外源信號物質MeJA誘導糖槭葉部營養物質含量的變化*
2018-01-30 08:32:56劉恒旭馬曉乾
林業科技 2018年1期
關鍵詞:信號
滑 莎 繼 丹 劉 欣 劉恒旭 馬曉乾
(黑龍江省森林保護研究所,哈爾濱 150081)
糖槭(Acer saccharum)是我國東北地區主要的綠化樹種,極具觀賞價值,但在園林綠化生產中,光肩星天牛對其危害比較嚴重,而普通的化學農藥防治會對環境造成巨大的污染,因此急需環境友好型的防治措施。植物的抗性是植物在自然界長期適應周圍環境的過程中,在受到生物因子或非生物因子侵擾時表現的一種自我防御的特性,從而實現自我保護的目的[1-3,14]。寄主植物中營養物質的情況是植食性昆蟲寄主選擇的物質基礎,具有決定性的作用。寄主植物在受到病蟲害侵入時,體內會產生多途徑的代謝變化,從而形成誘導防御,如酚類物質、防御相關酶活性及營養物質的變化[4,15]。茉莉酸甲酯(MeJA)是植物體內和個體間傳遞的重要化學信號物質,在植物體內具有調節植物生長發育作用[5-10,16-17],同時還能作為信息素信號在植物與植食性昆蟲間、植食性昆蟲種間和種內傳遞[11-13]。
大量研究證明,利用外源信號物質茉莉酸類化合物可以誘導植物產生防御反應,從而增強植物抗性[16-19]。本研究通過對糖槭局部噴施茉莉酸甲酯(MeJA)后,測定其葉部營養物質含量變化情況,旨在揭示外源信號物質MeJA誘導糖槭葉部防御反應的差異,分析外源信號物質MeJA誘導對糖槭葉部蛋白質總量、可溶性糖及游離氨基酸含量的差異和變化,探究外源信號物質對糖槭營養物質的調節作用,并為危害糖槭的主要害蟲與糖槭營養物質的相互依賴關系奠定基礎。
1 材料與方法
1.1 試驗材料
MeJA購自Sigma公司,蛋白質、可溶性糖及總氨基酸測定試劑盒均購于南京建成生物制劑公司。
1.2 試驗方法
用16 L超純水中含2 mL的丙酮水溶液,配制1 mM·L-1MeJA丙酮水溶液,2014年8月7日(晴天)上午11時,在黑龍江省哈爾濱市行道綠化帶上隨機選取4棵糖槭樹,其中1棵作為對照(CK),要與其它3棵試驗用樹距離10 m以上,防止交互作用。選擇朝陽一側的樹枝,其中3棵試驗用樹噴施MeJA丙酮水溶液進行誘導處理,CK噴施丙酮水溶液,分別在誘導處理 3、6、12、21、24、48 和 96 h 時采集標本,置入便攜式冰箱帶回室內,轉入-40℃冰箱待用。測定前將采集糖槭樹葉剪碎,稱取0.100~0.900 g,移入研缽研磨成勻漿,后移入5mL離心管中,放置于-20℃冰箱保存,待測。
1.3 營養物質含量的測定
蛋白質、可溶性糖及游離氨基酸含量參照試劑盒上的方法進行測定。
1.4 數據統計及分析
利用SPSS22.0及Excel軟件對所得試驗數據進行統計和處理。
2 結果與分析
2.1 外源信號物質MeJA局部誘導糖槭葉部蛋白質含量時序的變化
