羅氏沼蝦凍藏過程中質構與理化特性研究
2018-03-11 02:37:32柳佳彤姜啟興許艷順夏文水
食品與機械 2018年12期
柳佳彤 姜啟興 楊 方 王 斌 許艷順 夏文水 徐 巖
(1. 江南大學食品學院,江蘇 無錫 214122;2. 江蘇省食品安全與質量控制協同創新中心,江蘇 無錫 214122;3. 江南大學生物工程學院,江蘇 無錫 214122)
羅氏沼蝦(Macrobrachiumrosenbergii)又稱馬來西亞大蝦、淡水長臂大蝦,2016年中國羅氏沼蝦養殖產量為1.327×105t,比2015年增加2.49%[1]。羅氏沼蝦個體大,營養豐富,因此備受廣大消費者青睞,但其生長適宜水溫為20~34 ℃,季節性較強,使得市場銷售僅限于7~10月,所以目前很多企業將其加工成冷凍產品,以滿足消費者全年消費的需求。但是發現在傳統的速凍工藝及凍藏條件下,隨著凍藏時間的延長,蝦肉會變得松軟無彈性,嚴重影響產品的食用品質,因此迫切需要開展凍藏羅氏沼蝦品質劣變相關的研究,從而為開發高品質冷凍羅氏沼蝦加工關鍵技術提供理論依據。
研究[2]4-10表明,內源酶降解作用是導致水產品低溫保藏過程中品質變化的主要原因。Ge等[3]研究了草魚冰藏條件下組織蛋白酶在貯藏期間的活性和分布規律,并確定了內源組織蛋白酶和鈣激活蛋白酶是造成冰藏草魚質構劣化的主要因素。Navarrete等[4]研究發現凡納濱對蝦肝胰腺中的胰凝乳蛋白酶對膠原蛋白等底物水解性能很高。陳詩研等[5]發現凡納濱對蝦在冰藏過程中蝦頭中的內源酶會隨著貯藏時間的延長遷移到肉中。吉宏武等[6]研究了南美白對蝦冰藏期間內源酶對蝦肉蛋白的降解作用,發現隨著貯藏時間的延長,內源酶對肌原纖維蛋白的肌球蛋白重鏈(MHC)和肌動蛋白(Actin)降解程度增大,其中蝦頭中胰蛋白酶對肌原纖維蛋白的降解作用最強。顏龍杰等[7]發現凡納濱對蝦在4 ℃貯藏5 d后,組織蛋白酶L的相對活性仍保持在80%以上,SDS-PAGE分析發現組織蛋白酶L在貯藏過程中持續參與蝦肌肉蛋白的分解,尤其對組成肌肉的主要蛋白MHC的降解最為顯著。而目前中國關羅氏沼蝦尤其是對其凍藏期間質構劣變和內源酶變化的研究相對較少,且主要集中在凍藏溫度和凍結方式等條件對蝦類多酚氧化酶活性的影響,尚未見有關凍藏期間內源蛋白酶的變化情況及其與質構劣變相關性方面的研究。因此,本研究擬以羅氏沼蝦為研究對象,并以刀額新對蝦為對照,研究凍藏對羅氏沼蝦蛋白劣化和質構特性的影響,考察蝦頭中蛋白酶向蝦肉中遷移的情況,并分析蛋白劣化和內源酶之間的相關性,以期為闡明羅氏沼蝦品質劣變的機理提供理論依據。
1 材料與方法
1.1 材料與試劑
鮮活羅氏沼蝦、刀額新對蝦:購于江蘇省無錫市濱湖區華潤萬家超市,羅氏沼蝦平均體質量(25.00±5.00) g,體長(10.00±2.00) cm;刀額新對蝦平均體質量(10.50±0.50) g,體長(10.00±2.00) cm,在塑料袋中加入適量碎冰,30 min內活運至實驗室;
