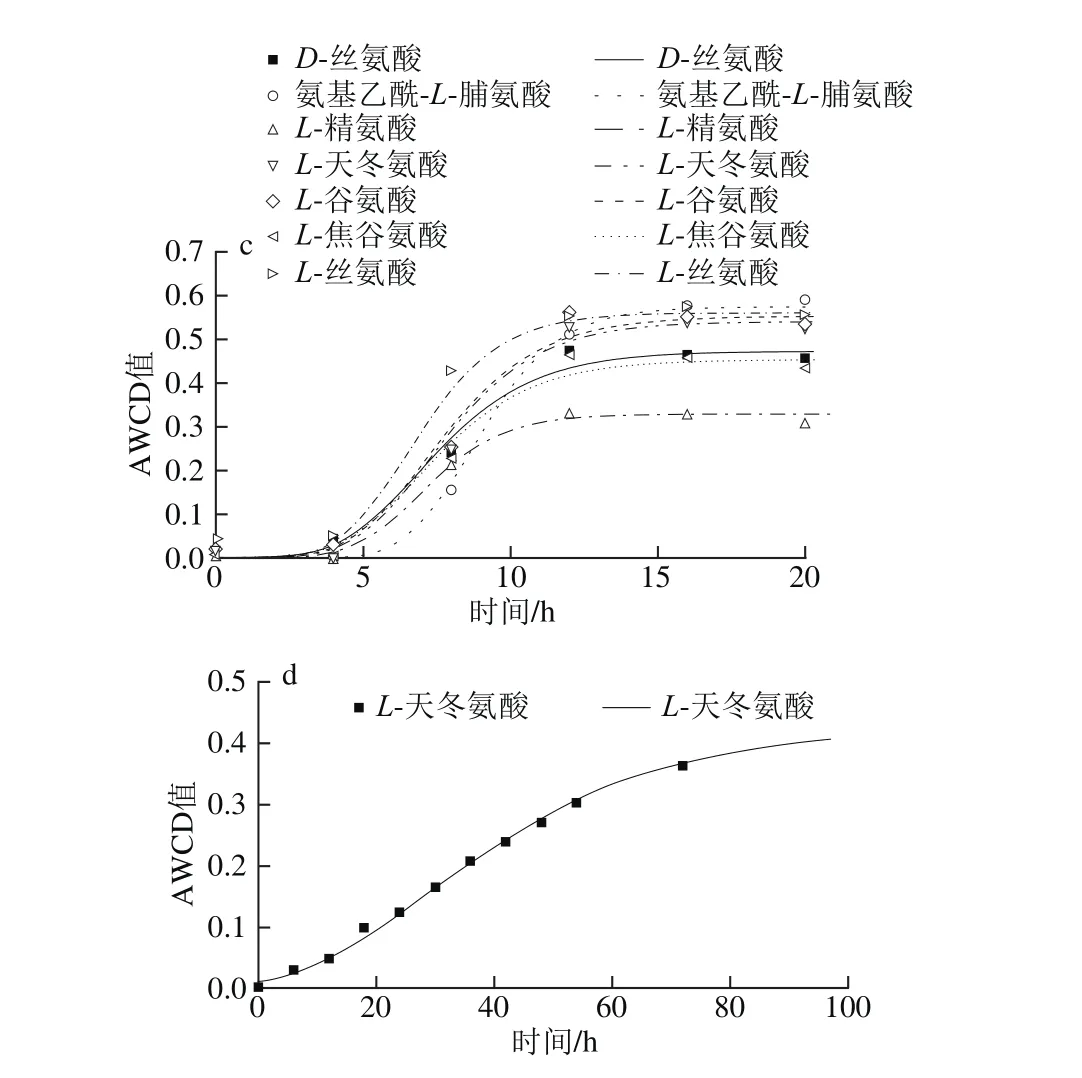
圖6 不同溫度條件下氨基酸類AWCD隨培養時間的變化Fig. 6 Changes in AWCD of amino acids during incubation at different temperatures
由圖6可知,曲線皆呈現“S”型。5 ℃時只有L-絲氨酸利用較好,μmax為0.109 h-1,λ為29.0 h;15 ℃時L-絲氨酸和氨基乙酰-L-脯氨酸利用較好,μmax分別為0.142 h-1和0.005 h-1,λ分別為11.2 h和1.2 h,但兩者達到平穩期的時間差距較大,L-絲氨酸在20 h左右達到穩定期,氨基乙酰-L-脯氨酸在70 h左右達到平穩期。25 ℃時利用較好的有氨基乙酰-L-脯氨酸、L-絲氨酸、L-天冬氨酸和L-谷氨酸,其μmax分別為0.144、0.13、0.103 h-1和0.102 h-1,λ分別為6.9、4.6、5.1 h和4.9 h;其次利用較好的有L-焦谷氨酸和D-絲氨酸,μmax分別為0.085 h-1和0.082 h-1,λ皆為4.6 h。33 ℃僅有L-天冬氨酸利用較好,μmax為0.007 h-1,λ為7.1 h,對數期很長,約90 h達到平穩期。
對10 種氨基酸(D-天冬氨酸、D-絲氨酸、氨基乙酰-L-脯氨酸、L-丙氨酸、L-精氨酸、L-天冬氨酸、L-谷氨酸、L-焦谷氨酸、L-絲氨酸、γ-氨基-丁酸)利用分析表明,腐敗希瓦氏菌只在5~15 ℃時完全不能利用天冬氨酸和D-絲氨酸,33 ℃時氨基乙酰-L-脯氨酸、L-丙氨酸、L-天冬氨酸和L-絲氨酸完全不能利用。丙氨酸利用較差(AWCD<0.25),天冬氨酸在25~33 ℃利用較好。研究顯示,腐敗希瓦氏菌是變形細菌γ紫色家族的第一個利用絲氨酸途徑的成員,在其適應厭氧環境的能力起重要作用[26]。
2.2.4 不同溫度條件下腐敗希瓦氏菌脂肪酸/脂類、胺/酰胺類及其他的代謝利用分析
由圖7可知,曲線皆呈現“S”型,5 ℃和33 ℃時λ比較接近,均值為(26.1±2.3)h(除5 ℃溴-丁二酸λ為40.2 h外),但5 ℃ μmax(0.114±0.070)h-1遠大于33 ℃時(μmax≤0.04 h-1)。15 ℃和25 ℃時平均μmax比較接近,分別為(0.120±0.063)h-1和(0.101±0.039)h-1,但15 ℃ˉλ約為25 ℃時的2 倍,分別為(11.0±1.2)h和(6.3±2.4)h。且不同溫度下達到平穩期的時間也不同,順序依次為25 ℃(12 h)<15 ℃(15 h)<5 ℃(35 h)<33 ℃(40 h以上)。
