內(nèi)源酶對(duì)刺參腸自溶的作用
2018-01-04 05:43:24李傲婷杜椅楠段秀紅柴曉倩于翠平吳海濤
食品科學(xué) 2018年2期
李傲婷,杜椅楠,段秀紅,柴曉倩,吳 超,唐 越,于翠平,吳海濤*
內(nèi)源酶對(duì)刺參腸自溶的作用
李傲婷,杜椅楠,段秀紅,柴曉倩,吳 超,唐 越,于翠平,吳海濤*
(大連工業(yè)大學(xué)食品學(xué)院,國(guó)家海洋食品工程技術(shù)研究中心,遼寧 大連 116034)
以刺參腸為原料,通過誘導(dǎo)自溶并添加各種酶抑制劑,以三氯乙酸可溶性寡肽和還原糖釋放量為指標(biāo),并采用聚丙烯酰胺凝膠電泳檢測(cè)方法,研究?jī)?nèi)源酶在刺參腸自溶過程中的作用。結(jié)果表明,在一定的濃度范圍內(nèi),胃蛋白酶抑制劑、胰蛋白酶抑制劑、絲氨酸蛋白酶抑制劑、α-淀粉酶抑制劑和β-1,3-葡萄糖苷酶抑制劑對(duì)刺參腸的自溶均具有一定的抑制作用,且絲氨酸蛋白酶抑制劑N-甲苯磺酰-L-賴氨酸氯甲基化酮作用較強(qiáng)。說明胃蛋白酶、胰蛋白酶、絲氨酸蛋白酶、α-淀粉酶和β-1,3-葡萄糖苷酶參與刺參腸自溶過程。
刺參;腸;自溶;內(nèi)源酶;絲氨酸蛋白酶
海參屬棘皮動(dòng)物門海參綱,是一種重要的海洋軟體動(dòng)物[1]。近年來海參養(yǎng)殖業(yè)在亞洲國(guó)家發(fā)展迅速[2]。2015年,我國(guó)海參產(chǎn)量達(dá)20.50萬 t[3],而刺參(Stichopus japonicus)是我國(guó)最重要的人工養(yǎng)殖經(jīng)濟(jì)品種之一[4]。刺參具有極強(qiáng)的自溶能力,在受到外界物理因素、化學(xué)因素和生理因素等刺激后,原本正常的生命活動(dòng)出現(xiàn)紊亂,會(huì)發(fā)生吐腸和體壁軟化的現(xiàn)象[5]。刺參體內(nèi)含有豐富的內(nèi)源酶,主要為體壁和腸中的復(fù)雜酶類[6-7]。鄭杰等[8]采用環(huán)境因子紫外線誘導(dǎo)海參自溶,在考察溫度、pH值、NaCl質(zhì)量濃度和液料比對(duì)海參自溶過程中主要化學(xué)成分變化影響的基礎(chǔ)上,建立了海參自溶的評(píng)價(jià)指標(biāo),即三氯乙酸(trichloroacetic acid,TCA)可溶性寡肽和還原糖釋放量。
刺參腸作為刺參加工的副產(chǎn)物,其中含有胃蛋白酶和胰蛋白酶[9]、組織蛋白酶B[10]、高堿性蛋白酶[11]、肽水解酶[12]及β-1,3-葡聚糖酶[13]等多種酶類。這些蛋白酶的存在為利用自溶手段制備生物活性寡肽創(chuàng)造了良好的條件,目前已成功獲得具有抗氧化活性的刺參腸自溶寡肽[14-15]。同時(shí),前期研究針對(duì)刺參體壁的自溶過程,研究了蛋白酶對(duì)其的貢獻(xiàn)作用[16]。然而,針對(duì)刺參腸自溶過程中,其內(nèi)源酶對(duì)腸蛋白質(zhì)作用的影響,目前還鮮見報(bào)道。
因此,本實(shí)驗(yàn)以刺參腸為原料,利用胃蛋白酶抑制劑(Pepstain)、胰蛋白酶抑制劑(soybean trypsin inhibitor,SBTI)、絲氨酸蛋白酶抑制劑及糖苷酶抑制劑,以TCA可溶性寡肽和還原糖釋放量為指標(biāo),并采用聚丙烯酰胺-凝膠電泳(sodium-dodecyl sulfatepolyacrylamide gel electrophoresis,SDS-PAGE)檢測(cè)刺參腸蛋白質(zhì)的變化情況,明確各種內(nèi)源酶在刺參腸自溶過程中的作用,為深入開發(fā)利用刺參腸提供理論參考依據(jù)。
