組胺對抑郁癥大鼠運動欲望與空間記憶能力的影響
2018-01-31 06:27:15何葉成李東印
中國比較醫學雜志 2018年1期
何葉成,姚 娟,陳 尚,李東印
(蘇州衛生職業技術學院,江蘇 蘇州 215009)
近年來抑郁癥的發病率呈快速增長趨勢,其主要表現為情緒低落并且伴隨認知功能減退[1-2]。抑郁癥致病因素和臨床癥狀呈多樣性,發病機制至今尚未完全闡明。截至目前為止,下丘腦-垂體-腎上腺軸(HPA軸)功能亢進學說為多數學者認可的抑郁癥主要發病機制[3-4]。然而,外界應激刺激信號作用于機體導致下丘腦興奮后,除了通過HPA軸-體液途徑發揮作用,其傳出神經通路在抑郁癥的發生發展過程中發揮了什么作用還有待進一步研究。
組胺(histamine)是繼5-羥色胺(5-HT)、去甲腎上腺素(NE)之后在腦內新發現的一類單胺類神經遞質,中樞組胺能神經元都聚集在下丘腦,神經纖維投射遍布各個腦區[5-6],調節神經元基礎興奮性從而對各個腦區功能進行調控[7]。海馬作為邊緣系統重要組成部分,在機體情緒和認知的調節過程中發揮重要作用。神經解剖學研究顯示,下丘腦發出大量組胺能神經纖維投射到海馬[7-8],提示下丘腦可能通過神經通路影響海馬神經元的功能狀態,進而對機體情緒和認知能力產生影響。C1區作為海馬主要信號傳出通路,是空間記憶形成的關鍵部位[3, 9]。本文以大鼠海馬C1區為研究目標,通過離體腦片和在體細胞外記錄的方法觀察了組胺對海馬C1區神經元放電頻率的調節作用,并在整體水平觀察了海馬C1區注射組胺相關試劑對抑郁癥模型大鼠運動欲望和空間記憶能力的影響,探索單胺類遞質組胺在抑郁癥發生發展中所起的作用。
1 材料和方法
1.1 實驗動物
本文共使用216只清潔級雄性成年SD大鼠(150~250 g),由蘇州大學實驗動物中心供應 [SCXK (蘇) 2012-0005]。單獨飼養,常溫環境,12 h明/暗交替,供給充足的標準大鼠飼料和水 [SYXK (蘇) 2012-0045]。
1.2 主要試劑
組胺(Tocris,1A/198112);組胺H1受體激動劑2-pyridylethylamine(Tocris,2A/195553);組胺H1受體阻斷劑triprolidine(Tocris,2A/189683);組胺H2受體激動劑dimaprit(Tocris,2B/182087);組胺H2受體阻斷劑ranitidine(Tocris,1A/194394);組胺H3受體激動劑IMETIT(Tocris,5A/160009);組胺H3受體阻斷劑clobenpropit(Tocris,3B/185670)。組胺和組胺受體激動劑、阻斷劑的母液用蒸餾水配置,冷凍保存。使用前再用新鮮人工腦脊液將各藥品母液稀釋至實驗所需的濃度。
1.3 實驗方法
1.3.1 抑郁癥模型大鼠的制備
采用經典的慢性未預知應激法構建大鼠抑郁癥模型[10-11],慢性應激刺激方式包括:禁水(鼠籠無水瓶12 h后放空瓶1 h,其余時間正常飲水);禁食(鼠籠無鼠糧12 h后放3 g鼠糧2 h,其余時間正常飲食);夾尾l min;電擊(50 V電壓電擊大鼠15次,每次10 s,中間間隔30 s);冷刺激(在4℃冰水中強迫游泳5 min);熱刺激(在45℃烘箱中6 min);將墊料浸濕,持續24 h。一共7種形式,每日隨機選取一種,持續刺激21 d,每種刺激出現3次,同種刺激不能連續出現,使動物不能預料刺激的發生,并以曠場實驗鑒定造模成功與否。實驗過程中按實驗動物使用的3R原則給予人道主義關懷。
1.3.2 離體腦片細胞外記錄
(1)腦片的制備:SD大鼠經乙醚麻醉,迅速斷頭取腦,將含有海馬部分的腦組織塊放入人工腦脊液中,通以95% O2/5% CO2混合氣體,溫度保持在4℃左右。根據定位圖譜[12],將腦組織塊切片,厚度為350~400 μm,選取含有海馬神經元的腦片并將其轉移到記錄浴槽內進行孵育。記錄浴槽被持續地灌流人工腦脊液孵育40 min后開始記錄神經元放電[13]。
