蘋果樹腐爛病菌β-葡萄糖苷酶基因的克隆及表達分析
2018-01-12 00:42:22李保華梁文星王彩霞
華北農學報 2017年6期
李 婷,練 森,李保華,梁文星,王彩霞
(青島農業大學 植物醫學學院,山東省植物病蟲害綜合防控重點實驗室,山東 青島 266109)
由黑腐皮殼菌(Valsamalivar.mali,Vmm)引起的蘋果樹腐爛病是蘋果生產上的三大重要病害之一,該病在我國各蘋果產區發生普遍,可造成果樹主干及整樹死亡,甚至毀園[1-2]。2008年,全國范圍內蘋果樹腐爛病的總體發病率高達52.7%,且病情呈蔓延趨勢[3];2011年,煙臺蘋果產區腐爛病再次大發生,發病程度明顯重于一般年份[4]。現代蘋果產業技術體系病蟲害防控研究室的有關專家認為,我國蘋果生產正面臨腐爛病第5次發病高峰的威脅[2-3]。目前,由于缺乏對蘋果樹腐爛病菌致病機制的全面了解,導致該病尚無法得到有效控制。
蘋果樹腐爛病菌的主要致病機制,除產生毒素外,還可分泌一系列的細胞壁降解酶,其中包括木聚糖酶、纖維素酶、β-葡萄糖苷酶、多聚半乳糖醛酸酶和果膠甲基半乳糖醛酸酶[5-7]。臧睿[8]研究發現蘋果樹腐爛病菌不同菌株的致病力與其分泌的果膠酶活性關系密切,許春景等[7]報道腐爛病菌的果膠裂解酶基因Vmpl4及多聚半乳糖醛酸酶基因Vmpg7和Vmpg8被敲除后,突變體菌株的致病力均有不同程度的降低。蘋果樹腐爛病菌侵染致病過程中,分泌的5種細胞壁降解酶中以木聚糖酶的活性最高,且對蘋果組織的浸漬能力最強[5],史祥鵬[9]對腐爛病菌的木聚糖酶基因功能進行了研究,發現敲除Xylanase突變體菌株的致病力和產生的木聚糖酶活性均顯著降低。以上研究表明,果膠裂解酶、多聚半乳糖醛酸酶和木聚糖酶基因在腐爛病菌致病過程中具有重要作用。
β-葡萄糖苷酶能夠水解結合于末端非還原性β-D-葡萄糖苷鍵,釋放出β-D-葡萄糖和相應的配基,作為纖維素酶系的一個重要成員,β-葡萄糖苷酶在降解纖維素的過程中具有關鍵作用[10-13]。目前,已篩選到多種產β-葡萄糖苷酶的微生物菌株,如里氏木霉菌(Trichodermareesei)、木蹄層孔菌(Fomesfomentarius)和白色念珠菌(Candidaalbicans)等,并廣泛應用于食品、醫藥和飼料等領域[14-17]。研究表明,多種植物病原菌也可分泌高活性的β-葡萄糖苷酶,在其致病性中發揮重要作用。張大智等[18]報道細菌性角斑病菌(Xanthomonascampestris)接種杧果葉片后4 d,β-葡萄糖苷酶活性到達峰值是對照的4.4倍;金勤等[19]測定了炭疽菌(Collectotrichumgloeosporioide)接種油茶葉片后β-葡萄糖苷酶的活性變化,于接種后6 d酶活性水平最高。陳曉林等[5]對蘋果樹腐爛病菌分泌的β-葡萄糖苷酶活性進行了分析,發現隨腐爛病菌的侵染和擴展,β-葡萄糖苷酶活性持續升高,最大酶活性是對照的4.5倍。
隨著越來越多的β-葡萄糖苷酶基因被克隆和成功表達,為其基因結構和功能研究提供了重要依據[20-21]。Waksman[22]克隆了油菜菌核病菌(Sclerotiniasclerotiorum)的β-葡萄糖苷酶基因,并證實其原核表達蛋白具有β-葡萄糖苷酶活性。目前,有關其他植物病原菌β-葡萄糖苷酶基因的克隆及其在致病性中的作用機制研究還鮮有報道。本研究以蘋果樹腐爛病菌強致病力菌株LXS080601為材料,利用RT-PCR結合TA克隆技術獲得其β-葡萄糖苷酶基因序列,對其序列進行生物信息學分析,并通過實時熒光定量PCR技術測定該基因在腐爛病菌侵染致病過程中的表達,以期為明確β-葡萄糖苷酶基因在蘋果樹腐爛病菌致病性中的作用機理奠定基礎。