1 材料與方法
1.1 材料、試劑與儀器
刺參腸取自新鮮刺參購于大連長(zhǎng)興市場(chǎng),取出刺參腸,去除內(nèi)容物,去離子水清洗后于-80 ℃冷凍。
Pepstain、SBTI、N-甲苯磺酰-L-賴氨酸氯甲基化酮(N-tosyl-L-lysine chloromethyl ketone,TLCK)、N-甲苯磺基-L-苯乙胺酰氯甲基酮(N-tosyl-L-phenylalaninylchloromethyl ketone,TPCK)、N,N,N,N-四甲基乙二胺(N,N,N’,N’-tetramethylethylenediamine,TEMED)、丙烯酰胺、N,N-亞甲基雙丙烯酰胺、β-巰基乙醇、考馬斯亮藍(lán)R-250、Folin-酚溶液 生工生物工程(上海)有限公司;即用型蛋白分子質(zhì)量標(biāo)準(zhǔn)(高)、即用型蛋白分子質(zhì)量標(biāo)準(zhǔn)(低) 寶生物工程(大連)有限公司;α-Amylase 、Nojirimycin 美國(guó)Sigma公司;TCA、DNS試劑、甘氨酸 天津市科密歐化學(xué)試劑有限公司;其他試劑均為分析純。
PHS-3精密pH計(jì) 上海雷磁儀器廠;AE-6401垂直電泳儀槽 日本ATTO公司;HH-S型水浴鍋 鞏義市予華儀器有限責(zé)任公司;CF16RXII離心機(jī) 日本株式會(huì)社日立制作所;XW-80A旋渦振蕩器 上海精科實(shí)業(yè)有限公司;Infinite200 NANO酶標(biāo)定量測(cè)定儀 瑞士Tecan公司;UV-5200型紫外-可見分光光度計(jì) 上海元析儀器有限公司;TS.B-108往復(fù)式脫色搖床 海門市其林貝爾儀器制造有限公司;MF-Chemi BIS 2.0凝膠成像儀以色列DNR成像系統(tǒng)有限公司。
1.2 方法
1.2.1 刺參腸自溶的誘導(dǎo)
根據(jù)前期優(yōu)化的刺參腸自溶最優(yōu)條件[15],將置于冰上緩化的刺參腸勻漿處理后,經(jīng)58 μW/cm2紫外線照射30 min,于48 ℃誘導(dǎo)自溶3 h,采用McIIvaine’s(0.2 mol/L)緩沖體系(pH 2.2、4.4或8.0),進(jìn)行蛋白酶抑制劑對(duì)刺參腸自溶中蛋白質(zhì)降解及TCA可溶性寡肽釋放的影響研究,并采用水相體系(自然pH值)研究糖苷酶抑制劑對(duì)刺參腸自溶中還原糖釋放量的影響作用。
1.2.2 蛋白酶抑制劑對(duì)刺參腸蛋白質(zhì)降解作用的影響
根據(jù)Laemmli[17]的方法并作適當(dāng)修改。取1.2.1節(jié)所得經(jīng)紫外照射或未經(jīng)照射的刺參腸勻漿液300 μL,加入900 μL的200 mmol/L McIIvaine’s緩沖溶液(Pepstain pH 2.2、SBTI pH 8.0、絲氨酸蛋白酶抑制劑pH 4.4)后,于48 ℃孵育3 h。加入等體積的5×上樣緩沖溶液(8 mol/L尿素、5% SDS、5%巰基乙醇、250 mmol/L pH 7.5 Tris-HCl),煮沸5 min后樣品于搖床過夜。10 000×g離心10 min后,取上清液10 μL,進(jìn)行SDS-PAGE分析,濃縮膠質(zhì)量濃度5 g/100 mL,分離膠質(zhì)量濃度15 g/100 mL,每塊膠濃縮膠電流8 mA,分離膠電流15 mA,電泳緩沖液采用SDS-Tris-甘氨酸體系。電泳完畢后,利用考馬斯亮藍(lán)R-250往復(fù)振蕩染色,采用凝膠成像儀進(jìn)行成像。以未自溶的刺參腸組為空白對(duì)照,以未添加蛋白酶抑制劑的自溶組為陽性對(duì)照。