(2)離體腦片細胞外記錄:采用玻璃微電極記錄海馬C1區神經元的自發放電活動。記錄到神經元自發放電活動之后,維持20 min,確認其放電活動處于穩定狀態。然后在灌流的人工腦脊液中加入藥物刺激目標神經元,觀察細胞的反應。神經元的放電信號輸入計算機,用Spike 2(CED,UK)軟件分析神經元的放電頻率,并做出刺激-時間直方圖,采樣間隔1 s,以評價組胺相關試劑對海馬C1區神經元的作用[14]。
1.3.3 在體細胞外記錄
(1)動物手術:由烏拉坦(0.8 g/kg)和α-氯醛糖(65 mg/kg)組成的混合麻醉劑,經腹腔注射麻醉SD大鼠,將動物固定于腦立體定位儀上。依據鼠腦立體定位圖譜找到下丘腦結節乳頭體核(tuberomammillary nucleus,TMN)和同側海馬C1區上方,打開顱骨并移除硬腦膜,腦表面暴露后并覆以2%瓊脂。將同心雙極電極(內徑0.1 mm,外徑0.4 mm,尖端0.2 mm)插入下丘腦TMN以備刺激,將微量注射器針頭插至海馬C1區上方,用以加藥[15]。
(2)刺激、電生理學記錄、數據的采集和分析:以負相矩形雙脈沖形式電刺激下丘腦TMN,用玻璃微電極記錄海馬C1區神經元的自發放電(與打藥針頭呈30°角進針)。海馬神經元的放電信號輸入計算機,用Spike 2軟件進行實時數據分析。通過比較刺激或給藥前后神經元的放電頻率變化,判斷所記錄海馬神經元的反應類型。對兩窗口內的放電頻率進行t檢驗,P< 0.05為差異有顯著性[15]。
1.3.4 行為學實驗
(1)手術和定點微量注射:腹腔注射戊巴比妥鈉麻醉SD大鼠,將其頭部固定于立體定位儀上。根據鼠腦立體定位圖譜在海馬C1區上方開顱,并將用于微量注射的兩根不銹鋼套管(長6 mm,外徑0.8 mm,內徑0.5 mm)植入大腦,使其下端分別定位在雙側海馬C1區上方2 mm的位置,用不銹鋼螺絲和牙科水泥固定于顱骨。手術后每只大鼠腹腔注射青霉素,并給予72 h的恢復期[16-17]。
給藥時使用的微量注射器不銹鋼針管長8 mm,其下端正好位于海馬C1區表面,不至損傷神經核團。雙側同時緩慢注射,每側每次注射量為1 μL,過程持續60 s以上。
(2)分組:完成手術的抑郁癥模型大鼠恢復后,進行隨機分組,共分8組,每組15只:對照組、histamine組、2-PyEA(H1受體激動劑)組、triprolidine(H1受體阻斷劑)組、dimaprit(H2受體激動劑)組、ranitidine(H2受體阻斷劑)組、IMETIT(H3受體激動劑)組和clobenpropit(H3受體阻斷劑)組。各組以注射藥物命名,對照組不注射藥物。以上藥物濃度均為10 mmol/L,劑量每次每側注射1 μL。根據離體腦片細胞外記錄實驗[16],組胺及其相關試劑灌流濃度在10 μmol/L左右能夠引起神經元發生明顯的反應,而1 μL的藥液注射至大鼠腦內擴散體積約為1 mL,稀釋了約1000倍,故將在體實驗注射藥物濃度提高至10 mmol/L。
(3)曠場實驗:曠場實驗主要檢測動物在新環境中的運動和探索欲望,是評價大鼠抑郁程度的經典方法,分水平運動和垂直運動兩個觀察指標,以觀察得分為評分計算單位。將大鼠至于一個獨立的空間(50 cm × 50 cm × 40 cm),整個開場被劃分成(5 × 5)個邊長為10 cm的方格。輕輕地將大鼠放入空間底部正中間,觀察大鼠在5 min內水平和垂直兩個方向的得分(水平得分:三爪均進入一個方格計一分;垂直得分:大鼠直立一次計一分)[13]。在曠場實驗中,水平得分代表了大鼠的興奮程度,垂直得分代表了大鼠對新環境的探索欲望。造模結束后,大鼠水平或者垂直得分與正常大鼠相比如果減少程度高于30%,則認為造模成功。每組大鼠造模成功后,第2天相同時間段打藥后再進行檢測對比。
