甲基營養型芽孢桿菌BH21對葡萄灰霉病菌的拮抗作用
2018-03-24 03:14:06魏新燕黃媛媛黃亞麗杜克久
中國農業科學 2018年5期
魏新燕,黃媛媛,黃亞麗,杜克久
?
甲基營養型芽孢桿菌BH21對葡萄灰霉病菌的拮抗作用
魏新燕1,黃媛媛2,黃亞麗2,杜克久1
(1河北農業大學林學院/國家北方山區農業工程技術研究中心,河北保定 071001;2河北省科學院生物研究所,石家莊 050051)
【目的】甲基營養型芽孢桿菌()BH21是一株對葡萄灰霉病菌()有較好拮抗作用的海洋源細菌,鑒定該菌株脂肽類抗菌物質合成基因,檢測脂肽粗提物對葡萄灰霉病菌的拮抗作用,為應用該菌株防治葡萄灰霉病提供科學依據。【方法】通過特異引物的基因組PCR法檢測甲基營養型芽孢桿菌BH21菌株合成脂肽的能力;鹽酸沉淀和甲醇抽提法從無菌發酵液中提取脂肽粗提物;排油圈法檢測脂肽粗提物的表面活性;采用菌絲生長速率法檢測脂肽粗提物對灰霉病菌菌絲生長的抑制作用并計算有效中濃度EC50;利用高效液相色譜技術(HPLC)對脂肽粗提物洗脫分離,并采用菌絲生長速率法檢測各組分對葡萄灰霉病菌的抑制能力;采用反相高效液相色譜(RP-HPLC)分析主要抑菌成分的脂肽類型。采用離體葉片接種法檢測脂肽粗提物對葡萄灰霉病的防治效果。【結果】選取11對特異引物對菌株BH21基因組擴增,其中7對引物擴增出預期核酸片段;擴增產物經測序、BLAST比對分析,結果顯示擴增產物與相關菌株脂肽基因相似度為96%—99%,擴增產物翻譯的蛋白與相關菌株的脂肽合成蛋白相似度為96%—100%,表明甲基營養型芽孢桿菌BH21基因組中含有、、、,該菌株具有合成surfactin、iturin及fengycin等多種脂肽類抗菌物質的能力。鹽酸沉淀和甲醇抽提從LB無菌發酵液中獲得脂肽粗提物,得率為428 mg·L-1。排油圈檢測結果顯示,脂肽粗提物使橄欖油膜形成排油圈,表明脂肽粗提物具有表面活性。脂肽粗提物對葡萄灰霉病菌菌絲生長具有顯著的抑制作用,脂肽粗提物濃度為440 μg·mL-1時,對葡萄灰霉病菌菌絲生長的相對抑制率為82.8%。根據毒力方程計算,抑制葡萄灰霉病菌菌絲生長的脂肽粗提物EC50為144.39 μg·mL-1。HPLC分離純化脂肽粗提物獲得6個組分,只有組分BH21-2和BH21-3抑制葡萄灰霉病菌的生長,RP-HPLC色譜圖分析表明組分BH21-2和BH21-3屬于fengycin家族脂肽。葡萄灰霉病離體葉片試驗結果表明,脂肽粗提物濃度為400 μg·mL-1時,對葡萄葉片灰霉病防病效果為100%;脂肽粗提物濃度為220 μg·mL-1時,對葡萄葉片病斑擴展相對抑制率為94.4%。【結論】甲基營養型芽孢桿菌菌株BH21具有合成surfactin、iturin及fengycin等多種脂肽類抗菌物質的基因,該菌株脂肽粗提物對葡萄灰霉病菌具有較強的拮抗作用,在葡萄灰霉病生物防治中具有應用潛力。
甲基營養型芽孢桿菌;葡萄灰霉病菌;抗真菌活性;脂肽;葡萄灰霉病
0 引言
【研究意義】葡萄灰霉病是由灰葡萄孢()引起的真菌病害,不僅在生長期危害,也是葡萄采后重要病害之一[1]。隨著化學農藥的長期、大量使用,其防控效果因葡萄灰霉病菌產生抗藥性而降低,積極開發微生物農藥具有實際應用價值[1-2]。甲基營養型芽孢桿菌()是芽孢桿菌屬的一個新成員[3],是植物根際促生菌,也是植物病害生物防治中重要組成部分[4-6]。甲基營養型芽孢桿菌BH21分離自滄州渤海海域,前期研究表明該菌株的無菌發酵液對葡萄灰霉病菌有較強的抑制作用(另文發表),明確其抑菌活性物質對該菌在灰霉病生物防治上的應用具有重要意義。【前人研究進展】伏波等[7]研究表明,小麥內生細菌枯草芽孢桿菌()Em7對葡萄灰霉病菌有良好的抑制作用,離體果實相對防治效果為78.92%;周泠璇等[8]從紅提葡萄中分離得到5株對葡萄灰霉病菌有較好拮抗作用的內生菌,其中內生菌枯草芽孢桿菌NR4-1對葡萄灰霉病菌抑菌效果最強,抑菌率為68.7%;Ge等[9]從長白山休眠火山土壤中分離到一株甲基營養型芽孢桿菌NKG-1,其不僅對番茄幼苗灰霉病具有60%的防治效果,且對番茄植株具有促生作用。芽孢桿菌產生多種抗菌化合物,其中環脂肽(LPs)是一類具有表面活性劑特性的抗菌物質,主要包括伊枯草菌素(iturin)、豐原素(fengycin)和表面活性素(surfactin)等3大類,具有抑菌、促生、誘導防御酶活性等特點[10]。