1.2.3 蛋白酶抑制劑對(duì)刺參腸自溶中TCA可溶性寡肽釋放的影響
利用Lowery法測(cè)定TCA可溶性寡肽含量[18]。取1.2.2節(jié)所得自溶或未自溶的刺參腸樣品煮沸5 min后,加入等體積的20%的TCA溶液,靜置20 min,經(jīng)12 000×g離心10 min,取100 μL稀釋一定倍數(shù)的上清液,加入Folin-酚甲液500 μL,振蕩后靜置10 min,再加入Folin-酚乙液50 μL,立即振蕩后靜置30 min,500 nm波長(zhǎng)處測(cè)定吸光度。甲液由2 g/100 mL碳酸鈉溶液及0.4 g/100 mL氫氧化鈉混合溶液-2 g/100 mL酒石酸鉀鈉溶液-1 g/100 mL硫酸銅(100∶1∶1,V/V)溶液混合,現(xiàn)用現(xiàn)配,乙液為Folin-酚溶液。以牛血清蛋白為標(biāo)準(zhǔn)品獲得標(biāo)準(zhǔn)曲線y=0.629x+0.006 (R2= 0.998)。以未自溶的新鮮刺參腸組為空白對(duì)照,以未添加蛋白酶抑制劑的自溶組為陽性對(duì)照。根據(jù)標(biāo)準(zhǔn)曲線計(jì)算TCA可溶性寡肽含量,設(shè)空白組樣品為數(shù)值為1,計(jì)算TCA可溶性寡肽相對(duì)含量。
1.2.4 糖苷酶抑制劑對(duì)刺參腸自溶中還原糖釋放的影響
利用3,5-二硝基水楊酸法測(cè)定還原糖[19],并稍作修改。取1.2.1節(jié)所得經(jīng)紫外照射或未經(jīng)照射的刺參腸勻漿液300 μL,加入900 μL去離子水,加入淀粉酶抑制劑α-Amylase及β-1,3-葡聚糖酶抑制劑Nojirimycin于50 ℃孵育3 h。經(jīng)12 000×g離心10 min后取上清液500 μL,加入DNS試劑500 μL,于沸水浴中加熱5 min,取出后立即浸入冰上冷卻至室溫,搖勻,在540 nm波長(zhǎng)處測(cè)定吸光度。以葡萄糖為標(biāo)準(zhǔn)品繪制標(biāo)準(zhǔn)曲線y=0.000 9x-0.031(R2=0.998 4)。以未自溶的新鮮刺參腸組為空白對(duì)照,以未添加糖苷酶抑制劑的自溶組為陽性對(duì)照。根據(jù)標(biāo)準(zhǔn)曲線計(jì)算還原糖的釋放量,設(shè)空白組樣品的還原糖含量為1,計(jì)算還原糖的相對(duì)含量。
1.2.5 數(shù)據(jù)處理及統(tǒng)計(jì)學(xué)分析
2 結(jié)果與分析
2.1 胃蛋白酶對(duì)刺參腸自溶的影響
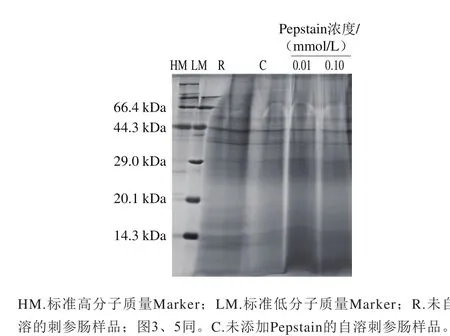
圖1 Pepstain對(duì)刺參腸自溶過程中蛋白質(zhì)降解作用的影響Fig. 1 Effects of pepstatin on proteolysis during autolysis of sea cucumber guts