通過局部噴施外源信號物質MeJA后,糖槭葉部蛋白質含量呈現增加的趨勢,在不同時間內具有一定的差異性(圖1)。糖槭葉部蛋白質含量在誘導處理3h時,與CK相比變化不明顯;當誘導處理6 h時,含量為0.484 9 mg/mL,較CK的0.386 9 mg/mL有了明顯的增加;誘導處理9 h時與誘導處理6 h時的含量水平基本一致;在誘導處理12 h時,含量為0.382 8 mg/mL,也明顯高于CK的0.290 4 mg/mL;在誘導處理21 h時,含量為0.320 0 mg/mL,較CK的0.348 9 mg/mL有所降低;當誘導處理24 h時,含量為0.5317mg/mL,明顯高于CK為0.295 4 mg/mL;誘導處理48 h和96 h時,也明顯高于CK。局部噴施外源信號物質MeJA后,誘導處理24 h時,蛋白質含量變化最大,相較CK及其他試驗誘導處理時間,蛋白質含量值最高;同時,不同時間點MeJA處理和CK蛋白質含量均表現出一定的波動性,這可能是由于采集時間、溫度、濕度和光照等環境因子造成的,但從整個試驗時間點的選取及所得到的數據可以看出,經MeJA處理后,在不同時間內,糖槭葉部蛋白質均不同程度的增加。
2.2 外源信號物質MeJA局部誘導糖槭葉部可溶性糖含量時序的變化
通過局部噴施外源信號物質MeJA后,在不同時間糖槭葉部可溶性糖含量較CK呈現出降低的現象,但每個試驗時間點降低幅度具有一定的差異性(圖2)。糖槭葉部可溶性糖含量在噴施MeJA誘導處理3h時,為4.8215mg/g,較CK的4.5084mg/g降低了0.3131mg/g;在誘導處理6h時,含量為3.9420mg/g,CK的為3.283 2 mg/g,降低了0.658 8 mg/g,降低比較明顯;在誘導處理后9、21、24和48 h時,可溶性糖含量較CK降低都比較明顯,分別降低1.8047mg/g、0.9200mg/g、1.3852mg/g和2.6694mg/g;在誘導處理后96 h時,MeJA處理與CK的可溶性糖含量基本接近。與CK相比較,在誘導處理48 h時,可溶性糖含量降低幅度最大;在整個試驗的不同時間點,經MeJA處理的糖槭葉部可溶性糖含量具有一定的波動性,在誘導處理6 h時,含量最低,誘導處理12 h時最高,這種波動性與CK的波動性基本一致,但與CK相比較,可溶性糖含量降低的幅度在不同的時間點,表現出一定的差異性。

圖1 外源信號物質MeJA誘導糖槭葉部蛋白質含量的時序變化
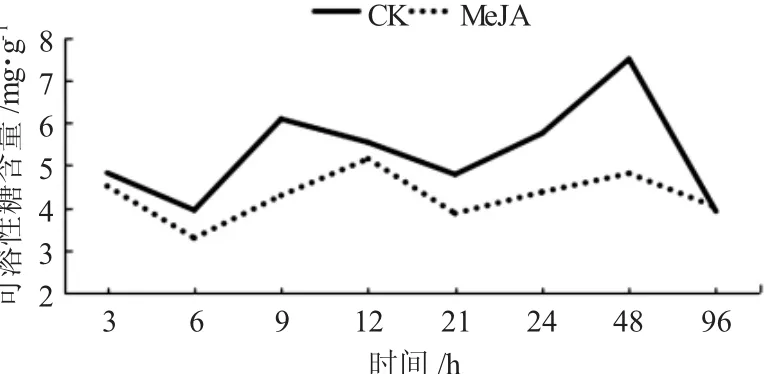
圖2 外源信號物質MeJA誘導糖槭葉部可溶性糖含量的時序變化
2.3 外源信號物質MeJA局部誘導糖槭葉部氨基酸含量時序的變化
通過局部噴施外源信號物質MeJA后,在持續不同時間利用茚三酮試劑顯色法測得糖槭葉部氨基酸含量,與CK相比較,表現出不同程度的增加,而這種增加在不同時間內具有一定差異(圖3)。在噴施MeJA處理后,游離氨基酸含量與CK比較,在誘導處理3、21、48和96 h時增加很明顯,且在誘導處理21 h時增加幅度最大;在誘導處理6 h和9 h時含量增加較明顯,而在誘導處理12 h和24 h增加不明顯。同時還可以看出,在選取的不同時間點獲得的氨基酸含量呈現出一定的波動現象,而這種現象在MeJA處理及CK試驗結果中都出現,且兩者波動基本一致。