Mora等[11]對64株芽孢桿菌進行了脂肽基因鑒定,結果表明48.4%的菌株含有2—4個脂肽基因,環脂肽是拮抗芽孢桿菌的主要抑菌活性物質;向亞萍等[12]對55株拮抗芽孢桿菌分泌的脂肽抗生素的定性定量分析結果表明,芽孢桿菌脂肽類抗生素是抑制植物病原真菌的主要成分,具有多樣性;iturin結構的變化及其分泌量與抑菌活性有相關性;呂倩等[13]從南海沉積樣品中分離出甲基營養型芽孢桿菌SHB114,其主要抗菌物質為iturin的bacillomycin Lc脂肽化合物;Liu等[14]發現從堿蓬根際分離的海洋芽孢桿菌()B-9987產生的抗菌物質是脂肽類物質,其中Marinhysin A是一種新結構的環脂肽類化合物;李德全等[15]驗證海藻中分離的枯草芽孢桿菌NH-8拮抗草莓灰霉病的主要活性物質為iturin。【本研究切入點】目前,灰霉病的生物防治研究主要集中在番茄[16-17]等草本植物,葡萄等木本植物灰霉病的生物防治研究相對較少,生防微生物大多來自陸生環境,海洋源甲基營養型芽孢桿菌在葡萄灰霉病防治中的應用鮮有報道。【擬解決的關鍵問題】鑒定甲基營養型芽孢桿菌BH21菌株脂肽類抗菌物質合成基因,檢測脂肽粗提物對葡萄灰霉病菌的拮抗作用,推測有效活性成分的脂肽類別,為防控植物灰霉病提供依據。
1 材料與方法
試驗于2016年在河北省科學院生物研究所和河北農業大學林木病理實驗室完成。
1.1 供試材料
灰霉病菌菌株B-20由河北農業大學從罹病葡萄葉片上分離、鑒定,在PDA培養基上培養和保存。
拮抗芽孢桿菌BH21菌株分離于滄州渤海灣海洋樣品,經形態特征、生理生化、16S rDNA和基因序列分析,鑒定為甲基營養型芽孢桿菌,保藏于中國微生物菌種保藏委員會普通微生物菌種保藏中心(CGMCC No. 11840)。該菌株具有耐高鹽、高濃度甲醇的特性,對灰霉菌具有較強的拮抗作用。BH21菌株菌懸液與25%甘油按1﹕1比例混合,保存于-80℃超低溫冰箱。
供試葡萄品種為維多利亞(Victoria),采自河北省衡水市饒陽縣。
PDA培養基[8]、LB培養基[18]、ZoBell’s 2216E培養基[19]:蛋白胨5 g,酵母膏1 g,FePO40.1 g,瓊脂粉12 g,陳海水定容至1 L,pH 7.6。
1.2 菌株培養
將保存于4℃的灰霉病菌以菌餅接種于PDA培養基平板中心,(23±1)℃黑暗培養5 d,備用。用接種環取一環保存于-80℃冰箱的BH21菌株,接種于ZoBell’s 2216E培養基,(23±1)℃黑暗培養24 h,活化備用。
1.3 芽孢桿菌產脂肽基因檢測
根據Cao等[20]方法,分別合成ituA1F/ituA1R、bamB1F/bamB1R、bamC2F/bamC2R、ITUCF1/ITUCR3、ituD2F/ituD2R、FenB1F/FenB1R、FNDF1/FNDR1、110F/110R、Sbo1F/Sbo1R、Qk1F/Qk1R和147F/147R等11對引物,并按照各引物擴增程序進行PCR擴增。反應體系為25 μL:10×PCR Buffer 2.5 μL,2.5 mmol·L-1dNTP 2.0 μL,Taq DNA聚合酶1.25 U,DNA模板(菌懸液)1.0 μL,25 μmol·L-1上下游引物各0.5 μL,加ddH2O至總體積25 μL。PCR產物用1%的瓊脂糖凝膠電泳,染色劑為GoldView。凝膠成像系統GDS-8000檢測,并記錄檢測結果。將瓊脂糖凝膠電泳檢測符合要求的PCR產物交由上海立菲生物技術有限公司純化測序。將所得核酸序列通過NCBI的比對工具BLAST進行在線比對。
1.4 脂肽粗提物的提取和表面活性檢測
菌株BH21培養:LB培養基、100 mL/250 mL三角瓶裝量、28℃、180 r/min恒溫搖床振蕩培養72 h。發酵液經8 000 r/min離心20 min,取上清發酵液用孔徑為0.22mm無菌過濾器過濾,獲得無菌發酵液。采取鹽酸沉淀和甲醇抽提法從無菌發酵液中獲得脂肽粗提物[13]。采用排油圈法[21]檢測粗提物表面活性。將粗提物配置成10 mg·mL-1的甲醇溶解液,用孔徑為0.22mm無菌過濾器過濾。取適量橄欖油,加入蘇丹黑Ⅲ染色劑,混勻。取直徑9 cm的潔凈培養皿,加入60 mL純凈水,緩慢加入染色的橄欖油使水面形成一層油膜。用移液器吸取脂肽粗提物的甲醇溶解液,向培養皿油膜中心持續滴加,看有無排油圈形成。以滴加甲醇的為對照。