利用SDS-PAGE檢測(cè)pH 2.2條件下不同濃度的Pepstain對(duì)刺參腸蛋白質(zhì)降解的影響作用,結(jié)果如圖1所示。刺參腸本身所含蛋白質(zhì)較多,約為干質(zhì)量的70.9%[15],未自溶樣品電泳條帶呈現(xiàn)彌散狀。經(jīng)自溶誘導(dǎo)后條帶整體變淺。Pepstain在0.01 mmol/L和0.1 mmol/L這兩種濃度下對(duì)刺參腸蛋白質(zhì)的降解作用有一定抑制,表現(xiàn)為29 kDa以下彌散條帶加深。這表明胃蛋白酶參與刺參腸自溶過程中蛋白質(zhì)的降解。有研究表明,蛋白酶的活性受pH值的影響很大,硬骨魚類的胃蛋白酶只有在強(qiáng)酸中才能有效地分解蛋白質(zhì)[20-21],一般最適pH值在2.0~3.0之間[22]。刺參腸中含有酸性蛋白酶,F(xiàn)u Xueyan等[11]利用SDS-PAGE法證明刺參腸中的酸性蛋白酶最適pH值在2.0~5.0之間。前期研究顯示刺參腸在pH 4.4條件下,自溶效果最好,在本研究顯示刺參腸在pH 2.2條件下,蛋白質(zhì)降解程度并不強(qiáng)烈,這與前期發(fā)現(xiàn)一致。
為進(jìn)一步明確Pepstain對(duì)刺參腸自溶的抑制作用,在pH 2.2條件下,進(jìn)一步考察0.01 mmol/L和0.1 mmol/L的Pepstain對(duì)刺參腸自溶過程中TCA可溶性寡肽釋放量的影響,如圖2所示。在pH 2.2條件下,刺參腸經(jīng)自溶誘導(dǎo)后,其TCA可溶性寡肽含量顯著提高(P<0.05),向自溶樣品中加入一定濃度的Pepstain后,與自溶組相比,TCA可溶性寡肽的含量顯著降低(P<0.05)。且隨著濃度提高抑制效果加強(qiáng),結(jié)合SDS-PAGE檢測(cè)結(jié)果,說明胃蛋白酶參與刺參腸自溶過程。
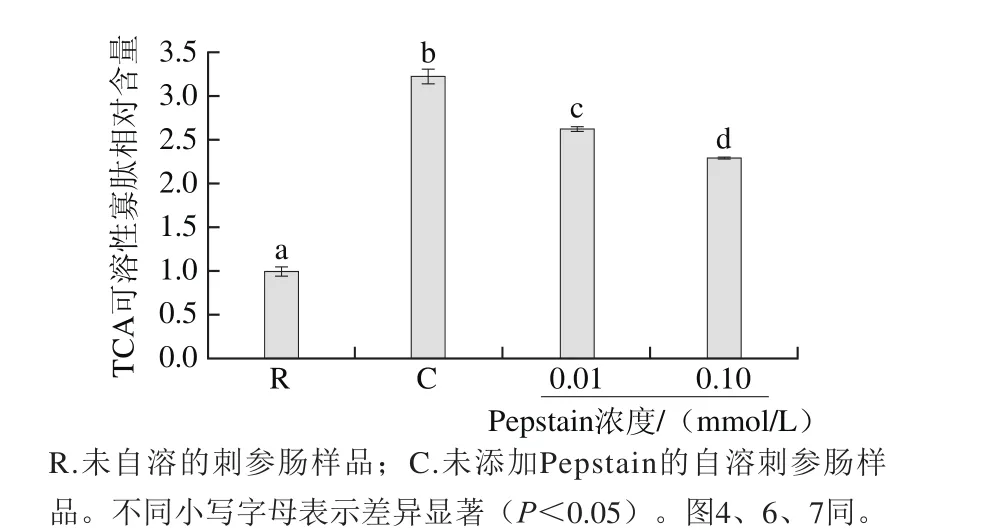
圖2 Pepstain對(duì)刺參腸自溶過程中TCA可溶性寡肽釋放的影響(pH 2.2)Fig. 2 Effects of pepstatin on the release of TCA-soluble oligopeptide during autolysis of sea cucumber guts at pH 2.2
2.2 胰蛋白酶對(duì)刺參腸自溶的影響
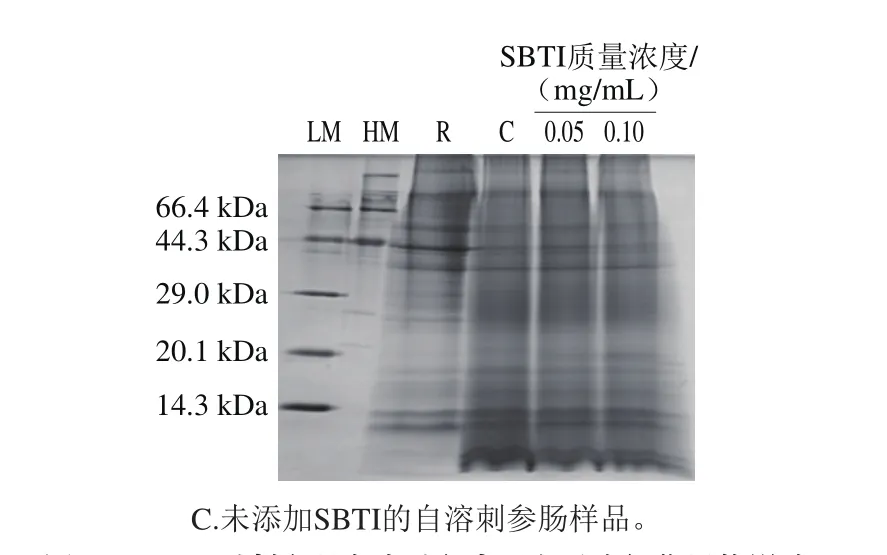
圖3 SBTI對(duì)刺參腸自溶過程中蛋白質(zhì)降解作用的影響Fig. 3 Effects of trypsin inhibitor on proteolysis during autolysis of sea cucumber guts
采用SDS-PAGE分析pH 8.0條件下SBTI對(duì)刺參腸自溶過程中蛋白質(zhì)降解的影響,結(jié)果見圖3。經(jīng)自溶誘導(dǎo)后,與未自溶樣品對(duì)比,44.3 kDa以上的蛋白質(zhì)降解明顯,29 kDa以下產(chǎn)生很多的降解產(chǎn)物。在pH 8.0的自溶環(huán)境下,SBTI在0.05 mg/mL質(zhì)量濃度下對(duì)刺參腸蛋白質(zhì)的降解僅有略微的抑制作用,表現(xiàn)為44.3 kDa以上的蛋白質(zhì)條帶與自溶樣品相比僅略微加深。大量研究表明水產(chǎn)動(dòng)物如虹鱒(Oncorhynchus mykiss)[23]、金槍魚(Thunnus albacores)[24]及嘉陵江鲇[25]等,其胰蛋白酶在pH 8.0條件下活性較高,本實(shí)驗(yàn)結(jié)果說明,在pH 8.0條件下胰蛋白酶對(duì)刺參腸的自溶影響微弱。