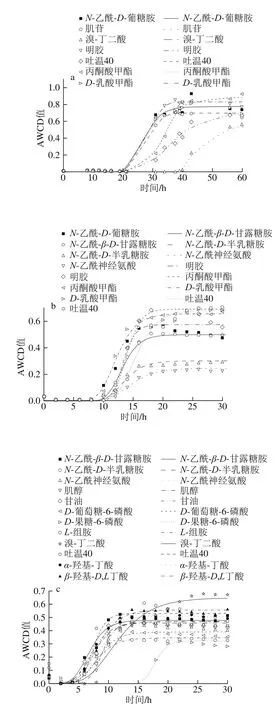
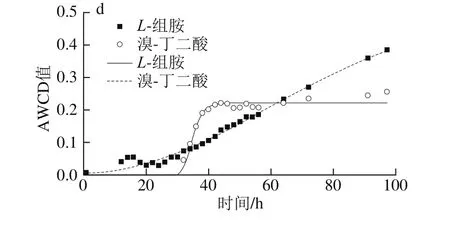
圖7 不同溫度條件下脂肪酸/脂類、胺/酰胺類及其他AWCD隨培養時間的變化Fig. 7 Changes in AWCD of fatty acids/lipids, amines/amides and other substances during incubation at different temperatures
5 ℃時能較好利用明膠和肌苷,μmax為0.234 h-1和0.19 h-1,λ分別為28.0 h和27.7 h;15 ℃時利用較好的為明膠和丙酮酸甲酯,μmax為0.212 h-1和0.208 h-1,λ為11.8 h和12.2 h,其次為N-乙酰-β-D-甘露糖胺、D-乳酸甲酯和吐溫40,μmax分別為0.113、0.123 h-1和0.116 h-1,λ分別為10.9、9.5 h和12.5 h;25 ℃時利用較好的有吐溫40和肌醇,μmax為0.188、0.163 h-1和0.131 h-1,λ為6.0 h和7.4 h,此外D-果糖-6-磷酸μmax較大(0.163 h-1),但λ較長(16.6 h),故利用差于吐溫40和肌醇。33 ℃只可利用L-組胺和溴-丁二酸,μmax小,λ長。
對6 種胺/酰胺類(N-乙酰-D-葡糖胺、N-乙酰-β-D-甘露糖胺、N-乙酰-D-半乳糖胺、N-乙酰神經氨酸、L-組胺和葡糖醛酰胺),6種脂肪酸/脂類(丙酮酸甲酯、吐溫40、α-羥基-丁酸、β-羥基-D,L丁酸、L-半乳糖醛酸內酯和D-乳酸甲酯),7種其他類(甘油、D-葡萄糖-6-磷酸、D-果糖-6-磷酸、明膠、溴-丁二酸、肌苷和肌醇)利用分析表明,5~15 ℃時,甘油、D-果糖-6-磷酸、葡糖醛酰胺和β-羥基-D,L丁酸完全不能利用,33 ℃時N-乙酰-D-葡糖胺、肌苷、明膠、葡糖醛酰胺、丙酮酸甲酯、D-乳酸甲酯和吐溫40完全不能利用。α-羥基-丁酸在低溫(5 ℃)和高溫時(33 ℃)都不能利用,溴-丁二酸只在15 ℃不能利用,L-半乳糖醛酸內酯在高溫(33 ℃)時可輕微利用(AWCD<0.25)。研究表明希瓦氏菌能夠分解明膠,利用吐溫40、肌苷和甘油等[28-29],明膠和殼聚糖等常作成復合膜用于水產品保鮮。同時,希瓦氏菌屬代謝產生較多脂肪酶,使用微生物分泌的脂肪酶有專一性強、活性高、副產物少、成本低和環保等優點[30]。因此,探究腐敗希瓦氏菌對明膠等物質的利用情況,可為進深入探究其代謝機理及環境保護等提供理論依據。