1 材料和方法
1.1 試驗材料
供試菌株和植物材料:蘋果樹腐爛病菌菌株LXS080601采集自山東省煙臺富士蘋果園發病枝干,經單菌絲分離后鑒定并保存。3-4年生富士蘋果(MalusdomesticaBorkh)幼樹栽植于青島農業大學試驗田。
試劑:TRIzol試劑,美國Invitrogen公司;PrimeScriptTMⅡ 1st StrandcDNA Synthesis Kit、Prime ScriptTMRT reagent Kit with gDNA Eraser試劑盒、SYBR Master Mix、Prime STAR Max DNA Polymerase、pMD18-T克隆載體,寶生物工程(大連)有限公司;DNA凝膠回收試劑盒、DNA Marker DL2000,青島擎科梓熙生物公司;大腸桿菌菌株DH5α,山東省植物病蟲害綜合防控重點實驗室保存;氨芐青霉素、Oligo(dT)18,生工生物工程(上海)股份有限公司。
培養基:馬鈴薯葡萄糖瓊脂(PDA)培養基:馬鈴薯200 g,葡萄糖20 g,瓊脂20 g,定容至1 000 mL;LB (Luria-Bertani)培養基:胰蛋白胨10 g,酵母抽提物5 g,氯化鈉5 g,定容至1 000 mL。
1.2 試驗方法
1.2.1 蘋果樹腐爛病菌總RNA提取與cDNA合成 蘋果樹腐爛病菌于PDA培養基中培養3 d后刮取菌絲,用TRIzol法提取總RNA。超微量分光光度計測定提取RNA的濃度,并用1.2%瓊脂糖凝膠電泳檢測RNA質量。以1 μg總RNA為模版,Oligo dT18為反轉錄引物,參照cDNA合成試劑盒說明書合成第一鏈cDNA,-20 ℃保存備用。
1.2.2 蘋果樹腐爛病菌VmGluI基因cDNA序列的克隆 根據本實驗室前期蘋果樹腐爛病菌基因組序列結果,利用Oligle 7軟件設計β-葡萄糖苷酶基因的上下游引物(VmGluIFwd:5′-ATGGCCCGGATAATTTCGATG-3′;VmGluIRev:5′-TTATGTCTCCTGGTATATGTTGAACATATT-3′)。以第一鏈cDNA為模版,PCR反應體系為25 μL:2×PrimeSTAR Max DNA Polymerase 12.5 μL,上下游引物各0.5 μL,cDNA 0.25 μL、ddH2O 11.25 μL。反應條件為:95 ℃預變性3 min;95 ℃變性10 s,55 ℃退火15 s,72 ℃延伸8 s,35個循環;72 ℃延伸10 min。PCR產物經1.0%的瓊脂糖凝膠電泳進行檢測,回收純化后連接至克隆載體pMD18-T,熱擊轉入大腸桿菌DH5α,篩選陽性克隆提取質粒,委托青島擎科梓熙生物技術有限公司進行測序。