為進(jìn)一步明確SBTI對(duì)刺參腸自溶的抑制效果,考察pH 8.0條件下,0.05 mg/mL和0.1 mg/mL質(zhì)量濃度的SBTI對(duì)刺參腸自溶過程中TCA可溶性寡肽釋放量的影響,結(jié)果如圖4所示。在pH 8.0條件下,刺參腸經(jīng)自溶誘導(dǎo)后,TCA可溶性寡肽含量顯著提高(P<0.05),TCA可溶性寡肽的相對(duì)含量?jī)H為2.68±0.03,小于pH 2.2酸性條件下的TCA可溶性寡肽的相對(duì)含量為3.33±0.07,說明酸性條件更有利于刺參腸的降解。向自溶樣品中加入一定濃度的SBTI后,與自溶組相比,TCA可溶性寡肽的含量顯著降低(P<0.05),但兩種濃度作用基本相同(P>0.05),綜合SDS-PAGE分析和TCA可溶性寡肽兩項(xiàng)指標(biāo),表明胰蛋白酶雖參與刺參腸的自溶,但作用微弱。
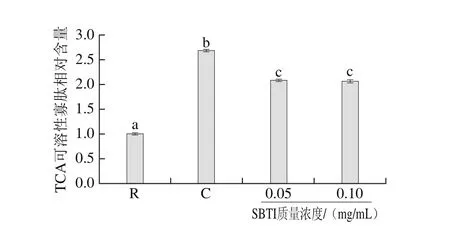
圖4 SBTI對(duì)刺參腸自溶過程中TCA可溶性寡肽釋放的影響Fig. 4 Effects of trypsin inhibitor on the release of TCA-soluble oligopeptide during autolysis of sea cucumber guts
2.3 絲氨酸蛋白酶抑制劑對(duì)刺參腸自溶的影響

圖5 絲氨酸蛋白酶抑制劑對(duì)刺參腸自溶過程中蛋白質(zhì)降解作用的影響Fig. 5 Effects of serine protease inhibitors on proteolysis during autolysis of sea cucumber guts
Yan Longjie等[26]成功從海參腸中純化得到一種絲氨酸蛋白酶,由此可見海參腸中含有含量豐富的絲氨酸蛋白酶,利用SDS-PAGE分析pH 4.4條件下不同濃度的絲氨酸蛋白酶抑制劑,TLCK、TPCK對(duì)刺參腸蛋白質(zhì)降解的影響作用,如圖5所示。經(jīng)自溶誘導(dǎo)后,與未自溶樣品對(duì)比刺參腸蛋白質(zhì)降解劇烈。在pH 4.4的自溶條件下,絲氨酸蛋白酶抑制劑TPCK在0.1 mmol/L和1 mmol/L兩種濃度下對(duì)刺參腸蛋白質(zhì)的降解均有一定的抑制作用,表現(xiàn)為分子質(zhì)量大于29.0 kDa及小于20.1 kDa的蛋白質(zhì)的條帶加深,與TPCK相似,TLCK對(duì)自溶刺參腸的蛋白質(zhì)降解有明顯的抑制作用,上述結(jié)果說明,絲氨酸蛋白酶參與刺參腸自溶過程中。

圖6 絲氨酸蛋白酶抑制劑對(duì)刺參腸自溶過程中TCA可溶性寡肽釋放的影響Fig. 6 Effects of serine protease inhibitors on the release of TCA-soluble oligopeptide during autolysis of sea cucumber guts
考察pH 4.4條件下,0.1 mmol/L和1 mmol/L濃度下的兩種絲氨酸蛋白酶抑制劑TLCK、TPCK對(duì)刺參腸自溶過程中TCA可溶性寡肽釋放量的影響作用,如圖6所示。在pH 4.4條件下,刺參腸經(jīng)自溶誘導(dǎo)后,TCA可溶性寡肽含量顯著提高(P<0.05),其相對(duì)含量可達(dá)到3.98±0.31,強(qiáng)于在pH 2.2和pH 8.0條件下的刺參腸TCA可溶性寡肽相對(duì)含量,這與前期發(fā)現(xiàn)一致[15]。向自溶樣品中加入一定濃度的絲氨酸蛋白酶抑制劑后,與自溶組相比,TCA可溶性寡肽的含量顯著降低(P<0.05),且TLCK的作用效果強(qiáng)于TPCK。