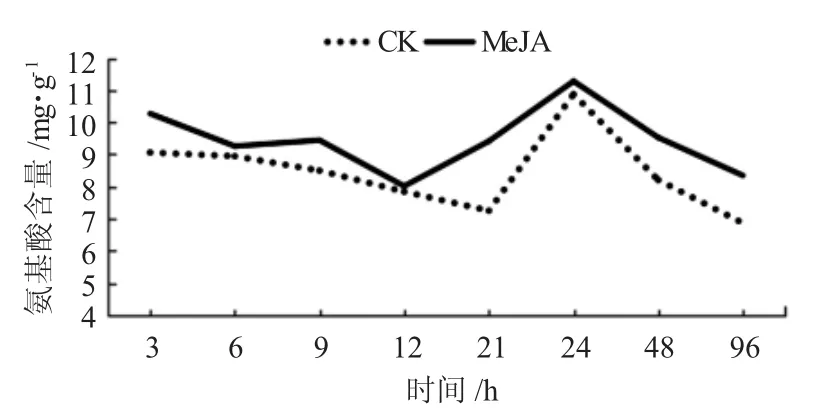
圖3 外源信號物質MeJA誘導糖槭葉部氨基酸含量的時序變化
3 結論與討論
MeJA是植物體內傳遞外界不利因素及啟動消除這種不利因素的激素物質,在植物生長過程中,扮演著重要的作用。而諸多研究證實,植物在受到生物或非生物因子侵擾時,體內營養物質等也會呈現出一定的變化產生抗性[19-20,22],噴施外源信號物質MeJA和昆蟲取食誘導寄主植物形成的抗性基本一致。在自然界中,植物主要是通過根際、氣管和滲透調節體內營養物質等一系列的變化,從而來適應生境中帶來的一系列不利因素,消弱或抑制對其生長發育的影響[22]。植食性昆蟲對寄主植物的選擇,其營養物質的差異及變化具有決定性作用,如余德億等人通過研究證實,榕管薊馬的危害程度與榕樹不同品種葉片營養物質含量關系十分密切[21]。該項研究結果也表明,局部噴施MeJA誘導了糖槭葉部營養物質發生了一定的變化,蛋白質和氨基酸含量均呈現出一定的增加現象,而可溶性糖含量呈現出降低的現象。這與Scott等[11]發現被危害的寄主植物蛋白質含量高于健康植物一致;也表明局部噴施MeJA誘導了糖槭葉部可溶性糖不同程度的降低[22]。通過外源信號物質誘導糖槭營養物質的變化情況,有助于揭示危害糖槭的主要害蟲與糖槭營養物質間相互依賴的關系,從而通過探討通過育種或者其他策略改變寄主營養物質組成,深入研究寄主植物營養物質的變化或營養物質的差異性對植食性昆蟲取食的影響具有重要的實踐意義及理論價值。
[1] Howe GA.Jasmonates as signals in the wound response[J].Jourrral ofPlant Growth Regulation,2004,23(3):223-237.
[2] Karban R,Baldwin IT.Induced Responses to herbivory[J].UniversityofChicagoPress Ltd.London,1997,187-191.
[3] Dicke M,Van Loon JJ.Multitrophic effects of herbivore-induced plant volatiles in an evolutionary context [J].Entomologia Experimentalis et Applicata,2000,97:237-249.
[4] War A R.,Sharma H C.Effect of jasmonic acid and salicylic acid induced resistance in groundnut on Helicoverpa armigera[J]..Physiological Entomology,2014,39(2):136-142.