1.2.3 蘋果樹腐爛病菌VmGluI序列的生物信息學分析 將克隆到的VmGluI序列在NCBI數據庫(http://blast.ncbi.nlm.nih.gov/Blast.cgi)中進行Blast驗證和同源性分析,并利用DNAMAN對蘋果樹腐爛病菌VmGluI和其他不同來源β-葡萄糖苷酶基因編碼氨基酸序列進行多重比對。采用軟件MEGA 5.1以鄰接法構建VmGluI蛋白分子系統發育樹。使用ExPASy生物信息學網站(http://web.expasy.org/protparam/)預測VmGluI編碼蛋白的理化性質,使用SOPMA(http://npsa-pbil.ibcp.fr/cgi-bin/npsautomat.pl)在線軟件進行蛋白質二級結構預測。
1.2.4 蘋果樹腐爛病菌VmGluI的表達分析 選取長勢良好且粗細一致的1-2年生富士蘋果枝條,用電烙鐵造成直徑約1~2 mm燙傷,活化培養3 d的腐爛病菌在菌落邊緣處打取菌餅,接種于枝條的燙傷處,用保鮮膜固定并保濕[4]。接種后6,12,24,48,72 h分別取病健交界處組織,提取樣品總RNA,利用Prime ScriptTMRT reagent Kit with gDNA Eraser試劑盒反轉錄獲得cDNA。根據VmGluI基因序列,設計合成定量PCR引物VmGluI-qF(5′-ACAAACAAGACATTGCTCGGAT-3′)和VmGluI-qR(5′-ATTATCTGCTCGTCCAACCAC-3′)。以真菌EF1α基因作為內參[23],合成引物EF1α-qF (5′-TGAGTTCGAGGCTGGTATCTCCAA-3′)和EF1α-qR (5′-TGTCCATCTTGTTGATGGCGACGA-3′)。反應體系20 μL:SYBR Premix Ex TaqTM(TliRNaseH Plus)10.0 μL,上下游引物各0.4 μL,cDNA模板1.0 μL,去離子8.2 μL。反應條件為:95 ℃預變性30 s;95 ℃變性5 s,60 ℃退火20 s,72 ℃延伸15 s,40個循環。上述反轉錄的cDNA梯度稀釋后作為模板,分別進行熒光定量PCR反應,繪制標準曲線,確定2對引物的擴增效率[24-25]。以PDA上培養的蘋果樹腐爛病菌作為對照,每個樣品重復3次,根據2-ΔΔCT法計算VmGluI在腐爛病菌侵染致病過程中的相對表達量[26]。
2 結果與分析
2.1 蘋果樹腐爛病菌VmGluI的克隆和序列分析
提取蘋果樹腐爛病菌LXS080601的總RNA,反轉錄成cDNA,以該cDNA為模版,用VmGluIFwd和VmGluIRev特異性引物進行PCR擴增,產物經1%瓊脂糖凝膠電泳檢測,約1 700 bp處有一特異性條帶(圖1)。進行克隆測序獲得1 689 bp的核苷酸序列,經預測該基因編碼562個氨基酸殘基,BlastN比對結果顯示,該基因序列與NCBI數據庫中的核苷酸序列相似性均不高;再用BlastP軟件將該基因編碼的氨基酸序列進行同源性比對,發現與蘋果(登錄號KUI68239.1)和梨樹(登錄號KUI56905.1)腐爛病菌的β-葡萄糖苷酶基因氨基酸序列相似性分別高達98.7%和87.8%。由此判斷,所克隆序列為蘋果樹腐爛病菌β-葡萄糖苷酶基因,命名為VmGluI,并將基因序列提交至GenBank數據庫(ID:KY646110)。
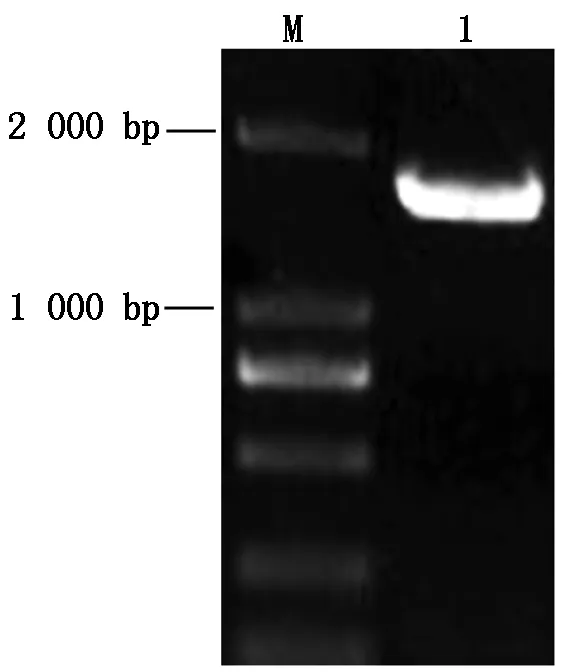
M.DNA分子標準;1.VmGluI PCR產物。M.DNA Marker DL2000;1.PCR product of VmGluI.