均與SDS-PAGE檢測(cè)結(jié)果一致。上述結(jié)果說明,絲氨酸參與刺參腸的自溶,相比于胃蛋白酶和胰蛋白酶,其作用效果較強(qiáng)。研究發(fā)現(xiàn),刺參腸的天然pH值為6.0±0.02,呈現(xiàn)弱酸性,由于消化酶的活性受pH值影響較大,因此本實(shí)驗(yàn)在應(yīng)用各種蛋白酶抑制劑時(shí),對(duì)于消化酶選擇其最適pH值條件下進(jìn)行,而對(duì)于絲氨酸蛋白酶,選擇在刺參腸最適的pH值下進(jìn)行,這可能導(dǎo)致不同抑制劑對(duì)刺參腸自溶作用不同的原因。
2.4 糖苷酶對(duì)刺參腸自溶中還原糖釋放的影響
α-淀粉酶抑制劑是屬于糖苷水解酶抑制劑中的一種,它能有效地抑制淀粉酶的活性[27]。β-1,3-葡聚糖酶抑制劑是一類新的抗真菌抗生素,它通過抑制葡聚糖合成酶,而干擾細(xì)胞壁的合成,從而產(chǎn)生抗真菌作用[28]。β-1,3-葡聚糖苷酶主要分布于海洋無脊椎動(dòng)物動(dòng)物中,有研究表明該酶參與海洋無脊椎動(dòng)物對(duì)藻類食物的消化,并在其胚胎發(fā)育過程中發(fā)揮著重要的作用[29-30]。為明確糖苷酶抑制劑對(duì)刺參腸自溶中還原糖釋放量的影響,分別考察自然pH值條件下,0.01 mmol/L和0.05 mmol/L兩種濃度的糖苷酶抑制劑,α-Amylase、Nojirimycin的作用,如圖7所示。自然pH值條件下,刺參腸經(jīng)自溶誘導(dǎo)后,還原糖釋放量顯著提高(P<0.05),這與前期結(jié)果一致[8],向自溶樣品中加入一定濃度的β-1-3-葡聚糖酶抑制劑和α-淀粉酶抑制劑后,與自溶組相比,自溶過程中的溶出的還原糖明顯下降(P<0.05),上述結(jié)果表明淀粉酶和β-1,3葡萄糖苷酶均參與了刺參腸的自溶過程。
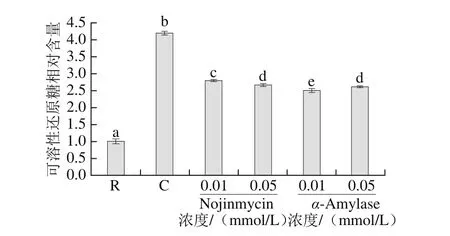
圖7 糖苷酶抑制劑對(duì)刺參腸自溶過程中還原糖釋放量的影響Fig. 7 Effects of glycosidase inhibitors on the release of reducing sugar during autolysis of sea cucumber S. japonicus guts
3 結(jié) 論
刺參腸自溶后,其蛋白質(zhì)會(huì)發(fā)生明顯的降解,同時(shí)TCA可溶性寡肽和還原糖釋放量顯著提高。結(jié)合SDSPAGE檢測(cè),發(fā)現(xiàn)刺參腸自溶過程中,多種酶參與其中,胃蛋白酶、胰蛋白酶、絲氨酸蛋白酶、α-淀粉酶和β-1,3-葡萄糖苷酶均參與了刺參腸自溶過程。
[1] 朱蓓薇. 海珍品加工理論與技術(shù)的研究[M]. 北京: 科學(xué)出版社,2010(9): 3-5.
[2] NGO D H, WIJESEKARA I, VO T S, et al. Marine food-derived functional ingredients as potential antioxidants in the food industry:an overview[J]. Food Research International, 2011, 44(2): 523-529.DOI:10.1016/j.foodres.2010.12.030.
[3] 農(nóng)業(yè)部漁業(yè)局. 中國(guó)漁業(yè)年鑒[M]. 北京: 中國(guó)農(nóng)業(yè)出版社, 2016:57-58.