甲醛、三氯乙酸、NaOH、KCl、NaCl、EGTANa2、MgCl2、KH2PO4、K2HPO4、K3PO4:分析純,國藥集團化學試劑有限公司;
平板計數瓊脂:生化試劑,國藥集團化學試劑有限公司。
1.2 儀器與設備
FRINOX速凍柜:FDS-35型,上海漯城餐飲設備有限公司;
高速冷凍離心機:3K-30 型,德國 Sigma 公司;
物性分析儀:TA-XT2i 型,英國 Stable Micro System 公司;
紫外可見分光光度計:UV-1000型,上海天美科學儀器有限公司;
激光粒度分析儀:S3500 型,美國Microtrac公司;
高速分散機:T18型,德國 IKA 公司;
pH計:EL20 型,梅特勒-托利多儀器上海有限公司;
立式臺式殺菌鍋:LDZX-50KBS型,上海躍進醫療器械廠;
超凈工作臺:S.SW-CJ-1F型,上海躍進醫療器械廠;
恒溫培養箱:PYX-DHS·350-BY-II型,上海躍進醫療器械廠。
1.3 試驗方法
1.3.1 樣品處理與貯藏試驗 將鮮活的羅氏沼蝦和刀額新對蝦加冰猝死,低溫流水沖洗干凈后分成3組。A:整只羅氏沼蝦凍品;B:去頭羅氏沼蝦凍品;C:整只刀額新對蝦凍品。A、B、C 3組單層擺放于不銹鋼盤上,置于(-33±2) ℃的鼓風速凍機中進行凍結,用直徑約1 mm的溫度探針插入蝦肉第一腹節中心位置待其溫度達到-18 ℃時取出,隨機裝入自封袋中,于(-18±2) ℃冰箱中貯藏。貯藏過程中每隔2周隨機取樣,置于4 ℃冰箱中過夜解凍,解凍后去頭去殼取蝦肉進行質構、理化和蛋白酶活等相關指標的分析。
1.3.2 剪切力測定 取蝦的一、二腹節,采用質構分析儀,垂直于肌肉纖維走向進行測定,每組樣品至少6個平行樣。參數設定:探頭型號A/CKB,刀片探頭下降速度 2 mm/s,壓縮比90%。
1.3.3 TCA-溶解肽測定 參照Sriket等[8]的方法稍加改動。將蝦解凍處理后取一、二腹節蝦肉絞碎,稱取2 g蝦肉,加入質量分數5%的三氯乙酸(TCA)溶液18 mL,用高速分散機均質1 min,冰浴30 min后,于4 ℃、11 000×g離心10 min,上清液中肽的含量用lowry法[9]測定,結果用μmol Tyr/g 肉表示。每組樣品做3個平行樣。
1.3.4 肌原纖維小片化指數(MFI值)測定 根據文獻[9],按式(1)計算MFI值。每組樣品做3個平行樣。
MFI=A540×200,
(1)
式中:
MFI——肌原纖維小片化指數;
A540——540 nm處的吸光度值。