2.2 VmGluI編碼蛋白的理化性質和二級結構預測
利用Expasy Proteomics Server的在線軟件Protparam預測VmGluI編碼蛋白的理化性質,該蛋白由562個氨基酸組成,分子式為C2868H4253N715O847S15,相對分子質量62.7 kDa,理論等電點pI為4.64,脂肪系數79.29,不穩定系數38.02,預測為穩定蛋白。利用ProtScale進行蛋白質的疏水性預測,使用Hphob./Kyte&Doolittl算法,總平均親水性(Grand average of hydropathicity)為-0.162(圖2),表明VmGluI蛋白為親水蛋白。氨基酸組成中帶正電殘基(Arg+Lys)為34,帶負電殘基(Asp+Glu)為58,Gly (G)和Thr (T)使用頻率較高,而Cys (C)、His (H)和Met (M)使用頻率較低。此外,該基因編碼的氨基酸序列有信號肽,在24與25位氨基酸之間存在裂解位點。
利用SOMPA在線軟件對VmGluI蛋白的氨基酸序列的二級結構進行預測,發現VmGluI的二級結構由α-螺旋、β-轉角、延伸鏈和無規則卷曲構成(圖3),其中有562個氨基酸殘基組成的α-螺旋含量最高,占35.59%,其次為無規則卷曲,含量占34.70%,最少的是β-轉角,僅占9.79%,說明VmGluI蛋白的二級結構較穩定。對編碼區進行蛋白質結構功能域分析發現,VmGluI的87-555區段為糖基水解酶家族1功能域(Pfam:00232,E-value:4.75e-113),且存在酪蛋白激酶Ⅱ磷酸化位點、蛋白激酶C磷酸化位點、N-肉豆蔻酰化位點和N-糖基化位點。
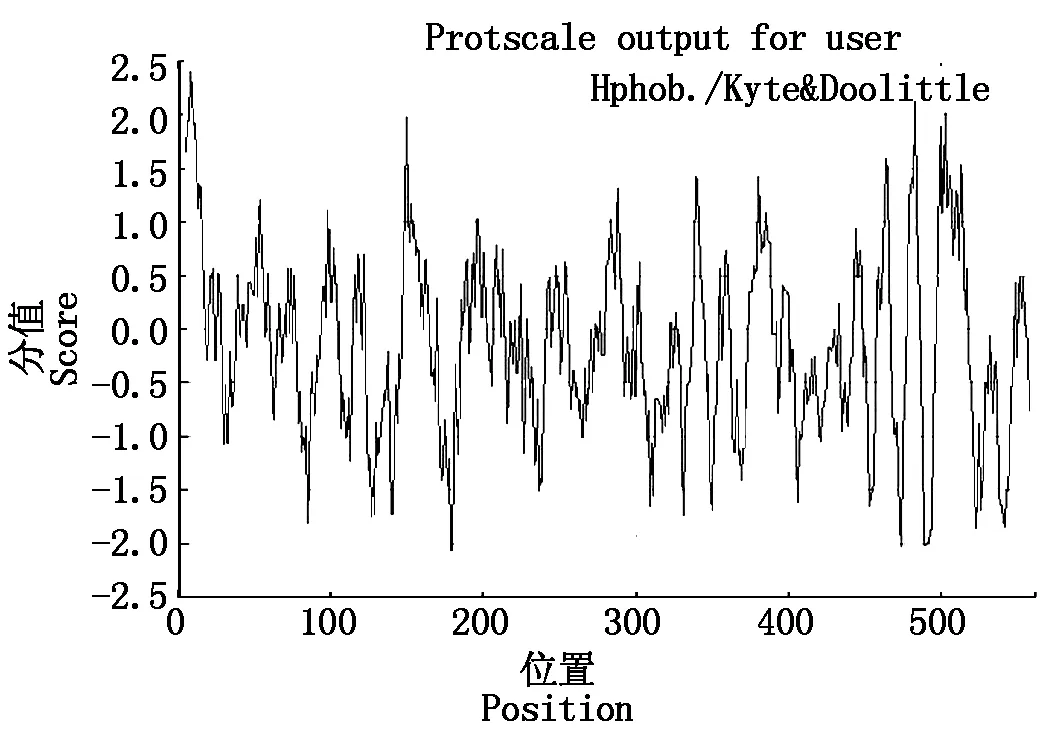
圖2 蘋果樹腐爛病菌VmGluI蛋白親水性/疏水性預測Fig.2 Predicted the hydrophobicity/hydrophilicity of VmGluI protein from Vmm

h.α-螺旋;e.延伸鏈;t.β-轉角;c.無規則卷曲。h.Alpha helix;e.Extended strand;t.Beta turn;c.Random coil.