(4)Morris水迷宮實驗:Morris水迷宮是檢測大鼠學習記憶能力的經典實驗。水迷宮直徑120 cm,高60 cm,實驗時注入23℃溫水至23 cm深。將水池平均分為四個象限,隨機取一個象限,放入一個高20 cm的白色平臺,位置保持固定,用食用鈦白粉將水染成白色使平臺隱蔽。實驗前先對SD大鼠進行訓練,訓練時將大鼠背對平臺放入水中,并記錄大鼠入水后找到平臺所需要的時間,記為逃避潛伏期。大鼠找到平臺后在其上停留20 s,然后擦干放回籠內。每次訓練選擇不同的象限入水,每兩次訓練間隔大于30 s,訓練持續4 d使大鼠逃避潛伏期保持穩定。第5天和第7天為測試期,測試時撤去平臺,記錄大鼠60 s內游過原平臺所在象限的時間[18]。
1.3.5 組織學鑒定
測試完成后,動物被深度麻醉,斷頭取腦。固定一周后,作厚度為80 μm的連續冠狀面冰凍切片,根據大鼠腦圖譜判讀插管創道的位置,如果插痕沒有到達預期位置,測試結果將被排除在數據統計之外。
1.4 統計學方法
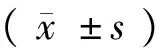
2 結果
2.1 離體腦片細胞外記錄
我們在36張海馬腦片上記錄了56個具有穩定自發放電頻率的海馬C1區神經元,其自發放電頻率范圍為1.78~2.86 Hz,平均放電頻率為(2.15±0.23) Hz。用含有組胺(3~30 μmol/L)的人工腦脊液灌流腦片,引起75%(42/56)的海馬神經元興奮反應(圖1)。所記錄到的海馬神經元對3、10和30 μmol/L的組胺刺激表現出劑量依賴性的興奮反應,神經元接受組胺刺激后的自發放電頻率峰值分別為2.85、3.25和3.54 Hz,與其接受刺激前的平均基礎自發放電頻率(2.15 Hz)相比,分別增加了32.6%、51.2%和64.7%。
2.2 在體細胞外記錄
我們在本實驗中共記錄了57個海馬C1區神經元,其中38個細胞(66.7%)對同側下丘腦結節乳頭核刺激有反應,均表現為先興奮后抑制的雙向反應,綜合表現出抑制效應(圖2,n=38,P< 0.05)。如果在海馬C1區微量注射組胺H1受體阻斷劑triprolidine(1 μmol/L)后電刺激結節乳頭核,則海馬神經元表現為簡單的抑制反應(n=13,P< 0.05);如果注射組胺H3受體阻斷劑clobenpropit(1 μmol/L)后電刺激結節乳頭核,則海馬神經元表現為簡單的興奮反應(n=15,P< 0.05)。

注:A:電刺激下丘腦結節乳頭核,海馬C1區神經元表現出先興奮后抑制的雙向反應;▲為電刺激時間點。B:在海馬C1區注射組胺H1受體阻斷劑triprolidine后,再電刺激下丘腦結節乳頭核,海馬C1區神經元表現出單一的抑制效應。C:在海馬C1區注射組胺H3受體阻斷劑clobenpropit后,再電刺激下丘腦結節乳頭核,海馬C1區神經元表現出單一的興奮效應。圖2 在體細胞外記錄Note. A: After the hypothalamus TMN was electrically stimulated, neurons in the hippocampal C1 area showed a bidirectional response, an excitatory response followed by an inhibitory response. ▲ represents the time point of electrical stimulation. B: Under electrical stimulation of TMN after injection of histamine H1 receptor antagonist, triprolidine into the hippocampal C1 area, hippocampal neurons only showed an inhibitory response. C: Under electrical stimulation of TMN after injection of histamine H3 receptor antagonist, clobenpropit into the hippocampal C1 area, hippocampal neurons only showed an excitatory response.Fig.2 Extracellular recording in vivo