3 結 論
本研究利用多孔平板技術探究了5、15、25 ℃和33 ℃條件下,腐敗希瓦氏菌對71 種碳源的利用能力及不同溫度下其生長動力學,為深入研究腐敗希瓦菌能量代謝及作用機制提供一定理論支持。
25 ℃時總體碳源利用能力和活性最強,15 ℃時次之,5 ℃時活性降至約25 ℃的1/6,33 ℃時活性最弱,5~25 ℃范圍內,溫度和、 呈現良好的線性關系。
5~33 ℃時糖類和羧酸類利用率較高,平均值分別為(30±1.4)%、(29±2.7)%。5~33 ℃氨基酸類、脂肪酸/脂類和其他類碳源利用比例隨溫度變化影響較大。溫度對腐敗希瓦氏菌不同類型碳源的利用情況有較大影響:相同溫度下各類碳源λ變化不明顯,但μmax變化明顯。
腐敗希瓦氏菌碳源代謝情況為:糖類中,5℃時糊精和D-麥芽糖利用較好;15 ℃時糊精和α-D-葡萄糖利用較好;25℃時D-阿拉伯醇、水蘇糖利用較好;33 ℃普遍利用較差。羧酸類中,5 ℃時L-蘋果酸和L-乳酸利用較好;15 ℃時丙酸和乙酸利用較好;25 ℃時丙酸、乙酰乙酸和D-蘋果酸利用較好;33 ℃時L-蘋果酸利用較好。氨基酸類中,5 ℃時L-絲氨酸利用較好;15 ℃時L-絲氨酸和氨基乙酰-L-脯氨酸利用較好;25 ℃時氨基乙酰-L-脯氨酸、L-絲氨酸、L-天冬氨酸和L-谷氨酸利用較好;33 ℃時L-天冬氨酸利用較好。脂肪酸/脂類、胺/酰胺類及其他類中,5 ℃時明膠和肌苷利用較好;15 ℃時明膠、丙酮酸甲酯利用較好;25 ℃時吐溫40和肌醇利用較好;33 ℃時L-組胺和溴-丁二酸利用較好。
[1] SCHELEGUEDA L I, ZALAZAR A L, GLIEMMO M F, et al.Inhibitory effect and cell damage on bacterial flora of fish caused by chitosan, nisin and sodium lactate[J]. International Journal of Biological Macromolecules, 2016, 83: 396-402. DOI:10.1016/j.ijbiomac.2015.11.033.