1.3.5 肌原纖維表觀直徑(D3,2)測定 參照Culler等[10]方法并略有改動。蝦解凍處理后取一、二腹節肉絞碎,取2 g肉加入30 mL預冷緩沖液(100 mmol/L KCl,20 mmol/L K3PO4,1 mmol/L EDTA,1 mmol/L MgCl2,pH 7.2)均質1 min,于4 ℃、1 000×g離心15 min,所得沉淀重復上述操作,再次得到的沉淀懸于15 mL上述緩沖液中,取1.5 mL用粒度分析儀測定粒徑分布,按照規則球體的折射系數1.54計算粒徑。肌原纖維表觀直徑以表面積計算的平均粒徑(MA)表示,即D3,2(μm)。每組樣品做3個平行樣。
1.3.6 蝦肉中總蛋白酶活測定 參照Sriket等[11]方法略有改動。蝦解凍處理后取一、二腹節蝦肉分別絞碎,加入3倍體積的磷酸鹽緩沖液(pH 7.6),均質,置于冰浴(<4 ℃)中,用磁力攪拌器連續攪拌30 min后,10 000×g離心30 min,上清液即為粗酶液。移取200 μL粗酶液加入200 μL蒸餾水和625 μL反應緩沖液(0.2 mol/L Na2HPO4-0.1 mol/L 檸檬酸,pH 5.0),最后加入10 g/L酪蛋白溶液200 μL,于50 ℃ 恒溫水浴反應15 min,加入質量分數50% TCA溶液200 μL終止反應,10 000×g、4 ℃離心10 min,上清液中肽的含量用lowry法測定。一個酶活單位(U)定義為50 ℃下每分鐘水解酪蛋白并釋放1 μmol酪氨酸的量。每組樣品做3個平行樣。
1.3.7 菌落總數測定 樣品的處理參照GB 4789.20—2003中蝦類檢樣的處理方法,于無菌條件下取蝦腹節內的肌肉2.5 g 放入滅菌乳缽中,用滅菌剪子剪碎加入22.5 mL生理鹽水,均質后,依次梯度稀釋;菌落總數的檢測參照GB 4789.2—2016,采用傾注平板法按照水產品菌落總數的培養條件在平板計數培養基(PCA)上(30±1) ℃培養(72±3) h。
1.3.8 微觀結構觀察 根據文獻[12]。
1.4 數據統計分析
數據采用Origin繪圖,采用SPSS對數據進行統計分析,利用Pearson雙變量分析法進行相關性計算分析。
2 結果與分析
2.1 凍藏過程中蝦肉的剪切力變化情況
質構特性是水產品加工過程中評價產品質量和可接受程度的一個重要指標。其中剪切力是表征質構變化常用的指標之一[13]。整只羅氏沼蝦凍品、去頭羅氏沼蝦和整只刀額新對蝦凍品在凍藏過程中剪切力的變化情況如圖1所示。由圖1可知,在整個凍藏期間,3組樣品的剪切力均隨著凍藏時間的延長而下降,貯藏16周后整只羅氏沼蝦凍品、去頭羅氏沼蝦凍品和整只刀額新對蝦凍品剪切力分別下降了36.81%,25.40%,22.83%。整只羅氏沼蝦凍品基本呈直線下降趨勢。整個凍藏周期內整只刀額新對蝦剪切力值始終高于其他樣品,且下降幅度最小。凍藏前期去頭羅氏沼蝦剪切力明顯低于整只羅氏沼蝦凍品,但在8周之后去頭羅氏沼蝦剪切力下降幅度變小,并在10周之后高于整只羅氏沼蝦凍品,圖2~4也呈現類似變化趨勢,去頭羅氏沼蝦凍品在前期各項指標劣化的程度要大于整只羅氏沼蝦凍品,而在后期劣變程度則低于整只羅氏沼蝦凍品。以上結果表明,相比于整只刀額新對蝦凍品和去頭羅氏沼蝦凍品,整只羅氏沼蝦凍品剪切力下降程度最大,說明凍藏期間其品質劣化最大。