2.3 VmGluI編碼氨基酸序列比對及系統發育分析
將VmGluI編碼的氨基酸序列與蘋果和梨樹腐爛病菌中β-葡萄糖苷酶基因氨基酸序列進行多重比對分析,圖4結果顯示,3條序列相似性高達94.2%,2個蘋果樹腐爛病菌β-葡萄糖苷酶基因氨基酸序列中有7個位點存在差異,其中6個位點集中在第235-304位氨基酸,而與梨樹腐爛病菌β-葡萄糖苷酶基因氨基酸序列的差異位點分布則較均勻。靠近N端的94位和靠近C端的532位谷氨酸Glu (G)是2個關鍵活性殘基,前者為親子供體,后者為親核試劑,與糖基水解酶家族1特有的保守氨基酸序列一致。
利用氨基酸BlastP比對功能進行同源性搜索,選取11條不同真菌來源的β-葡萄糖苷酶基因氨基酸序列,采用MEGA 5.1軟件中的鄰接法對包含VmGluI在內的12個β-葡萄糖苷酶基因氨基酸序列構建系統發育樹(圖5)。結果表明,蘋果樹腐爛病菌2個分離物β-葡萄糖苷酶基因所編碼的氨基酸序列同源性最高,與梨樹腐爛病菌(V.malivar.pyri,登錄號KUI56905.1)氨基酸序列親緣關系最近,并且與向日葵莖潰瘍病菌(Diaporthehelianthi,登錄號OCW40953.1)氨基酸序列相似性較高為71.0%,其聚在一個獨立分支上,Bootstrap值為100%,而與其他真菌β-葡萄糖苷酶基因同源關系較遠。白曲霉菌(Aspergilluskawachii,登錄號GAA82784.1)、黑曲霉菌(Aspergillusniger,登錄號XP 001401684.2)、青霉菌(Penicilliumsubrubescens,登錄號OKO99982.1)和短梗霉菌(Aureobasidiumnamibiae,登錄號XP 013431390.1)聚在一個大的獨立分支,與蘋果樹腐爛病菌VmGluI的進化關系最遠,可能與他們同屬于產β-葡萄糖苷酶真菌,而非植物病原真菌有關。

Vmm.蘋果樹腐爛病菌;Vmp.梨樹腐爛病菌;圖中黑色區域代表100%的相似度;灰色代表大于50%的相似度。Vmm.V.mali var.mali;Vmp.V.mali var.pyri;Blank areas indicate 100% similarity;Gray areas indicate at least 50% similarity.
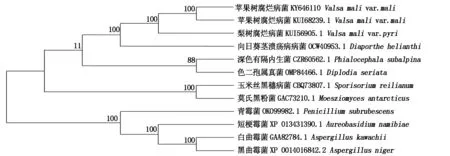
圖5 蘋果樹腐爛病菌與其他真菌來源β-葡萄糖苷酶基因氨基酸序列的系統進化樹Fig.5 Phylogenetic analysis of β-glucosidase genes from Vmm and other species
2.4 蘋果樹腐爛病菌侵染致病過程中VmGluI的表達分析
蘋果樹腐爛病菌LXS080601分別燙傷接種富士離體和活體枝條,接種后48~72 h可觀察到明顯的發病癥狀,隨接種時間延長,病斑不斷擴展。于接種后定期取樣,采用熒光定量PCR技術測定了蘋果樹腐爛病菌VmGluI表達量的時序變化。未接種蘋果樹腐爛病菌的枝條中無VmGluI的表達,接種病原菌的組織中均可檢測到VmGluI表達,且基因表達量存在顯著差異。圖6-A結果顯示,腐爛病菌接種離體枝條6~24 h后,VmGluI表達量相比PDA(0 h)對照無顯著差異,接種48 h后,基因表達量開始顯著升高為對照的3.4倍,接種72 h后,VmGluI表達量為對照6.3倍。