1.5 脂肽粗提物對葡萄灰霉病菌菌絲生長的影響
采用菌絲生長速率法[8]測定脂肽粗提物的生物活性。用無菌水配置成初濃度為22 mg·mL-1的脂肽溶液,并將該水溶液加入融化的PDA培養基中制成脂肽濃度為440、220、110、55 μg·mL-1的脂肽平板,以加等量無菌水的PDA平板為對照。在平板中央接種直徑為8 mm的灰霉病菌菌塊,每處理重復3次,23℃培養4 d。采用十字交叉法測量菌落直徑,計算抑制率。相對抑制率(%)=[(對照菌落直徑-脂肽粗提物處理菌落直徑)/對照菌落直徑]×100。根據濃度對數()與菌落生長相對抑制率的概率值(),利用Excel軟件求取脂肽粗提物對灰霉病菌菌絲生長的毒力回歸方程=a+b和相關系數(),并計算抑制菌落生長的有效中濃度EC50。
1.6 脂肽粗提物對離體葡萄葉片灰霉病的防治效果
采用離體組織接種法[22],用無菌水分別制成440、220、110、55、27.5 μg·mL-1的脂肽溶液。選取大小相近葡萄健康葉片,分別用不同濃度的脂肽溶液將葡萄葉片兩面浸濕5 min,晾干,以無菌水處理的葡萄葉片為對照。在葉片正面的3個不同部位接種直徑為0.8 cm的灰霉病菌菌餅(培養7 d,具有大量孢子)。22℃保濕4 d,記錄發病情況及病斑直徑,利用SPSS軟件分析防治效果。
1.7 脂肽粗提物洗脫分離及抑菌檢測
1.7.1 脂肽粗提物洗脫分離 采用高效液相色譜進行BH21產生的脂肽物質的洗脫分離,色譜柱為TricornTM10/300 column;流動相:A液95%去離子水,5%乙腈,0.1%三氟乙酸,B液5%去離子水,95%乙腈,0.1%三氟乙酸;洗脫條件:0.1% HFA乙腈,0→100% 線性洗脫120 min;檢測波長:215 nm;流速:3 mL·min-1;進樣量:10 mL;柱溫:室溫。使用之前將A液和B液超聲處理30 min,以去除液體中的氣泡。
1.7.2 分離物各組分對灰霉病菌的抑制作用 根據各組混合物洗脫時,A液和B液所占比例,配置相對應的洗脫液(均值)作為對照。采用瓊脂擴散法檢測分離物各組分對灰霉病菌的抑制作用。具體操作:向PDA平板中央接種直徑為8 mm的灰霉病菌菌餅,距中心3 cm處打孔,分別加入50 μL洗脫液和相對溶劑,(23±1)℃黑暗培養。
1.7.3 脂肽分離物有效組分分析 將對灰霉病菌有抑制作用的脂肽分離物通過反相高效液相色譜分析。色譜柱:waters Xbridge Peptide BEHC18,300 A,5 μm;流動相:0.1% HFA Water,0.1% HFA乙腈;洗脫條件:0.1% HFA乙腈,0→100% 梯度洗脫80 min;檢測波長:215 nm;流速:1 mL·min-1;進樣量:1 mL;柱溫:30℃。
2 結果
2.1 BH21菌株脂肽基因的檢測
2.1.1 Iturin合成的相關基因序列分析 引物ituA1F/ituA1R、bamB1F/bamB1R、ITUCF1/ITUCR3和ituD2F/itud2R的PCR分別得到880、512、560、643 bp核酸片段,與Cao等[20]報道的大小相近。Nucleotide blast工具分析結果表明,4對引物獲得的核酸序列分別與inturin合成基因、、、相關,與相關菌株相似度為97%—99%(表1)。核酸序列產物分別為iturin A合成酶A、bacillomycin D合成蛋白BmyB、iturin A合成酶C、D,與相關菌株相似度為96%—98%。
2.1.2 Fengycin合成的相關基因序列分析 引物FNDF1/FNDR1擴增獲得288 bp核酸片段,Nucleotide blast工具分析結果表明,此核酸片段與解淀粉芽孢桿菌(subsp.)SQR9的相似度為99%(表1),其產物是fengycin合成酶FenD。FNDF1/FNDR1擴增產物翻譯的蛋白序列與枯草芽孢桿菌MBI600FenD蛋白序列的相似度為100%。
2.1.3 Sufactin合成的相關基因序列分析 引物110F/110R擴增獲得307 bp核酸片段,此核酸片段與解淀粉芽孢桿菌SQR9的相似度為96%(表1),其產物是sufactin合成酶SrfAB。110F/110R擴增產物翻譯的蛋白序列與枯草芽孢桿菌SQR9的SrfAB蛋白序列相似度為96%。