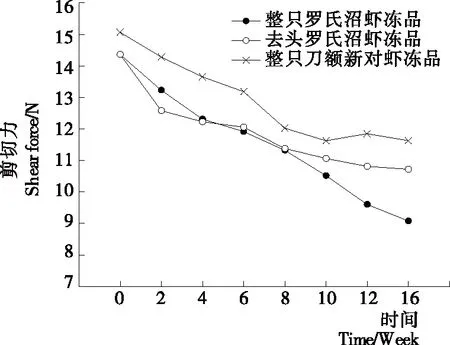
圖1 凍藏過程中蝦肉的剪切力變化情況Figure 1 Changes in shear force of different samples during frozen storage
2.2 凍藏過程中肌原纖維小片化指數(MFI)變化
MFI值反映了肌節Ι帶附近關鍵細胞骨架蛋白的降解程度[14]。MFI 越大,表明肌原纖維內部結構完整性破壞的程度就越大[15]。由圖2可知,隨著凍藏時間的延長,整只羅氏沼蝦凍品、去頭羅氏沼蝦凍品和整只刀額新對蝦凍品的MFI值均呈上升趨勢,且分別在前2周、前4周和前8周上升的較快,之后變化較平緩。整只羅氏沼蝦凍品、去頭羅氏沼蝦凍品和整只刀額新對蝦凍品在凍藏周期內分別較初始值上升了1.45,1.70,1.86倍。2~16周內3組樣品之間一直存在顯著性差異,且較為明顯(P<0.05)。凍藏初期去頭羅氏沼蝦的MFI值高于整只羅氏沼蝦凍品,可能是去頭過程對蝦肉造成了損傷,使部分細胞破裂,細胞內酶釋放出來,從而加劇了蛋白的降解和肌肉結構的破壞;從第4周開始整只羅氏沼蝦凍品的MFI值開始超過去頭羅氏沼蝦凍品,可能是隨著凍藏時間的延長,蝦頭中的內源蛋白酶緩慢遷移到蝦肉中,從而加劇了對蝦肉蛋白的降解和結構的破壞,這可以從圖5凍藏過程中不同羅氏沼蝦樣品中總蛋白酶活的變化得到驗證。而圖4中TCA-溶解肽的變化趨勢基本與MFI值的變化類似,也初步說明蛋白的降解可能是導致品質變化的原因之一。Jiang等[16]通過掃描電鏡(SEM)觀察烏鱧魚片在-20 ℃貯藏時肌原纖維的變化情況,發現不同凍結方式和溫度的烏鱧魚片在凍藏期間肌原纖維都會出現不同程度的片狀化,并推測這一變化是由蛋白脫水導致的,這些變化會直接引起魚片質構和持水力的變化。Kaale等[17]研究表明大西洋鮭魚微凍保藏過程中冰晶的形成往往伴隨著肌原纖維的碎片化。
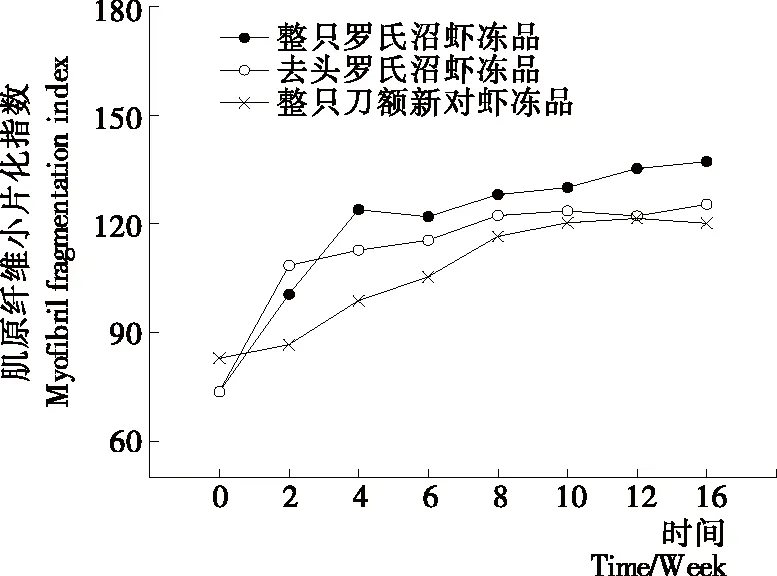
圖2 凍藏過程中肌原纖維小片化指數(MFI)變化情況Figure 2 Changes in myofibril fragmentation index of different samples during frozen storage
2.3 凍藏過程中肌原纖維表觀直徑變化