蘋果樹腐爛病菌接種活體枝條后,VmGluI表達呈現先升高后降低再升高的趨勢(圖6-B)。接種6 h后,基因表達與PDA對照(0 h)無明顯差異,但接種12 h后,基因表達量已顯著升高為對照的11.2倍,之后表達量逐漸降低,于接種48 h后基因表達量出現低谷,但仍顯著高于對照;隨后,VmGluI表達水平快速升高,接種96 h后,基因表達水平量為對照的16.8倍,表明蘋果樹腐爛病菌在侵染離體和活體枝條過程中,VmGluI表達水平和時序變化存在明顯差異。
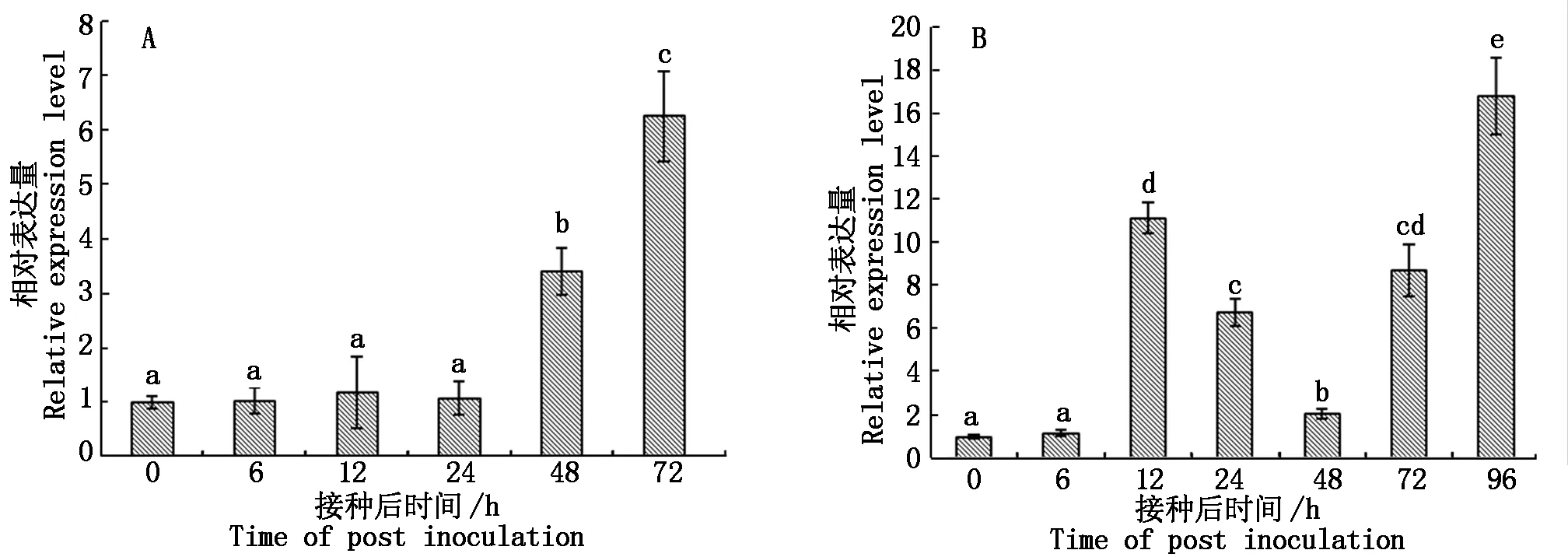
圖中不同字母表示經Duncan氏新復極差法檢驗在P<0.05水平差異顯著。Different letters indicate significant difference at P<0.05 level by Duncan′s multiple range test.
3 討論
β-葡萄糖苷酶能夠破壞植物細胞壁結構,使病原菌易于侵入和擴展,已證實該酶是蘋果樹腐爛病菌侵染致病過程中分泌的一種細胞壁降解酶,說明其在腐爛病菌致病中具有重要作用[5]。本研究以強致病力菌株LXS080601為材料,首次克隆了蘋果樹腐爛病菌β-葡萄糖苷酶基因VmGluI的cDNA序列,該基因開放閱讀框1 689 bp,編碼562個氨基酸,蛋白分子量62.7 kDa,房偉等[27]從海洋微生物中克隆到一個新型β-葡萄糖苷酶基因bgl1B,編碼449個氨基酸,蛋白分子量51.0 kDa,而陳輝等[28]克隆的斑玉蕈β-葡萄糖苷酶基因bgl1 cDNA全長2 574 bp,編碼857個氨基酸,說明不同微生物來源的β-葡萄糖苷酶基因片段長度存在較大差異。已知的大部分β-葡萄糖苷酶pI酸性范圍為3.5~5.5,VmGluI編碼的蛋白預測pI (4.64)也為酸性。經Protparam預測VmGluI編碼的蛋白為分泌型蛋白,1~24位氨基酸為信號肽序列,推測該基因對病原菌的侵染能力和特性具有重要作用[29]。
序列分析發現,VmGluI核苷酸序列與GenBank中提交序列相似性均不高,但其推導的氨基酸序列與蘋果和梨樹腐爛病菌β-葡萄糖苷酶基因氨基酸序列一致性分別為98.7%和87.8%,且與向日葵莖潰瘍病菌氨基酸序列相似性也高達71.0%,但這些序列均來自全基因組測序,未對β-葡萄糖苷酶基因進行克隆和序列分析。VmGluI系統進化樹分析表明,該基因編碼的氨基酸序列與蘋果和梨樹腐爛病菌、向日葵莖潰瘍病菌的親緣關系最近,聚在一個單獨的分支;而與其他非植物病原真菌β-葡萄糖苷酶基因氨基酸序列相似性較低,均不足40%,說明β-葡萄糖苷酶基因編碼蛋白在植物病原真菌中具有相對保守性,但不同物種間存在很大的變異,從而使得其能在不同生物物種中發揮獨特功能。
本研究發現,接種蘋果樹腐爛病菌的富士枝條組織中,VmGluI表達量顯著上調,表明在病原菌侵染過程中VmGluI起到了重要的作用,但該基因在離體和活體枝條中的表達水平和時序變化存在顯著差異。蘋果樹腐爛病菌接種離體枝條48 h后,VmGluI表達量開始顯著升高為對照的3.4倍,而活體枝條接種12 h后,基因表達水平已升高至對照的11.2倍,推測腐爛病菌侵染活體蘋果枝條要克服寄主的防御反應,需分泌更多的β-葡萄糖苷酶用于分解植物細胞壁。