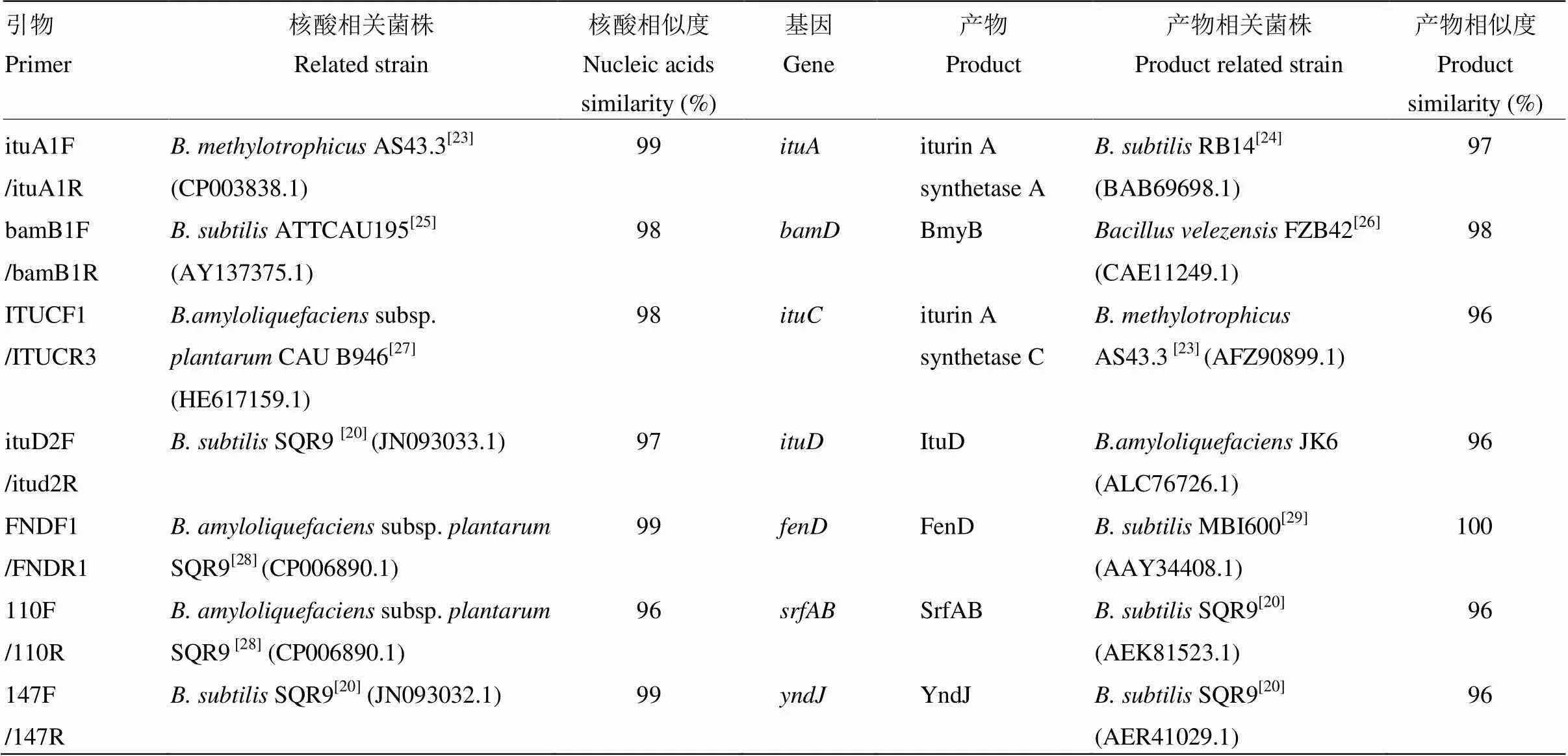
表1 PCR產物核酸序列與相關基因的相似度
2.1.4 假定蛋白YndJ合成的相關基因序列分析 引物147F/147R擴增獲得215 bp核酸片段,此核酸片段與枯草芽孢桿菌SQR9的相似度為99%(表1),其產物是假定蛋白合成酶YndJ。擴增產物翻譯的蛋白序列與枯草芽孢桿菌SQR9的YndJ蛋白序列相似度為96%。
2.2 脂肽粗提物的提取和表面活性檢測
采用鹽酸沉淀和甲醇抽提法從菌株BH21發酵液中提取脂肽粗提物,冷凍干燥后確定該粗提物的得率為428 mg·L-1。滴加該粗提物的甲醇溶解液可以在橄欖油油膜上形成排油圈,而甲醇對照則不能產生排油圈,進一步驗證該提取物為脂肽類物質,具有較強的表面活性。
2.3 BH21脂肽粗提物對葡萄灰霉病菌菌絲生長的抑制作用
葡萄灰霉病菌菌塊在含不同濃度BH21脂肽粗提物的PDA培養基上23℃培養4 d后,與對照相比,試驗濃度的BH21脂肽物質均對灰霉病菌菌絲生長有一定的抑制作用,而且隨著脂肽濃度的增加,菌絲生長受到的抑制作用增強,其中440 μg·mL-1脂肽物質對灰霉病菌菌絲生長的抑制率為82.8%(表2)。根據濃度對數()與菌落生長相對抑制率的概率值(),菌株BH21脂肽粗提物對灰霉菌菌絲生長抑制的毒力方程為=1.097+3.1157,相關系數(2=0.921),根據此方程計算菌株BH21產生的脂肽類混合物對灰霉病菌菌絲抑制的EC50為144.39 μg·mL-1。
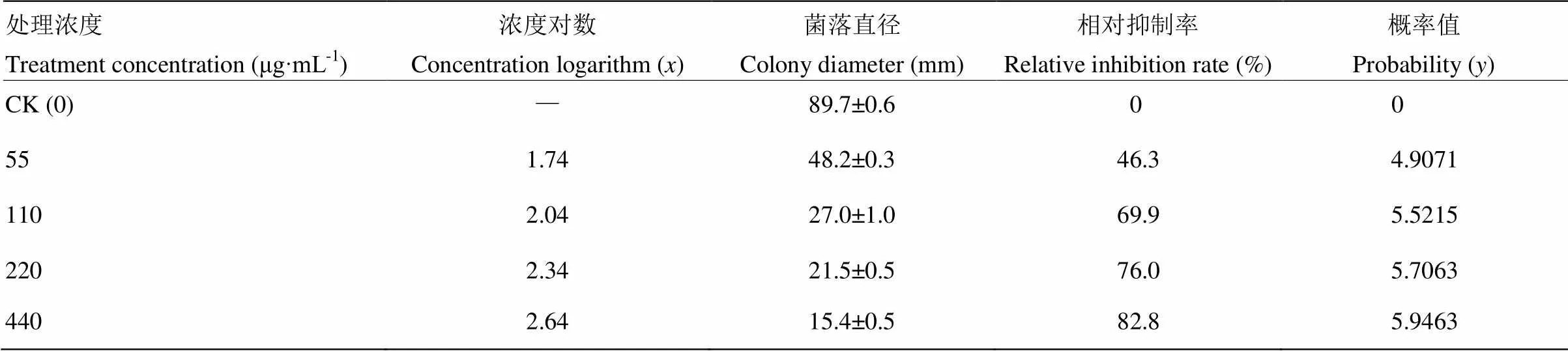
表2 BH21脂肽粗提物對灰霉病菌菌絲生長的抑制作用
2.4 BH21脂肽粗提物對離體葡萄葉片灰霉病的防治效果
采用不同濃度的BH21脂肽粗提物稀釋液處理葡萄葉片,保濕培養4 d后進行灰霉病病斑的測定(圖1、表3)。與對照相比,不同濃度的脂肽粗提物稀釋液對葡萄葉片灰霉病都有明顯防治效果,脂肽粗提物濃度越高,病斑直徑越小,其中濃度為440 μg·mL-1的BH21脂肽溶液的防治效果最好,接種部位不發病。根據病斑直徑計算BH21脂肽粗提物對灰霉病病斑擴展相對抑制率為32.8%—100%,防治效果顯著(表3)。

1: 440 μg·mL-1; 2: 220 μg·mL-1; 3: 110 μg·mL-1; 4: 55 μg·mL-1; 5: 27.5 μg·mL-1

表3 BH21脂肽粗提物對離體葡萄葉片灰霉病病斑數和病斑擴展的影響
同列數據后不同小寫字母表示在<0.05水平差異顯著
Data with different lowercases in the same column indicate significant difference at<0.05 level
2.5 菌株BH21脂肽粗提物的分離及抑菌效果檢測
2.5.1 脂肽粗提物洗脫分離 根據制備型液相色譜的峰值,BH21脂肽粗提物中共有6個明顯的單峰,根據峰值共收集了6個樣品,依據收集順序分別命名為組分1、組分2、……、組分6。根據各峰出現時洗脫液的成分比例,計算得到相應洗脫液的平均值(表4)。
2.5.2 各組分對灰霉病菌生長的抑制作用 接種培養4 d后,各組分及其相對應的洗脫液對灰霉菌菌絲的生長均有一定的抑制作用,并且洗脫液的揮發也使灰霉病菌生長受到抑制。組分1、4、5、6與其相對應的洗脫液1、4、5、6對灰霉菌絲的抑制作用基本相同,說明這4個成分無抑菌活性。組分2和組分3對灰霉菌抑制作用明顯大于其相應的緩沖液(圖2),其中組分2的抑菌帶寬度為(6.3±0.6)mm,組分3的抑菌帶寬度為(2.8±0.3)mm,由此判斷組分2和組分3在菌株BH21對灰霉病菌的拮抗中起主要作用,且組分2的抑菌活性大于組分3,將這兩個活性成分分別命名為BH21-2和BH21-3。
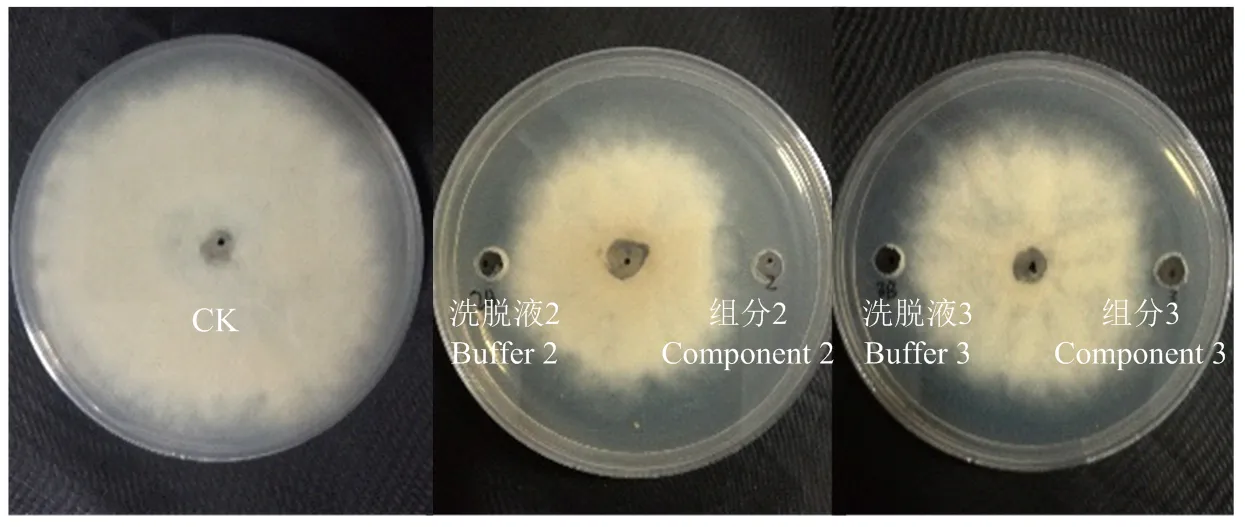
圖2 脂肽粗提物分離物對灰霉病菌的拮抗活性