肌原纖維表觀直徑是用于量化肉制品在貯藏過程中肌原纖維被破壞程度的指標[2]20。如圖3所示,3種樣品D3,2均隨著凍藏時間的延長而下降,前2周凍藏過程中3組樣品之間無顯著性差異(P>0.05),而4~16周時3組樣品之間存在顯著性差異(P<0.05)。到達凍藏終點時,刀額新對蝦、整只和去頭羅氏沼蝦凍品的D3,2分別下降了31.94%,38.32%,34.42%。整個凍藏周期內整只刀額新對蝦凍品D3,2值始終高于其他樣品,且下降幅度最小。貯藏前期去頭羅氏沼蝦凍品D3,2值始終低于整只羅氏沼蝦凍品,后期隨著去頭羅氏沼蝦凍品D3,2下降幅度變小,在6周之后整只羅氏沼蝦凍品D3,2值反而低于去頭羅氏沼蝦凍品,這一趨勢與剪切力的變化類似。說明整只羅氏沼蝦凍品凍藏過程中肌原纖維表觀直徑下降的最為嚴重,肌原纖維破壞程度最大。林婉玲等[18]在研究浸漬凍結和靜止空氣凍結對凍藏凡納濱對蝦肌肉組織結構影響時,同樣發現隨著凍藏時間的延長,浸漬凍結和靜止空氣凍結的對蝦肌纖維直徑顯著變小。

圖3 凍藏過程中肌原纖維表觀直徑變化情況Figure 3 Changes inmyofibril apparent diameter of different samples during frozen storage
2.4 凍藏過程中TCA-溶解肽變化
TCA-溶解肽是反映內肽酶和微生物等對蛋白降解程度的指標之一[2]19,TCA-溶解肽含量越高,說明蛋白降解程度越高[19]。從圖4中可以看出,3組樣品在凍藏過程中,隨著凍藏時間延長TCA-溶解肽的含量逐漸升高,整體而言,整只羅氏沼蝦凍品前8周上升較快,8周后變化平穩,而去頭羅氏沼蝦凍品和整只刀額新對蝦凍品則在前2周增加速率較快,之后變化緩慢。整只刀額新對蝦凍品TCA-溶解肽的含量在凍藏周期內始終低于其他樣品,并且與羅氏沼蝦之間存在顯著性差異(P<0.01),可能是刀額新對蝦中內源酶系較羅氏沼蝦而言相對不發達。去頭羅氏沼蝦在去頭時直接破壞了細胞,內源酶釋放出來,另外,去頭過程中會受到微生物的污染[20],微生物代謝也會產生酶,各種酶的釋放和產生加速了蛋白的降解,可能是導致前期去頭蝦TCA-溶解肽含量在凍藏前2周增加較快的原因。在凍藏2周之后,整只羅氏沼蝦凍品中TCA-溶解肽逐漸高于去頭羅氏沼蝦凍品,并逐漸呈現顯著性差異(P<0.01),可能是整只蝦凍品蝦頭中的內源酶向蝦肉中遷移,使得蝦肉中蛋白酶活增大,蛋白降解程度增大。圖5中凍藏過程中不同羅氏沼蝦樣品總酶活的變化情況可以很好地說明酶的遷移并解釋2組羅氏沼蝦樣品TCA-溶解肽的變化情況。Cai等[21]和Jiang等[16]在真鯛魚片和草魚片冰藏過程中發現了相似的結果,TCA-溶解肽的含量顯著增加,表明樣品在貯藏過程中受到了內源酶的作用,導致肌原性含氮降解產物的增加,因而促進了微生物的生長和繁殖,加速樣品品質的劣化[22]。以上結果表明,凍藏期間整只羅氏沼蝦凍品TCA-溶解肽含量升高最為明顯,去頭羅氏沼蝦凍品TCA-溶解肽的產生相對緩慢。

圖4 凍藏過程中TCA-溶解肽變化情況Figure 4 Changes in TCA-soluble peptide of different samples during frozen storage