2.3 行為學實驗
行為學實驗共分8組,每組15只大鼠。實驗結束后斷頭取腦切片,共有10只大鼠插痕偏離預期位置,這10只大鼠的實驗數據被排除在統計數據之外。
2.3.1 曠場實驗
曠場實驗檢測結果代表了大鼠運動欲望,具體數值見表1。水平維度,各組得分通過單因素方差分析顯示,F7, 112=0.168,P< 0.05。與對照組相比,注射組胺后得分明顯降低,注射H3受體阻斷劑clobenpropit后得分顯著升高,均P< 0.01。垂直維度,各組得分通過單因素方差分析顯示,F7, 112=0.253,P< 0.05。與對照組相比,注射組胺后得分明顯降低,而注射H3受體阻斷劑clobenpropit后得分顯著升高,均P< 0.01。

表1 大鼠曠場實驗測試成績
注:與對照組相比,*P< 0.05,**P< 0.01。
Note. Compared with the control group,*P< 0.05,**P< 0.01.
2.3.2 Morris水迷宮
水迷宮實驗檢測結果見表2,通過單因素方差分析顯示,F7, 102=0.024,P< 0.05。在抑郁癥模型大鼠海馬C1區注射組胺(n=15)后,逃避潛伏期與對照組相比明顯延長,P< 0.01;而注射組胺H3受體阻斷劑clobenpropit(n=15)后,逃避潛伏期明顯縮短,P< 0.01。

表2 大鼠Morris水迷宮測試期成績
注:與對照組相比,*P< 0.05,**P< 0.01。
Note. Compared with the control group,*P< 0.05,**P< 0.01.
3 討論
海馬作為邊緣系統重要組成部分,參與調節情緒、學習和記憶等功能,是抑郁癥發病機制中被廣泛研究的中樞結構之一[9,19-20]。神經系統形態學研究顯示,抑郁癥患者出現海馬體積減小并伴有神經元缺失等退化現象,導致情緒低落并伴隨認知記憶能力明顯下降[3]。然而,抑郁癥患者海馬中留存的神經元功能狀態如何,基礎放電頻率有何變化,還未見詳細報道。近年來,隨著對腦內單胺類遞質組胺的深入研究,起源于下丘腦的中樞組胺能神經系統被認為是“全腦功能調節者”,能夠調節各腦區神經元基礎興奮性,影響其放電頻率[6-7,21]。神經免疫組化研究表明,海馬神經元上有大量組胺受體表達[8,22-24],接受組胺能神經纖維的密集支配。因此,機體受到外界應激刺激導致下丘腦興奮后,可能通過傳出神經纖維調節邊緣系統海馬神經元興奮程度,進而對機體情緒和認知功能產生影響。
本實驗中,離體腦片細胞外記錄研究顯示,組胺能夠以劑量依賴的方式興奮海馬C1區神經元,且能夠被組胺H1受體阻斷劑triprolidine而不是H2受體阻斷劑ranitidine所阻斷,提示組胺能夠通過H1受體途徑興奮海馬C1區神經元。在體細胞外記錄研究顯示,刺激下丘腦結節乳頭核,能夠使海馬神經元出現先興奮后抑制的雙向反應;C1區注射組胺H1受體阻斷劑triprolidine后興奮效應消失,出現單一的抑制效應;注射組胺H3受體阻斷劑clobenpropit后,抑制效應消失,出現單一的興奮效應。這個結果提示海馬C1區組胺突觸后H1受體和突觸前H3受體同時發揮效應,組胺H1受體興奮導致海馬C1區神經元放電頻率提高;而組胺H3受體興奮能夠抑制海馬C1區興奮性遞質的釋放,使海馬神經元放電頻率降低;兩者的綜合效應是海馬神經元的放電頻率降低。在整體行為學實驗中,海馬C1區微量注射組胺和H1受體激動劑2-PyEA,都能使抑郁癥大鼠運動欲望和空間記憶能力進一步降低,而注射組胺H3受體阻斷劑clobenpropit能夠使抑郁癥大鼠運動欲望及空間記憶能力明顯改善。