[5] Preston CA,Laue q Baldwin IT.Methyl jasmonate is blowingin the wind,but can it act as a plant-plant airborne signal Biochem[J]..Syst Ecol,2001,29(10):1 007-1 023.
[6] Kautz S,Trisel J A,Ballhobrn D J.Jasmonic acid enhances plant cyanogenesis and resistance toherbovoryin limabean[J].Journal ofChemical Ecology,2014,40(11-12):1 186-1 196.
[7] Wasternack C.Action of jasmonates in plant stress reponses and development:applied aspect[J].Biotechnology Advances,2014,32(1):31-39.
[8] MartInez-EsplaA,ZapataPJ,CastilloS,etal.Preharvestapplication of methyl jasmonate(MeJA)in two plumcultivars.Improvement offruit growth and qualityattributes atharvest[J].Postharwest Biol.Tec.,,2014,98(3):98-105.
[9] Liao YC,Xu YH,Zhang Z,et al.Molecular cloning and sequence analysis of lipoxygenase (AsLOX1)gene in Aquilaria sinensis(Lour.)Gilg and its expression responding to MeJA and crush woundingtreatment[J].Plant Gene,2015,2(323):10-16.
[10] 孫曉玲,董文霞,蔡曉明,等.外用不同濃度茉莉酸甲酯誘導的茶樹揮發物的種類和時序變化[J].應用昆蟲學報,2016,53(3):499-506.
[11] Petty RL,Boppre M,Schneider D,et al.Identification and localization of volatile hairpencil components in male Amanris ochleabutterflies(Danaidae)[J].Experienria,1977,33(10):1 324-1 326.
[12] Hampel D,Mosandl A,Wust M.Induction of de novo volatile terpene biosynthesis via cytosolic and plastidial pathways by methyl jasmonate in foliage of Vitis vinifera[J].J.Agric.Food Chem.,2005,53(7):2 652-2 657.
[13] Hare JD.Variation in herbivore and methyl jasmonate-induced volatiles amonggenetic lines ofDatura wrightii[J]..J.Chem.Ecol.,2007,33(11):2 028-2 043.
[14] Kr ó l P,Igielski R,Pollmann S,et al.Priming of seeds with methyl jasmonate induced resistance to hemi-biotroph Fusarium oxyspornum f.sp.lycopersici in tomato via 12-oxo-phytodienoic acid,salicylicacid,andflavonolaccumulation[J].J.PlantPhysiol.,2015,179:122-132.
[15]Held D W,Boyd D.A newthrips threat[J].Ornamental Outlook,2005,14(7):14.
[16] Mantyla E,Blande JD,Klemola T.Does application of methyl jasmonate to birch mimic herbivory and attract insectivorous birds innature?[J].Arthropod-Plantinteractions,,2014,8(2):143-153.
[17] Nishida R,Baker TC,Roelofs WL.Hairpencil pheromone components of male oriental fruit moths,Grapholitha molesta[J].J.Chem.Ecol.,1982,8(6):947-959.
[18] Par é PW,Tumlinson JH.Plant volatile signals in response to herbivore feeding[J].Fla.Entomol.,1996,79(2):93-103.
[19] Petty RL,Boppré M,Schneider D,et al.Identification and localization ofvolatile hairpencil components in male Amauris ochlea butterflies (Danaidae)[J].Experientia,1977,33(10):
1 324-1 326.
[20] 曹宇,郅軍銳,孔譯賢.西花薊馬繁殖力與寄主營養物質的關系.應用昆蟲學報,2011,48(3):524-529.
[21] 余德億,姚錦愛,黃鵬,等.榕管薊馬危害與寄主葉片結構及營養物質的關系田.南京農業大學學報,2014,37(2):38-44。
[22] 馬曉乾,趙紅盈,鄧勛,等.模擬昆蟲取食后紅皮云杉幼苗營養物質的時序變化.東北林業大學學報,2013,41(3):24-25,29.
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