2.5 凍藏過程中總蛋白酶變化
由圖5可知,凍藏過程中,整只羅氏沼蝦凍品和去頭羅氏沼蝦第一、二腹節蝦肉中總蛋白酶活均有所增加,整個周期內整只羅氏沼蝦凍品總蛋白酶始終高于去頭羅氏沼蝦凍品,并且上升幅度大于去頭羅氏沼蝦凍品,而整只羅氏沼蝦凍品蝦頭內總蛋白酶活遠高于蝦肉中的蛋白酶活性,但呈逐漸下降趨勢,可能是蛋白酶從蝦頭中逐漸往蝦肉中遷移引起的。Benjakul等[23]發現凡納濱對蝦和羅氏沼蝦在冰藏2,4 d后蝦頭內的胰蛋白酶便會遷移到蝦肉的第一腹節內。Maria等[24]研究了紅鱒魚在-10~80 ℃貯藏18個月內組織蛋白酶D的活性變化,發現不同溫度貯藏對組織蛋白酶的活性影響不大。所以,整只羅氏沼蝦凍品在貯藏期間蝦頭內的內源酶發生遷移,從而使得酶活上升較快,進而可能促進了蛋白降解,同時也很好地解釋了TCA-溶解肽和MFI值等指標在貯藏后期劣化程度顯著高于去頭羅氏沼蝦凍品。
2.6 凍藏過程中菌落總數變化
圖6反映了3種樣品凍藏過程中菌落總數的變化情況。由圖6可知,隨著凍藏時間的延長,3種樣品的菌落總數均呈下降趨勢。在凍藏前2周菌落總數下降明顯,之后趨于緩和。這與李燕等[25]發現的羅氏沼蝦在-10 ℃凍藏過程中細菌總數在一定時間內隨著貯藏時間的延長不斷減少的現象一致。前2周菌落總數快速下降可能是速凍過程中溫度急劇下降,微生物的新陳代謝未能迅速重新調整,導致微生物的死亡率增加[26]。2周之后菌落總數仍處于下降趨勢,可能是在-18 ℃貯藏過程中,蛋白會發生降解和冷凍變性等,此外冰晶對微生物細胞也會產生機械破壞,以致微生物死亡率相應增加。在整個凍藏過程中去頭羅氏沼蝦的菌落總數始終高于整只羅氏沼蝦凍品,可能是在去頭過程中受到微生物的污染。凍藏過程中微生物數量逐漸下降,因此,可能微生物的作用對質構的影響較小。

圖5 凍藏過程中蝦肉總蛋白酶變化情況Figure 5 Changes in total proteolytic activity of different samples during frozen storage
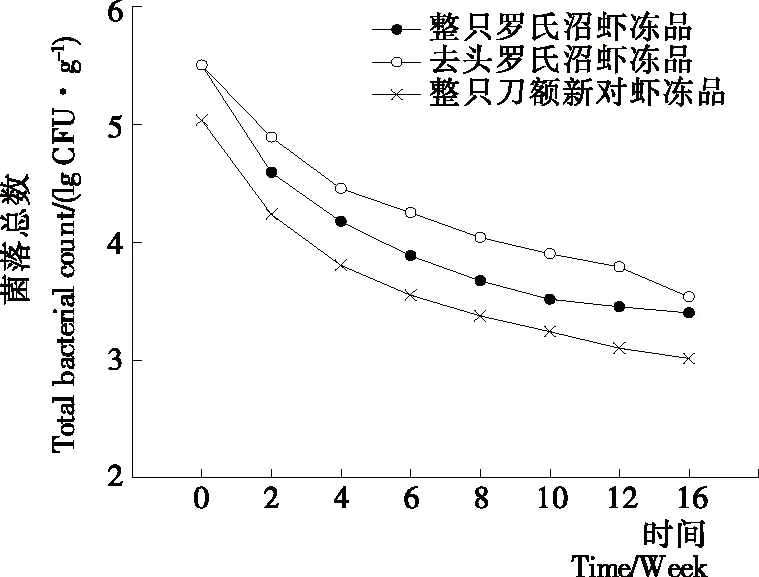
圖6 凍藏過程中菌落總數變化情況Figure 6 Changes in aerobic bacterial count of different samples during frozen storage
2.7 凍藏過程中微觀結構的變化
圖7、8分別為整只羅氏沼蝦凍品蝦肉橫斷面和縱斷面伊紅染色微觀組織結構的變化情況。新鮮的羅氏沼蝦蝦肉纖維在初期輪廓清晰,排列整齊,并且被一層薄的結締組織(肌內膜)包圍,呈不規則多邊形形狀。經過8周和16周的凍藏后,肌纖維出現明顯的變化,肌纖維內部間隙變大,肌纖維之間出現明顯分離。從縱斷面的結構圖可以看出,肌纖維在凍藏過程中發生了斷裂。Bahuaud等[27]研究了大西洋鮭魚在微凍貯藏過程中品質的變化,在對肌纖維的觀察過程中也發現了孔隙的出現,并指出這與冰晶的形成有直接關系。此外,有研究[28]表明,肌肉纖維是通過膠原纖維和結締組織相連接,貯藏過程中由于內源酶的作用,肌纖維和膠原纖維都會被降解,從而導致肌纖維和肌內膜之間脫離,進一步形成間隙。