2.5.3 脂肽粗提物分離物BH21-2和BH21-3的反相高效液相色譜檢測 將組分BH21-2和BH21-3分別進行RP-HPLC檢測,結果表明組分BH21-2、BH21-3保留時間范圍分別在43—51、48—55 min,兩個組分各包含多個小峰,兩者之間還有部分重疊(圖3)。此出峰時間與相似條件下fengycin類脂肽出峰時間相符(34—56 min)[30],因此推斷組分BH21-2和BH21-3均為fengycin類脂肽。
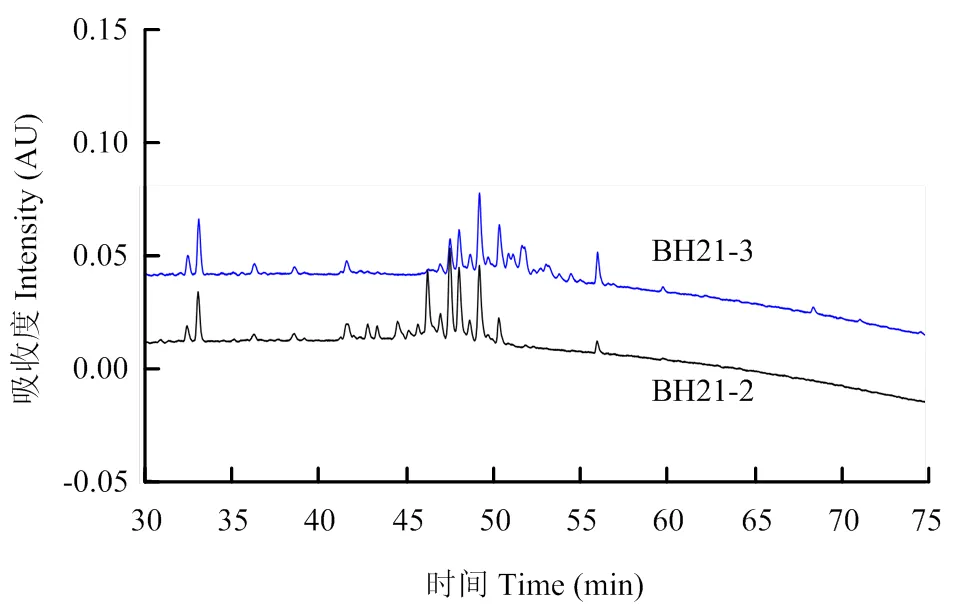
圖3 BH21-2和BH21-3反相高效液相色譜圖
3 討論
灰霉病生防菌株的篩選一直是灰霉病安全防控研究的重點,隨著陸地拮抗微生物篩選和研究的深入,人們將目光轉向生境獨特的海洋微生物資源[31]。高偉等[32]研究發現,從渤海潮間帶鹽生植物堿蓬根際分離的海洋芽孢桿菌B-9987對番茄灰霉病具有良好的拮抗作用;李德全等[15]從南通近海海藻中分離到枯草芽孢桿菌NH-8,該菌能夠增強草莓系統抗性,促進草莓植株生長。然而,由于活體微生物資源的應用效果受外界條件影響較大,對拮抗菌株所產生的抑菌活性物質的分離、鑒定具有更為重要的意義。其中脂肽是芽孢桿菌產生的一類重要抑菌活性物質,呂倩等分別從海洋細菌發酵液中獲得了具有抑菌活性的bacillomycin Lc和Marinhysin A等脂肽類物質[13-14]。李德全等[15]的研究發現,海洋枯草芽孢桿菌NH-8中的主要抗菌物質為iturin,其粗提物對灰霉菌的抑菌率為82.7%。本研究的拮抗菌株BH21是從滄州渤海灣海洋樣品中分離的甲基營養型芽孢桿菌,其菌體和無菌發酵液對灰霉病菌均具有較強的拮抗作用,表明該菌株具有產生胞外抑菌物質的能力,初步研究表明該發酵液具有明顯的表面活性,推測該菌產生的活性物質有可能為脂肽類物質(另文發表)。
本研究采用鹽酸沉淀和甲醇抽提的方法進行BH21發酵液中的脂肽類物質的分離,通過平板和離體葉片試驗方法測定了不同濃度脂肽粗提物對灰霉病菌生長和葉面病害的防治能力。離體試驗是能夠快速、準確反應菌株拮抗效果的試驗方法,在拮抗菌株和抑菌物質篩選中多被采用[33-34]。本研究為了盡快確定BH21脂肽類粗提物的抑菌效果,采用葡萄離體葉片測定了脂肽粗提物對灰霉病的防治效果,在接種灰霉菌塊的情況下,不同濃度的粗提物均對灰霉病有良好的防治作用,400 μg·mL-1稀釋液對離體葡萄葉片的灰霉病防治效果為100%、55 μg·mL-1時的防治效果為55.0%,防病效果好于相關研究結果[15]。
對菌株產生的抑菌活性物質進行進一步純化和鑒定是明確菌株拮抗機理的基礎,對菌株脂肽類合成基因的檢測可以確定菌株存在何種物質的合成能力。研究發現,脂肽類物質主要包括surfactin、iturin和fengycin等3大類,隨著對這些物質合成基因研究的深入,研究者設計出多對特異性引物以進行菌株脂肽類物質的合成能力和種類的測定[35]。Cao等[20]應用已發表和自己設計的引物進行PCR擴增,鑒定出枯草芽孢桿菌SQR9中含有、、、、、、、和等9個脂肽合成基因;Mora等[11]則利用特異引物探針篩選出具有脂肽合成基因的拮抗菌株。本研究采用PCR方法并利用11對特異引物檢測菌株BH21中是否存在脂肽物質的合成基因,確定菌株BH21含有、、、、、和,推測該菌株具有合成多種脂肽類物質的能力。此外,在選擇的11對引物中只有7對引物擴增出與目標大小相近的產物,未得到擴增產物的4對引物分別為Sbo1F/Sbo1R(枯草芽孢桿菌素的)、FenB1F/ FenB1R(豐原素的)、bamC2F/bamC2R(芽孢菌霉素的)、Qk1F/Qk1R(枯草桿菌蛋白酶)。這4對引物雖多次調整擴增程序或始終無擴增產物或產生非特異性片段,推測其沒有特異產物的原因,其一可能由于菌株BH21沒有相似的基因序列,也可能是基因序列與特異引物不匹配,而后者則有可能是由于海洋細菌中含有特異成分所引起的。
那么,菌株BH21發酵液的甲醇粗提物中拮抗灰霉病菌的脂肽物質有幾類呢?本研究通過制備型HPLC進行了脂肽粗提物的分離,并將不同出峰時間的脂肽分離物進行抑制灰霉病菌的活性驗證。對峙試驗結果表明組分2和組分3具有明顯的抑菌活性,其中組分2抑菌能力最強。反相高效液相色譜圖顯示組分2和組分3與fengycin類脂肽[29]出峰時間相符合,特異引物PCR也驗證菌株BH21含有fengycin類脂肽合成基因。然而,組分2和組分3中均含有多個小峰,其分子結構仍需采用核磁共振等方法進行鑒定。另外,其他4種組分的功能性質、菌株所具有的iturin和surfactin類脂肽合成基因是否表達需要進一步研究。