[2] 郭全友, 修艷輝, 姜朝軍, 等. 環境因子對大黃魚腐敗希瓦氏菌生長影響的計數法分析[J]. 農業機械學報, 2016, 47(10): 250-257; 317.DOI:10.6041/j.issn.1000-1298.2016.10.031.
[3] ZHAO A, ZHU J, YE X, et al. Inhibition of biof i lm development and spoilage potential of Shewanella baltica by quorum sensing signal in cell-free supernatant from Pseudomonas fl uorescens[J]. International Journal of Food Microbiology, 2016, 230: 73-80. DOI:10.1016/j.ijfoodmicro.2016.04.015.
[4] LóPEZ A C, BOSH B E, CHAMORRO B I, et al. Peritonitis by Shewanella putrefaciens: apropos of a case[J]. Nefrologia, 2016, 36(4):444-445. DOI:10.1016/j.nefroe.2016.06.008.
[5] 修艷輝, 郭全友, 姜朝軍. pH、水分活度和NaCl對腐敗希瓦氏菌生長/非生長界限及生長動力學參數的影響[J]. 現代食品科技,2016(6): 156-162. DOI:10.13982/j.mfst.1673-9078.2016.6.025.
[6] 王慧敏, 朱軍莉, 陸海霞, 等. 茶多酚抑制腐敗希瓦氏菌機理研究[J]. 茶葉科學, 2014, 34(2): 149-155. DOI:10.3969/j.issn.1000-369X.2014.02.009.
[7] NYSTR M T, GUSTAVSSON N. Maintenance energy requirement:what is required for stasis survival of Escherichia coli?[J]. Biochimica et Biophysica Acta, 1998, 1365(1/2): 225-231. DOI:10.1016/S0005-2728(98)00072-3.
[8] 周麗英, 林素蘭, 李藝, 等. 水稻根際耐鎘細菌碳源代謝功能分析[J]. 中國生態農業學報, 2012, 20(8): 1083-1087. DOI:10.3724/SP.J.1011.2012.01083.
[9] 柏中中, 許婷婷, 何小丹, 等. 利用Biolog系統進行乳酸生產菌代謝能力的快速分析[J]. 食品與生物技術學報, 2009, 28(3): 347-351.DOI:10.3321/j.issn:1673-1689.2009.03.013.
[10] 陳玉梅, 常忠義, 王疆元, 等. 乳酸鈉和葡萄糖對薛氏丙酸桿菌生長及代謝物抑菌活性的影響[J]. 西北農林科技大學學報(自然科學版),2007, 35(2): 178-182. DOI:10.3321/j.issn:1671-9387.2007.02.036.
[11] 劉文, 楊漸, 吳耿, 等. 青藏高原北部湖泊沉積物中基于不同碳源可培養細菌多樣性[J]. 鹽湖研究, 2016(2): 92-101.
[12] 曹永昌, 譚向平, 和文祥, 等. 秦嶺地區不同林分土壤微生物群落代謝特征[J]. 生態學報, 2016(10): 2978-2986. DOI:10.5846/stxb201411222316.
[13] SPERANZA B, BEVILACQUA A, CONTE A, et al. Use of desirability approach to predict the inhibition of Pseudomonas fluorescens, Shewanella putrefaciens and Photobacterium phosphoreum in fi sh fi llets through natural antimicrobials and modif i ed atmosphere packaging[J]. Food and Bioprocess Technology, 2012,6(9): 2319-2330. DOI:10.1016/j.ajic.2003.10.004.
[14] 郭全友, 何木, 李保國, 等. 大黃魚腐敗細菌鑒定及PLFAs初級模型的建立[J]. 現代食品科技, 2015, 31(12): 227-234. DOI:10.13982/j.mfst.1673-9078.2015.12.034.