活體枝條接種腐爛病菌24~48 h后,VmGluI的表達量急劇下降,分析可能是寄主與腐爛病菌互作競爭時寄主的防御反應占優勢,抑制了VmGluI表達;接種72~96 h后,該基因表達量快速上升,說明腐爛病菌在寄主組織中已成功侵染,此時可觀察到明顯的發病癥狀。這與王彩霞等[30]和雍道敬等[31]的報道結果一致,富士愈傷組織和葉片接種蘋果樹腐爛病菌后1~2 d,多酚氧化酶、過氧化物酶等多種防御酶活性顯著升高并到達峰值,其可催化木質素和酚類氧化產物的形成,加厚細胞壁,形成阻止病原菌入侵的機械屏障,隨接種時間延長防御酶活性快速降低至對照水平。纖維素酶各酶組分單獨作用時效果不是很明顯,但當其協同作用時表現出很強的分解活性[32]。本研究也發現,接種腐爛病菌的蘋果枝條中,VmGluI的表達量顯著低于木聚糖基因[9],但VmGluI與其他纖維素酶基因表達是否存在協同作用,以及該基因的作用機制尚待進一步研究。
[1] 陳 策.蘋果樹腐爛病發生規律和防治研究[M].北京:中國農業科學技術出版社,2009.
[2] 李保華,王彩霞,董向麗.我國蘋果主要病害研究進展與病害防治中的問題[J].植物保護,2013,39(5):46-54.
[3] 曹克強,國立耘,李保華,等.中國蘋果樹腐爛病發生和防治情況調查[J].植物保護,2009,35(2):114-117.
[4] 王彩霞,董向麗,張振芳,等.2011年煙臺蘋果產區腐爛病發病情況調查與原因分析[J].農業科學與技術:英文版,2014,38(1):83-86.
[5] 陳曉林,牛程旺,李保華,等.蘋果樹腐爛病菌產生細胞壁降解酶的種類及其活性分析[J].華北農學報,2012,27(2):207-212.
[6] Shi X,Yu D,Kong Q,et al.Optimizing prokaryotic expression of a xylanasegene from the apple pathogenValsamalivar.mali[J].Advance Journal of Food Science&Technology,2015,9(9):701-705.
[7] 許春景,吳玉星,戴青青,等.蘋果樹腐爛病菌多聚半乳糖醛酸酶基因Vmpg7和Vmpg8的功能[J].中國農業科學,2016,49(8):1489-1498.
[8] 臧 睿.陜西蘋果樹腐爛病菌不同分離株生物學特性及致病性研究[D].楊凌:西北農林科技大學,2006.
[9] 史祥鵬.蘋果樹腐爛病菌木聚糖酶基因的克隆及功能分析[D].青島:青島農業大學,2016.
[10] Saloheimo M,Kujapanula J,Yl?sm?ki E,et al.Enzymatic properties and intracellular localization of the novelTrichodermareeseiβ-Glucosidase BGLII(CellA)[J].Applied & Environmental Microbiology,2002,68(9):4546-4553.
[11] Singhania R R,Patel A K,Sukumaran R K,et al.Role and significance of beta-glucosidases in the hydrolysis of cellulose for bioethanol production[J].Bioresource Technology,2013,127(1):500-507.