甲基營養型芽孢桿菌SHB114[13]是來自南海深海沉積物的一株對黃瓜炭疽病菌()有較強抑制作用的生防菌株,其產生的脂肽為iturins類的bacillomycin LC。本文的菌株BH21同樣是一株海洋源甲基營養型芽孢桿菌,其對葡萄灰霉病菌有效的活性成分是fengycin類脂肽,不僅為葡萄灰霉病的生物防控提供了備選資源,同時也擴展了海洋源甲基營養型芽孢桿菌的抑菌范圍。
4 結論
來自海洋環境的甲基營養型芽孢桿菌菌株BH21具有產生多種抗菌脂肽的潛力,發酵液脂肽粗提物對葡萄灰霉病菌具有較強的拮抗作用,其有效活性成分為fengycin類脂肽,在葡萄灰霉病生物防治中具有潛在應用價值。
[1] 張瑋, 喬廣行, 黃金寶, 王忠躍, 李興紅. 中國葡萄灰霉病菌對嘧霉胺的抗藥性檢測. 中國農業科學, 2013, 46(6): 1208-1212.
Zhang W, Qiao G X, Huang J B, Wang Z Y, Li X H. Evaluation on resistance of grape gray mold pathogento pyrimethanil in China.,2013, 46(6): 1208-1212. (in Chinese)
[2] 嚴紅, 燕繼曄, 王忠躍, 李亞寧, 金桂華, 李興紅. 葡萄灰霉病菌對3種殺菌劑的多重抗藥性檢測. 果樹學報, 2012, 29(4): 625-629.
Yan H, Yan J Y, Wang Z Y, Li Y N, Jin G H, Li X H. Multiple fungicide resistance offrom grapevine to three fungicides., 2012, 29(4): 625-629. (in Chinese)
[3] Madhaiyan M, Poonguzhali S, Kwon S W, Sa T M.sp. nov., a methanol-utilizing, plant- growth-promoting bacterium isolated from rice rhizosphere soil., 2010, 60(10): 2490-2495.
[4] 王洪梅, 吳云成, 沈標. 青枯病生防菌N5的特性及其生物學效應. 土壤, 2013, 45(6): 1082-1090.
Wang H M, Wu Y C, Shen B.Characterization and biological effects of antagonistic N5 against., 2013, 45(6): 1082-1090. (in Chinese)
[5] 劉偉, 宋雙, 沈小英, 安天賜, 牛小義, 安德榮. 番茄灰霉病拮抗芽孢桿菌LW-6-1的篩選、鑒定及抑菌活性研究. 西北農林科技大學學報(自然科學版), 2013, 41(11): 73-79.
Liu W, Song S, Shen X Y, An T C, Niu X Y, An D R. Screening, identification and antibacterial activity of antagonistic bacteria LW-6-1 against., 2013, 41(11): 73-79. (in Chinese)
[6] 黃霄, 陳波, 周登博, 譚昕, 張錫炎. 菌株BM-24的分離鑒定及對香蕉枯萎病菌的抑菌活性. 植物保護學報, 2013, 40(2): 121-127.
Huang X, Chen B, Zhou D B, Tan X, Zhang X Y.Isolation, identification of BM-24 strains and its antifungal activity of antagonistic bacteria againstf. sp.., 2013, 40(2): 121-127. (in Chinese)
[7] 伏波, 姚娟妮, 高小寧, 黃麗麗, 康振生, 韓青梅. 植物內生枯草芽孢桿菌Em7菌株對葡萄灰霉病菌的抑菌活性. 農藥學學報, 2016, 18(4): 465-471.