[15] BRAUN P, SUTHERLAND J P. Predictive modelling of growth and enzyme production and activity by a cocktail of Pseudomonas spp.,Shewanella putrefaciens and Acinetobacter sp.[J]. International Journal of Food Microbiology, 2003, 86(3): 271-282. DOI:10.1016/S0168-1605(02)00564-0.
[16] 鄭華, 陳法霖, 歐陽志云, 等. 不同森林土壤微生物群落對Biolog-GN板碳源的利用[J]. 環境科學, 2007, 1(5): 1126-1130. DOI:10.3321/j.issn:0250-3301.2007.05.034.
[17] 李志斐, 王廣軍, 謝駿, 等. 草魚養殖池塘生物膜固著微生物群落碳代謝Biolog分析[J]. 水產學報, 2014, 38(12): 1985-1995. DOI:10.3724 1231.2014.49435.
[18] VERSCHUERE L, FIEVEZ V, VOOREN L V, et al. The contribution of individual populations to the Biolog pattern of model microbial communities[J]. Fems Microbiology Ecology, 1997, 24(4): 353-362.DOI:10.1016/s0168-6496(97)00076-7.
[19] 王凱, 葉可萍, 白紅武, 等. 冷卻豬肉中單增李斯特菌的定量暴露評估[J]. 食品科學, 2016, 37(11): 79-83. DOI:10.7506/spkx1002-6630-201611014.
[20] 周晏, 周國燕, 徐斐, 等. 單增李斯特菌在生食魚片中生長模型的建立[J]. 食品科學, 2015, 36(15): 157-162. DOI:10.7506/spkx1002-6630-201515029.
[21] 劉亞兵, 何臘平, 高澤鑫, 等. 基于假單胞菌生長模型預測冷卻牛肉的貨架期[J]. 中國釀造, 2017, 36(8): 114-119. DOI:10.11882/j.issn.0254-5071.2017.08.025.
[22] 黃丹陽, 曹慧, 徐斐, 等. 米飯中蠟樣芽孢桿菌生長模型的建立[J]. 上海預防醫學, 2017, 29(6): 431-434. DOI:10.19428/j.cnki.sjpm.2017.06.004.
[23] 桂萌, 章志超, 劉恩歧, 等. 托盤包裝鱘魚中腐敗希瓦氏菌和總菌的生長動力學及貨架期預測[J]. 食品工業科技, 2016, 37(20): 331-335;40. DOI:10.13386/j.issn1002-0306.2016.20.057.
[24] 周桂香, 陳林, 張叢志, 等. 溫度水分對秸稈降解微生物群落功能多樣性影響[J]. 土壤, 2015, 47(5): 911-918. DOI:10.13758/j.cnki.tr.2015.05.016.
[25] 樂毅全, 王士芬, 朱核光. 脫色菌腐敗希瓦氏菌的分離及其脫色性能研究[J]. 環境科學與技術, 2004, 27(3): 14-15. DOI:10.3969/j.issn.1003-6504.2004.03.006.
[26] XU J, HE W, WANG Z, et al. A comparison of molecular biology mechanism of Shewanella putrefaciens between fresh and terrestrial sewage waste water[J]. Frontiers in Bioengineering and Biotechnology,2016, 4: 86. DOI:10.3389/fbioe.2016.00086.
[27] 許玫英, 郭俊, 鐘小燕, 等. 一個降解染料的希瓦氏菌新種: 中國希瓦氏菌[J]. 微生物學報, 2004, 44(5): 561-566. DOI:10.3321/j.issn:0001-6209.2004.05.001.
[28] IVANOVA E P, NEDASHKOVSKAYA O I, ZHUKOVA N V,et al. Shewanella waksmanii sp. nov., isolated from a sipuncula(Phascolosoma japonicum)[J]. International Journal of Systematic and Evolutionary Microbiology, 2003, 53(Pt 5): 1471-1477. DOI:10.1099/ijs.0.02630-0.
[29] IVANOVA E P, SAWABE T, HAYASHI K, et al. Shewanella fi delis sp.nov., isolated from sediments and sea water[J]. International Journal of Systematic and Evolutionary Microbiology, 2003, 53(Pt 2): 577-582.DOI:10.1099/ijs.0.02198-0.