[12] Chen R Y,Baiya S,Sangkyu L,et al.Recombinant expression and characterization of the cytoplasmic rice β-glucosidase Os1BGlu4[J].PLoS One,2014,9(5):e96712.
[13] 陳 斌,錢 驊,閆淑珍,等.半夏內生真菌所產β-葡萄糖苷酶的酶學特性研究[J].中國野生植物資源,2016,35(2):7-9.
[14] Fang X,Shen Y,Zhao J,et al.Status and prospect of lignocellulosic bioethanol production in China[J].Bioresource Technology,2010,101(13):4814-4819.
[15] Jeng W Y,Wang N C,Lin M H,et al.Structural and functional analysis of three β-glucosidases from bacteriumClostridiumcellulovorans,fungusTrichodermareeseiand termiteNeotermeskoshunensis[J].Journal of Structural Biology,2011,173(1):46-56.
[16] Větrovsky T,Baldrian P,Gabriel J.Extracellular enzymes of the white-rot fungus fomes fomentarius and purification of 1,4-β-glucosidase[J].Applied Biochemistry & Biotechnology,2013,169(1):100-109.
[17] 劉中美,謝夢圓,周哲敏.黑曲霉-葡萄糖苷酶的分離純化及酶學性質[J].食品與生物技術學報,2015,34(11):1198-1202.
[18] 張大智,詹儒林,柳 鳳,等.杧果細菌性角斑病菌細胞壁降解酶的致病作用[J].果樹學報,2016(5):585-593.
[19] 金 勤,周國英,劉君昂,等.細胞壁降解酶在油茶炭疽病菌致病過程中的作用研究[J].植物保護,2017,43(3):97-102.
[20] 唐德芳,裴小瓊,李曉璐,等.黑曲霉β-葡萄糖苷酶的篩選,克隆及表達[J].應用與環境生物學報,2009,15(3):423-426.
[21] Li L,Ji Q L,Hao J F,et al.Cloning and expression of β-Glucosidase gene fromBacilluspumilus[J].Applied Mechanics & Materials,2013,301:1271-1274.
[22] Waksman G.Molecular cloning of a beta-glucosidase-encoding gene fromSclerotiniasclerotiorum,by expression inEscherichiacoli[J].Current Genetics,1989,15(4):295-297.
[23] Yin Z,Ke X,Huang D,et al.Validation of reference genes for gene expression analysis inValsamalivar.mali,using Real-time quantitative PCR[J].World Journal of Microbiology & Biotechnology,2013,29(9):1563-1571.
[24] Zhang Y,Shi X P,Li B H,et al.Salicylic acid confers enhanced resistance toGlomerellaleaf spot in apple[J].Plant Physiology and Biochemistry,2016,106:64-72.
[25] 李玉婷,趙 奇,房 冰,等.禾谷縊管蚜熱激蛋白Hsp90基因的克隆和表達分析[J].植物保護學報,2017,44(1):16-23.
[26] Kenneth J,Livak T D.Analysis of relative gene expression data using Real-time quantitative PCR and the 2-ΔΔCTmethod[J].Method,2001,25(4):402-408.
[27] 房 偉,方澤民,劉娟娟,等.新型海洋微生物β-葡萄糖苷酶基因的克隆,表達及重組酶性質[J].生物工程學報,2009,25(12):1914-1920.
[28] 陳 輝,馮志勇,馮 川,等.斑玉蕈β-葡萄糖苷酶基因的序列分析及皂苷對基因表達的影響[J].食用菌學報,2017,24(1):1-6.
[29] 陳相永,陳捷胤,肖紅利,等.植物病原真菌寄生性與分泌蛋白組CAZymes的比較分析[J].植物病理學報,2014,44(2):163-172.
[30] 王彩霞,陳曉林,李保華.腐爛病菌侵染對蘋果愈傷組織防御酶活性及丙二醛含量的影響[J].植物生理學報,2014(7):909-916.
[31] 雍道敬,李保華,王彩霞.蘋果樹腐爛病菌不同致病力菌株對蘋果的誘導效應[J].植物生理學報,2014(6):810-816.
[32] Bhat S,Goodenough P W,Bhat M K,et al.Isolation of four major subunits fromClostridiumthermocellumcellulosome and their synergism in the hydrolysis of crystalline cellulose[J].International Journal of Biological Macromolecules,1994,16(6):335-342.