FU B, YAO J N, GAO X N, HUANG L L, KANG Z S, HAN Q M. Antifungal activity of plant endophyticstrain Em7 against., 2016, 18(4): 465-471. (in Chinese)
[8] 周泠璇, 劉婭. 紅提葡萄內生細菌的分離鑒定及灰霉病拮抗菌的篩選. 生物技術通報, 2016, 32(4): 184-189.
Zhou L X, Liu Y.Isolation and identification of endophytic bacteria in red grape, and screening of antagonistic bacteria against., 2016, 32(4): 184-189. (in Chinese)
[9] Ge B B, Liu B H, Nwet T T, zhao W J, Shi L M, ZHANG K.strain NKG-1, isolated from Changbai Mountain, China, has potential applications as a biofertilizer or biocontrol agent., 2016,11(11): e0166079.
[10] Ongena M, Jacques P.lipopeptides: versatile weapons for plant disease biocontrol., 2008, 16(3): 115-125.
[11] Mora I, Cabrefiga J, Montesinos E. Cyclic lipopeptide biosynthetic genes and products, and inhibitory activity of plant- associatedagainst phytopathogenic bacteria., 2015, 10(5): e0127738.
[12] 向亞萍, 陳志誼, 羅楚平, 周華飛, 劉永鋒.芽孢桿菌的抑菌活性與其產脂肽類抗生素的相關性. 中國農業科學, 2015, 48(20): 4064-4076.
Xiang Y P, Chen Z Y, Luo C P, Zhou H F, Liu Y F.The antifungal activities ofspp. and its relationship with lipopeptide antibiotics produced byspp.., 2015, 48(20): 4064-4076. (in Chinese)
[13] 呂倩, 胡江春, 王楠, 王雪梅, 王書錦. 南海深海甲基營養型芽孢桿菌SHB114抗真菌脂肽活性產物的研究. 中國生物防治學報, 2014, 30(1): 113-120.
Lü Q, Hu J C, Wang N, Wang X M, Wang S J. Anti-fungal lipopeptides produced bySHB114 isolated from south China sea., 2014, 30(1): 113-120. (in Chinese)
[14] Liu R F, Zhang D J, Li Y G, Li M T, Li T. A new antifungal cyclic lipopeptide fromB-9987., 2010, 93(12):2419-2425.
[15] 李德全, 錢亞明, 周鳴鳴, 談蓉, 鄧自發, 袁素霞. 海洋細菌NH-8防治草莓灰霉病機理及其抗菌物質分析. 植物保護學報, 2016, 43(2): 215-221.
Li D Q, Qian Y M, Zhou M M, Tan R, Deng Z F, Yuan S X. The mechanism of biological control of strawberry gray mould using the marine bacterial NH-8 strain and analysis of the antifungal substances from the strain., 2016, 43(2): 215-221. (in Chinese)
[16] 趙楊, 苗則彥, 李穎, 白元俊. 番茄灰霉病防治研究進展. 中國植保導刊, 2014, 34(7): 21-29.
Zhao Y, Miao Z Y, Li Y, Bai Y J. Research progress on controlling against tomato gray mold., 2014, 34(7): 21-29. (in Chinese)
[17] 陳宇飛, 文景芝, 李立軍. 葡萄灰霉病研究進展. 東北農業大學學報, 2006, 37(5): 693-699.
Chen Y F, Wen J Z, Li L J. Research advance of grape grey mould., 2006, 37(5): 693-699. (in Chinese)
[18] 方中達. 植病研究方法. 北京: 中國農業出版社, 1998.
FANG Z D.. Beijing: China Agriculture Press, 1998. (in Chinese)
[19] 孫菽蔚, 王子峰, 岳海東, 肖天. 一株海洋幾丁質酶產生菌的篩選及其產酶條件的初步研究. 海洋科學, 2007, 31(5): 10-16.
Sun S W, Wang Z F, Yue H D, Xiao T. Screening of chitinase-producing marine bacterial strains and preliminary studies on chitinase-producing conditions., 2007, 31(5): 10-16. (in Chinese)
[20] Cao Y, Xu Z H, Ling N, Yuan Y J, Yang X M, Chen L H, Shen B, Shen Q R. Isolation and identification of lipopeptides produced bySQR 9 for suppressing Fusarium wilt of cucumber., 2012, 135(1): 32-39.
[21] 曹小紅, 廖振宇, 王春玲, 哈志瑞, 楊亞靜, 魯梅芳, 勵建榮.TK-1產脂肽的純化、抑菌活性及其表面活性劑特性. 中國生物工程雜志, 2008, 28(1): 44-48.
Cao X H, Liao Z Y, Wang C L, Ha Z R, Yang Y J, Lu M F, Li J R. Purification and antimicrobial activity of lipopeptide produced byTK-1 and its surfactant property., 2008, 28(1): 44-48. (in Chinese)
[22] 黃大野, 周婷, 姚經武, 劉曉艷, 曹春霞, 楊妮娜, 胡洪濤, 龍同, 楊自文. 死亡谷芽孢桿菌NBIF-001防治灰霉病研究. 中國蔬菜, 2016(10): 63-66.
Huang D Y, ZHOU T, Yao J W, Liu X Y, Cao C X, Yang N N, Hu H T, Long T, Yang Z W. Studies on controlling effect ofNBIF-001 on gray mold., 2016(10): 63-66. (in Chinese)
[23] Dunlap C A, Kim S J, Kwon S W, ROONEY A P. Phylogenomic analysis shows thatsubsp.is a later heterotypic synonym of., 2015, 65(7): 2104-2109.
[24] Tsuge K, Akiyama T, Shoda M. Cloning, sequencing, and characterization of the iturin A operon., 2001, 183(21): 6265-6273.
[25] Moyne A L, Cleveland T E, Tuzun S. Molecular characterization and analysis of the operon encoding the antifungal lipopeptide bacillomycin D., 2004, 234(1): 43-49.
[26] Koumoutsi A, Chen X H, Henne A, Liesegang H, Hitzeroth G, Franke P, Vater J, Borriss R. Structural and functional characterization of gene clusters directing nonribosomal synthesis of bioactive cyclic lipopeptides instrain FZB42., 2004, 186(4): 1084-1096.
[27] Blom J, Rueckert C, Niu B, WANG Q, BORRISS R. The complete genome ofsubsp.CAU B946 contains a gene cluster for nonribosomal synthesis of iturin A., 2012, 194(7): 1845-1846.