[30] 卜蒙蒙. 我國近海海藻表面附著細菌的多樣性及其產酶、抑菌能力研究[D]. 青島: 青島科技大學, 2015.
Carbon Utilization Prof i le and Growth Kinetics of Shewanella putrefaciens Isolated from Spoiling Fish
XIU Yanhui1,2, GUO Quanyou1,*, HAN Chengyi3, CHEN Qingrong4
(1. East China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Shanghai 200090, China;2. College of Food Science and Technology, Shanghai Ocean University, Shanghai 201306, China;3. Ningde Yifeng Aquatic Product Co. Ltd., Ningde 352100, China;4. Ningde Jiaocheng Aquatic Product Technology Promotion Department, Ningde 352100, China)
In order to investigate the carbon source utilization pro fi le of Shewanella putrefaciens at different temperatures,a kinetic model describing the growth of S. putrefaciens in multi-well plates at 5, 15, 25 and 33 ℃ was developed using modified Gompertz equation and the kinetic parameters maximum specific growth rate (μmax) and lag phase duration (λ)were obtained. Moreover, based on average well color development (AWCD) and utilization area, the carbon utilization rate of S. putrefaciens was analyzed. The results showed that S. putrefaciens presented the highest μmaxand lowest λ at 25 ℃.Moreover, at this temperature, the highest carbon utilization and cell viability were observed followed by growth at 15 ℃;the cell viability at 5 ℃ was one-sixth of that at 25 ℃, and the lowest cell viability was found at 33 ℃. Temperature in the range of 5–25 ℃ exhibited a good linear relationship withandThe utilization rates of of carbohydrates and carboxylic acids were 30% and 29% at 5–33 ℃, respectively. Carbohydrates (dextrin, D-maltose, α-D-glucose, D-arabinol and stachyose), carboxylic acids (L-malic acid, L-lactic acid and acetoacetic acid), amino acids (L-serine, amino-acetyl-L-proline, L-aspartic acid and L-glutamic acid), fatty acids/lipids, amines/amides and other substances (gelatin, methyl pyruvate, Tween 40 and L-histamine) could be better utilized by the strain.
Shewanella putrefaciens; carbon; growth kinetics; multi-well plate
10.7506/spkx1002-6630-201802013
TS254.4
A
1002-6630(2018)02-0080-08
修艷輝, 郭全友, 韓承義, 等. 魚源腐敗希瓦氏菌碳源利用及動力學分析[J]. 食品科學, 2018, 39(2): 80-87.
10.7506/spkx1002-6630-201802013. http://www.spkx.net.cn
2017-01-17
國家自然科學基金面上項目(31371867);上海市自然科學基金項目(16ZR1444900);
中國水產科學研究院東海水產研究所基本科研業務費項目(2014G02;2011M04)
修艷輝(1990—),女,碩士研究生,研究方向為水產品加工與安全保障。E-mail:xiuyanhui2014@163.com*通信作者簡介:郭全友(1974—),男,副研究員,博士,研究方向為水產品加工與安全保障。E-mail:dhsguoqy@163.com
XIU Yanhui, GUO Quanyou, HAN Chengyi, et al. Carbon utilization prof i le and growth kinetics of Shewanella putrefaciens isolated from spoiling fi sh[J]. Food Science, 2018, 39(2): 80-87. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-201802013. http://www.spkx.net.cn
猜你喜歡
中等數學(2022年2期)2022-06-05 07:10:50
中學生數理化·七年級數學人教版(2021年11期)2021-12-06 05:38:48
中學生數理化(高中版.高考數學)(2021年6期)2021-07-28 06:19:08
小學生學習指導(低年級)(2020年6期)2020-07-25 02:31:36
小學生學習指導(低年級)(2019年11期)2019-11-25 07:31:44
小學生學習指導(低年級)(2018年9期)2018-09-26 05:59:44
瘋狂英語·新讀寫(2018年2期)2018-09-07 09:32:10
數學小靈通·3-4年級(2017年6期)2017-06-22 11:28:50
工業設計(2016年5期)2016-05-04 04:00:33
河北遙感(2015年4期)2015-07-18 11:05:06