2.8 相關性分析
對凍藏期間整只羅氏沼蝦凍品剪切力、MFI值、D3,2、TCA-溶解肽以及蝦肉中總蛋白酶活性等指標進行相關性分析,結果如表1所示。剪切力與MFI值、TCA-溶解肽和蝦肉中總蛋白酶活性呈負相關(P<0.01),與D3,2呈正相關(P<0.01),說明剪切力和蛋白劣化、蛋白酶活之間存在一定聯系;而MFI值與TCA-溶解肽和蝦肉中總蛋白酶活性呈正相關(P<0.01),D3,2與TCA-溶解肽和蝦肉中總蛋白酶活性呈負相關(P<0.01),說明酶活與蛋白劣化之間存在一定聯系,通過以上可以推斷,蝦肉中蛋白酶的作用加速了蛋白的降解,導致了蛋白各項指標的劣化,進而導致剪切力的下降。

圖7 凍藏過程中羅氏沼蝦微觀組織結構(橫斷面)的影響(放大倍數:200×)Figure 7 Changes in cross sections of muscle tissue of Macrobrachium rosenbergii during frozen storage (magnification: 200×)

圖8 凍藏過程中羅氏沼蝦微觀組織結構(縱斷面)的影響(放大倍數:200×)Figure 8 Changes in longitudinal sections of muscle tissue of Macrobrachium rosenbergii during frozen storage (magnification: 200×)
3 結論
整只羅氏沼蝦凍品蛋白劣化和蝦肉中蛋白酶活性明顯并高于去頭羅氏沼蝦凍品,且質構下降程度也較大。這可能是凍藏期間蝦頭內的內源蛋白酶隨著時間的延長遷移至蝦肉中并加速了蛋白的降解,從而使得MFI值和TCA-溶解肽等指標在凍藏后期變化較快,蛋白劣化嚴重并最終導致質構劣變程度較高。

表1 整只羅氏沼蝦凍品凍藏過程中剪切力、蛋白變化和蛋白酶活性變化等指標的相關性分析?Table 1 Pearson, correlation between changes in protein, protease activity and texture deterioration of Macrobrachium rosenbergii during frozen storage
本研究中主要就總酶活進行了分析,但在整個凍藏過程中內源酶到底是否發揮了降解蛋白的作用還未知,因此還需要通過進一步研究,深入分析內源酶在整只羅氏沼蝦凍品蛋白劣化和質構變化進程中的作用,深入揭示整只羅氏沼蝦凍品貯藏期間質構劣變機理。