[28] Zhang N, Yang D Q, Wang D D, Miao Y Z, Shao J H, Zhou X, Xu Z H, Li Q, Feng H C, Li S Q, Shen Q R, Zhang R F. Whole transcriptomic analysis of the plant-beneficial rhizobacteriumSQR9 during enhanced biofilm formation regulated by maize root exudates., 2015, 16(1): 685.
[29] Joshi R, Gardener B B M. Identification and characterization of novel genetic markers associated with biological control activities in., 2006, 96(2): 145-154.
[30] Arrebola E, Jacobs R, Korsten L. Iturin A is the principal inhibitor in the biocontrol activity ofPPCB004 against postharvest fungal pathogens., 2010, 108(2): 386-395.
[31] 劉淼, 王繼紅, 姜健, 楊寶靈, 溫小紅, 陳玉飛, 劉麗. 海洋微生物應用于生物農藥的研究進展. 中國農學通報, 2014, 30(3): 232-236.
Liu M, Wang J H, Jiang J, Yang B L, Wen X H, Chen Y F, Liu L.Advances in the studies on the application of marine microorganism in biological pesticides., 2014, 30(3): 232-236. (in Chinese)
[32] 高偉, 田黎, 張久明, 周俊英, 鄭立, 崔志松, 李元廣. 海洋芽孢桿菌B-9987菌株對番茄灰霉病和早疫病的作用機制初探. 植物保護, 2010, 36(1): 55-59.
Gao W, Tian L, Zhang J M, Zhou J Y, Zheng L, Cui Z S, Li Y G. A primary study on the biocontrol mechanisms ofB-9987 against the tomato gray mold and early blight., 2010, 36(1): 55-59. (in Chinese)
[33] 鞏文峰, 李月飛, 上官妮妮, 趙新貝, 王陽, 馬青. 出芽短梗霉對蘋果采后灰霉病的防治. 中國生物防治學報, 2016, 32(2): 251-257.
Gong W F, Li Y F, Shangguan N N, Zhao X B, Wang Y, Ma Q. Control of apple postharvest gray mold by., 2016, 32(2): 251-257. (in Chinese)
[34] 申順善, 張瑩瑩, 張維娜, 呂雅悠, 朱卓琳, 樸鳳植. 綠針假單胞菌HL5-4對番茄灰霉菌的抑制活性及其定殖能力. 園藝學報, 2016, 43(6): 1195-1202.
Shen S S, Zhang Y Y, Zhang W N, Lü Y Y, Zhu Z L, Piao F Z. Antifungal activity ofHL5-4 against tomato gray mold and its colonization ability., 2016, 43(6): 1195-1202.(in Chinese)
[35] Hsieh F C, Lin T C, Meng M, Kao S S. Comparing methods for identifyingstrains capable of producing the antifungal lipopeptide iturin A., 2008, 56(1): 1-5.
(責任編輯 岳梅)
Antagonism ofstrain BH21 to
WEI XinYan1, HUANG YuanYuan2, HUANG YaLi2, DU KeJiu1
(1College of Forestry, Hebei Agricultural University/National Engineering Research Center for Agriculture in Northern Mountainous Areas, Baoding 071001, Hebei;2Institute of Biology, Hebei Academy of Sciences, Shijiazhuang 050051)
【Objective】strain BH21 is a marine derived bacterium that has a good antagonistic effect on. The objective of this study is to identify the lipopeptide synthesis genes of the strain BH21 and investigate the antagonism of the crude extracts of lipopeptide to, and to provide a scientific basis for the prevention and control of. 【Method】To determine the mechanisms of the antagonistic strain, PCR was used to screen strain BH21 for genes involved in biosynthesis of antimicrobial lipopeptide. Crude lipopeptide was extracted from the culture broth by hydrochloric acid precipitation and methanol extraction. surface activity of the crude lipopeptide was determined by oil spreading method. The inhibition ability of the crude lipopeptide on mycelial growth ofwas investigated by mycelial growth rate method and EC50was calculated. The crude lipopeptide was separated by liquid chromatography (HPLC) and the inhibition ability of each component towas detected by the mycelium growth rate method. Reversed phase high performance liquid chromatography (RP-HPLC) was used to analyze the types of the main antifungal components. the effect of the crude lipopeptide on the control ofin grape was detected by tissue inoculation. 【Result】A total of 11 pairs of specific primers were used for genomic amplification of strain BH21, and 7 gene fragments of the size expected to be correlated with biocontrol activities were efficiently amplified. after amplification, sequencing and BLAST analysis, the results showed that the similarity between the amplified products and the related lipopeptide genes was 96%-99%, the similarity between the protein produced by the nucleic acid fragment and the lipopeptide synthesized protein of the related strain was 96%-100%, which showed that the genome of strain BH21 contained,,,,,andgenes and the strain had the ability to synthesize antimicrobial lipopeptide such as surfactins, iturins and fengycins. Antifungal lipopeptide produced by BH21 was extracted by hydrochloric acid precipitation and methanol extraction, and the yield was 428 mg·L-1. The results of the oil spreading test showed that the crude lipopeptide had surface activity. The crude lipopeptide significantly inhibited mycelial growth ofwhen theconcentration was 440 μg·mL-1, the relative inhibition rate of mycelial growth ofwas 82.8%, and the effective medium concentration EC50was 144.39 μg·mL-1. Six fractions were collected with elution time through HPLC, only BH21-2 and BH21-3 inhibited the growth of. RP-HPLC chromatogram analysis showed that the components BH21-2 and BH21-3 belong to the fengycin family. Grape leaftest results showed that when the concentration of crude lipopeptide was 440 μg·mL-1,the control effect againstgrapegray leaf spot was 100%, while the concentration was 220 μg·mL-1, the relative inhibitory rate of grape leaf lesion was 94.4%.【Conclusion】The strain BH21 has the genes for synthesizing antimicrobial lipopeptide such as surfactins, iturins and fengycins, and the lipopeptide extracted from this strain has strong antagonism to, so it has potential application in the biological control of.
;; antifungal activity; lipopeptide; grapegray mold
2017-07-31;
2017-10-06
國家重點研發計劃(2016YFC0501303)、河北省應用基礎研究計劃重點基礎研究項目(15962904D)
魏新燕,E-mail:bdweixinyan@163.com。
黃亞麗,Tel:0311-83014618;E-mail:huangyali2291@163.com。通信作者杜克久,Tel:0312-7528491,E-mail:dukejiu@126.com