單胺類神經遞質如5-HT、NE等被認為在抑郁癥的發生發展過程中起到了重要作用。組胺作為起源于下丘腦的單胺類神經遞質,能夠對包括邊緣系統在內的各個腦區神經元興奮性進行調節[6-7]。機體受到應激刺激導致下丘腦興奮,必然會導致傳出神經纖維組胺釋放的改變,而組胺在抑郁癥的發生發展中所起的作用還未見詳細報道。本研究發現電刺激下丘腦TMN后,組胺能神經纖維釋放組胺增加能夠降低海馬神經元放電頻率,從而使抑郁癥模型大鼠運動欲望和空間記憶能力明顯降低,而注射clobenpropit阻斷H3受體作用途徑后,抑郁癥模型大鼠運動欲望和空間記憶能力明顯改善。由此可見,在抑郁癥發生發展過程中,下丘腦興奮不僅通過HPA軸功能亢進導致腦內皮質醇濃度增高,使海馬神經元凋亡缺失;還可以通過中樞組胺能神經系統,使海馬神經元放電頻率降低,抑制海馬神經元功能狀態,并導致抑郁癥大鼠運動欲望和認知能力的降低。考慮到中樞組胺能神經系統起源于應激反應高級中樞下丘腦,同時組胺能神經纖維廣泛支配了包括海馬在內的抑郁癥發病相關腦區神經元的興奮性,因此單胺類遞質組胺可能在抑郁癥的發生發展中發揮了重要作用,應當受到重視。
[1] Otte C, Gold SM, Penninx BW, et al. Major depressive disorder [J]. Nat Rev Dis Primers, 2016, 2: 16065.
[2] Willner P, Mitchell PJ. The validity of animal models of predisposition to depression [J]. Behav Pharmacol, 2002, 13(3): 169-188.
[3] Kim EJ, Pellman B, Kim JJ. Stress effects on the hippocampus: a critical review [J]. Learn Mem, 2015, 22(9): 411-416.
[4] Gotlib IH, Joormann J, Minor KL, et al. HPA axis reactivity: a mechanism underlying the associations among 5-HTTLPR, stress, and depression [J]. Biol Psychiatry, 2008, 63(9): 847-851.
[5] Panula P, Yang HY, Costa E. Histamine-containing neurons in the rat hypothalamus [J]. Proc Natl Acad Sci U S A, 1984, 81(8): 2572-2576.
[6] Haas HL, Sergeeva OA, Selbach O. Histamine in the nervous system [J]. Physiol Rev, 2008, 88(3): 1183-1241.
[7] Zhu JN, Yung WH, Kwok-Chong Chow B, et al. The cerebellar-hypothalamic circuits: potential pathways underlying cerebellar involvement in somatic-visceral integration [J]. Brain Res Rev, 2006, 52(1): 93-106.