[4] ANDERSON S C, FLEMMING J M, WATSON R, et al. Serial exploitation of global sea cucumber fisheries[J]. Fish & Fisheries,2011, 12(3): 317-339. DOI:10.1111/j.1467-2979.2010.00397.x.
[5] ZHU B W, ZHENG J, ZHANG Z S, et al. Autophagy plays a potential role in the process of sea cucumber body wall “Melting” induced by UV irradiation[J]. Wuhan University Journal of Natural Sciences,2008, 13(2): 232-238. DOI:10.1007/s11859-008-0220-3.
[6] ZHAO C C, YANG Y, WU H T, et al. Characterization of proteolysis in muscle tissues of sea cucumber Stichopus japonicus[J]. Food Science and Biotechnology, 2016, 25(6): 1529-1535. DOI:10.1007/s10068-016-0237-x.
[8] 鄭杰, 吳海濤, 董秀萍, 等. 基于主要化學(xué)成分變化建立的海參自溶評(píng)價(jià)指標(biāo)[J]. 大連工業(yè)大學(xué)學(xué)報(bào), 2010, 29(6): 391-395.
[9] GAO F, YANG H S, XU Q, et al. Effect of water temperature on digestive enzyme activity and gut mass in sea cucumber Apostichopus japonicus (Selenka), with special reference to aestivation[J]. Chinese Journal of Oceanology and Limnology, 2009, 27(4): 714-722.DOI:10.1007/s00343-009-9202-3.
[10] SUN L M, ZHU B W, WU H T, et al. Purif i cation and characterization of Cathepsin B from the gut of the sea cucumber (Stichopus japonicas)[J].Food Science and Biotechnology, 2011, 20(4): 919-925. DOI:10.1007/s10068-011-0127-1.
[11] FU X Y, XUE C H, MIAO B C et a1. Study of a highly alkaline protease extracted from digestive tract of sea cucumber (Stichopus japonicus)[J]. Food Research International, 2005, 38(3): 323-329.DOI:10.1016/j.foodres.2004.09.014.
[12] MCGETTIGAN S, CANNING M, O’CUINN G, et al. Peptide hydrolases in holothurian intestinal mucosa[J]. Comparative Biochemistry and Physiology Part C: Comparative Pharmacology,1981, 69(1): 169-170. DOI:10.1016/0306-4492(81)90123-4.
[13] ZHU B W, ZHAO J G, YANG J F, et al. Purification and partial characterization of a novel β-1,3-glucanase from the gut of sea cucumber Stichopus japonicus[J]. Process Biochem, 2008, 43(10):1102-1106. DOI:10.1016/j.procbio.2008.06.001.
[14] 鄭杰, 吳海濤, 朱蓓薇, 等. 海參腸自溶水解物抗氧化活性的研究[J].大連工業(yè)大學(xué)學(xué)報(bào), 2011, 30(5): 313-317.
[15] ZHENG J, WU H T, ZHU B W, et al. Identif i cation of antioxidative oligopeptides derived from autolysis hydrolysates of sea cucumber(Stichopus japonicus) guts[J]. European Food Research and Technology,2012, 234(5): 895-904. DOI:10.1007/s00217-012-1708-9.
[16] WU H T, LI D M, ZHU B W, et al. Proteolysis of noncollagenous proteins in sea cucumber, Stichopus japonicus, body wall:characterisation and the effects of cysteine protease inhibitors[J].Food Chemistry, 2012, 141(2): 1287-1294. DOI:10.1016/j.foodchem.2013.03.088.
[17] LAEMMLI U K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4[J]. Nature, 1970, 227: 680-685.
[18] LOWRY, O H, ROSENBROUGH, N J, FAIR, A L, et al.Proteinmeasurement with the Folin-phenol reagents[J]. Journal of Biological Chemistry, 1951, 193(1): 265-275.
[19] 張惟杰. 糖復(fù)合物生化研究技術(shù)[M]. 杭州: 浙江大學(xué)出版社, 1999:10-12.
[20] VYS W, HECHT T. Assay on the digestive enzymes of sharp tooth catifish clarias gariepinus (Pisces: clariidae)[J]. Aquaculture, 1987,63(1/2/3/4): 301-310. DOI:10.1016/0044-8486(87)90080-9.
[21] VIELMA J, RUOHONEN K, LALL S P. Supplemental citric acid and particle size of fi sh bone-meal inf l uence the availability of mineraIs in rainbow trout Oncorhynehus mykiss (Walbaum)[J]. Aquaculture Nutrition, 1999, 5(1): 65-71. DOI:10.1046/j.1365-2095.1999.00092.x.
[22] 周景樣, 陳勇, 黃權(quán), 等. 魚類消化酶的活性及環(huán)境條件的影響[J].北京大學(xué)學(xué)報(bào), 2001, 2(1): 70-83.