[8] Thomas SA. Neuromodulatory signaling in hippocampus-dependent memory retrieval [J]. Hippocampus, 2015, 25(4): 415-431.
[9] Anderson EB, Grossrubatscher I, Frank L. Dynamic hippocampal circuits support learning- and memory-guided behaviors [J]. Cold Spring Harb Symp Quant Biol, 2014, 79: 51-58.
[10] Katz RJ, Roth KA, Carroll BJ. Acute and chronic stress effects on open field activity in the rat: implications for a model of depression [J]. Neurosci Biobehav Rev, 1981, 5(2): 247-251.
[11] 張榮, 董世芬, 倪博然, 等. 白金膠囊對CUMS誘導的抑郁癥大鼠模型行為學及腦內單胺類神經遞質的影響 [J]. 中國比較醫學雜志, 2015, 25(7): 46-52.
[12] Paxinos G, Watson C. The rat brain in stereotaxic coordinates [M]. 6th edition. San Diego: Academic Press, 1998.
[13] He YC, Wu GY, Li D, et al. Histamine promotes rat motor performances by activation of H2receptors in the cerebellar fastigial nucleus [J]. Behav Brain Res, 2012, 228(1): 44-52.
[14] Peng SY, Zhuang QX, He YC, et al. Histamine excites neurons of the inferior vestibular nucleus in rats by activation of H1 and H2 receptors [J]. Neurosci Lett, 2013, 541: 87-92.
[15] Li B, Zhuang QX, Gao HR, et al. Medial cerebellar nucleus projects to feeding-related neurons in the ventromedial hypothalamic nucleus in rats [J]. Brain Struct Funct, 2017, 222(2): 957-971.
[16] Song YN, Li HZ, Zhu JN, et al. Histamine improves rat rota-rod and balance beam performances through H2receptors in the cerebellar interpositus nucleus [J]. Neuroscience, 2006, 140(1): 33-43.
[17] 孟盼, 朱青, 張秀麗, 等. 海馬內微量注射K252a對大鼠行為學及抑郁樣經典指標的影響 [J]. 中國比較醫學雜志, 2017, 27(4): 26-33.
[18] Rice JP, Wallace DG, Hamilton DA. Lesions of the hippocampus or dorsolateral striatum disrupt distinct aspects of spatial navigation strategies based on proximal and distal information in a cued variant of the Morris water task [J]. Behav Brain Res, 2015, 289: 105-117.
[19] Lazarov O, Hollands C. Hippocampal neurogenesis: learning to remember [J]. Prog Neurobiol, 2016, 138-140: 1-18.
[20] Rozov AV, Valiullina FF, Bolshakov AP. Mechanisms of long-term plasticity of hippocampal GABAergic synapses [J]. Biochemistry (Mosc), 2017, 82(3): 257-263.
[21] Cataldi M, Borriello F, Granata F, et al. Histamine receptors and antihistamines: from discovery to clinical applications [J]. Chem Immunol Allergy, 2014, 100: 214-226.
[22] Chang RS, Tran VT, Snyder SH. Heterogeneity of histamine H1-receptors: species variations in [3H]mepyramine binding of brain membranes [J]. J Neurochem, 1979, 32(6): 1653-1663.
[23] Diewald L, Heimrich B, Büsselberg D, et al. Histaminergic system in co-cultures of hippocampus and posterior hypothalamus: a morphological and electrophysiological study in the rat [J]. Eur J Neurosci, 1997, 9(11): 2406-2413.
[24] Lozeva V, Tarhanen J, Attila M, et al. Brain histamine and histamine H3receptors following repeatedL-histidineadministration in rats [J]. Life Sci, 2003, 73(12): 1491-1503.
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
作文周刊·小學二年級版(2022年20期)2022-05-05 01:33:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
創新作文(小學版)(2019年10期)2019-09-25 08:12:28
小學生學習指導(低年級)(2017年5期)2017-05-04 04:14:38
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
大灰狼(2015年6期)2015-07-16 21:01:00
作文與考試·小學高年級版(2015年17期)2015-05-30 10:48:04