[23] HIDALGO M C, UREA E, SANZ A. Comparative study of digestive enzymes in fish with different nutritional habits: photolytic and amylase activities[J]. Aquaculture, 1999, 170(3/4): 267-283.
[24] ALEJANDRO B J, CAMILO P, DANIEL M, et al. A preliminary study of digestive enzyme activities and amino acid composition of early juvenile yellowfin tuna (Thunnus albacaras)[J]. Aquaculture,2011, 312(4): 205-211. DOI:10.1016/j.aquaculture.2010.12.027.
[25] 向梟, 周興華, 陳建, 等. pH對(duì)嘉陵江鲇消化酶活性的影響[J]. 淡水漁業(yè), 2013, 43(4): 13-17.
[26] YAN L J, ZHAN C L, CAO M J, et al. Purif i cation, characterization,cDNA cloning and in vitro expression of a serine proteinase from the intestinal tract of a sea cucumber (Stichopus japonicus) with collagen degradation activity[J]. Journal of Agriculatural and Food Chemistry,2014, 62(20): 4769-4777. DOI:10.1021/jf500923y.
[27] 呂鳳霞, 陸兆新. α-淀粉酶抑制劑的研究進(jìn)展[J]. 食品科學(xué), 2002,23(3): 152-155.
[28] 顧覺奮, 張紹譚. β-1,3-葡聚糖合成酶抑制劑類抗真菌抗生素[J]. 藥品評(píng)價(jià), 2004, 1(4): 300-304.
[29] 趙軍崗, 董秀萍, 朱蓓薇, 等. 海參腸道β-1,3-葡聚糖酶的提取條件及其酶學(xué)性質(zhì)[J]. 大連輕工業(yè)學(xué)院學(xué)報(bào), 2007, 26(4): 289-294.
[30] KOZHEMYAKO V B, REBRIKOV D V, LUKYANOV S A, et al. Molecular cloning and characterization of an endo-1,3-β-D-glucanase from the mollusk Spisula sachalinensis[J]. Comparative Biochemistry and Physiology Part B, 2004, 137(2): 169. DOI:10.1016/j.cbpc.2003.10.018.
Effect of Endogenous Enzymes on Autolysis of Sea Cucumber Stichopus japonicus Guts
LI Aoting, DU Yinan, DUAN Xiuhong, CHAI Xiaoqian, WU Chao, TANG Yue, YU Cuiping, WU Haitao*
(National Engineering Research Center of Seafood, School of Food Science and Technology,Dalian Polytechnic University, Dalian 116034, China)
The effect of endogenous enzymes on the autolysis of sea cucumber (Stichopus japonicas) guts was studied by addition of various protease inhibitors. The autolysis was evaluated by the release of trichloroacetic acid (TCA)-soluble oligopeptides and reducing sugar, and detected by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE).The results indicated that different inhibitors, including pepsin inhibitor, trypsin inhibitor, serine protease inhibitor, amylase inhibitor and β-1,3-glucanase inhibitor showed inhibitory effects on the autolysis of S. japonicus guts in a certain range of concentration. The serine protease inhibitor N-tosyl-L-phenylalaninyl-chloromethyl ketone (TPCK) showed relatively higher inhibitory effect than the other inhibitors. These results suggest endogenous enzymes, including pepsin, trypsin, serine proteases, amylase and β-1,3-glucanase, may be involved in the autolysis of S. japonicus guts.
Stichopus japonicus; guts; autolysis; endogenous enzymes; serine proteases
10.7506/spkx1002-6630-201802014
TS254
A
1002-6630(2018)02-0088-05
李傲婷, 杜椅楠, 段秀紅, 等. 內(nèi)源酶對(duì)刺參腸自溶的作用[J]. 食品科學(xué), 2018, 39(2): 88-92.
10.7506/spkx1002-6630-201802014. http://www.spkx.net.cn
LI Aoting, DU Yinan, DUAN Xiuhong, et al. Effect of endogenous enzymes on autolysis of sea cucumber Stichopus japonicus guts[J]. Food Science, 2018, 39(2): 88-92. (in Chinese with English abstract) DOI:10.7506/spkx1002-6630-201802014. http://www.spkx.net.cn
2016-11-02
國(guó)家自然科學(xué)基金面上項(xiàng)目(31370037);大連市高層次人才創(chuàng)新支持計(jì)劃項(xiàng)目(2015R083)
李傲婷(1992—),女,碩士研究生,研究方向?yàn)槭称飞锛夹g(shù)。E-mail:756574359@qq.com
*通信作者簡(jiǎn)介:吳海濤(1980—),女,副教授,博士,研究方向?yàn)槭称飞锛夹g(shù)。E-mail:wht205@163.com
