持續低溫對水稻芽期出苗狀況的影響
2018-03-26 10:16:18王海媛段里成章起明楊愛萍劉文英姚俊萌金國花郭瑞鴿
江蘇農業科學 2018年4期
關鍵詞:水稻
張 坤, 王海媛, 段里成,, 章起明, 楊愛萍, 劉文英, 蔡 哲, 姚俊萌, 金國花, 郭瑞鴿
(1. 江西省農業氣象中心,江西南昌 330045; 2.江西農業大學,江西南昌 330045)
溫度是水稻生長的主要限制因子之一,不同生育時期遭遇低溫脅迫后響應各不相同,芽期低溫易導致出芽時間延長或爛種爛秧;苗期低溫易導致秧苗失綠、發僵、分蘗減少、枯萎甚至死亡;孕穗期低溫易導致花粉發育不良;揚花期低溫易導致無法正常授粉[1-2]。
江西省為雙季稻區,由于季節緊張,為銜接好早晚稻播種及收獲,對農時要求較為嚴格[3]。加之早稻芽期與苗期易遭受低溫冷害,晚稻生長孕穗及揚花期易受“寒露風”影響,因此選擇適宜播種期就更為重要。
近年來,因農村勞動力匱乏,省工省時、操作便捷的直播種植方式在江西省得以迅速發展。江西省早稻直播普適期一般要求日平均氣溫穩定通過12 ℃,普遍為4月上旬,但由于早稻直播會延期收獲,對晚稻品種選擇、移栽方式以及安全齊穗有一定不利影響[4]。為保證晚稻產量,留足充裕時間,鄱陽湖地區(贛北)早稻直播普遍提前,個別年份甚至提前至3月中旬,導致直播水稻芽期遭受低溫冷害的風險較大。因此,探究不同低溫條件對水稻出苗狀況的影響,選擇適宜播種溫度及播種期對規避芽期低溫冷害,確保早稻早播早收,為雙季晚稻栽插提供充足時間具有重要現實意義。
同時,有關苗期低溫冷害研究[2,5-10]表明,低溫處理顯著降低秧苗株高、葉面積、莖葉干質量,并且株高、葉長以及葉片卷枯程度、失綠程度可以作為鑒定水稻苗期抗寒能力的指標;但此時水稻均已播種10 d以上(1葉1心至4葉1心期),已具有獨立的根系,并較耐水淹,采取晝灌夜排方式可有效降低低溫冷害風險。而播種后芽期其根系尚未形成,水淹致死風險較大,遭遇低溫冷害難以采取有效措施加以應對。有關低溫冷害對早稻芽期的影響研究鮮有報道,因此探究低溫脅迫對芽期成苗率和幼苗生長的影響,明確其耐低溫特性及其低溫傷害閾值,可為合理利用光熱資源和降低低溫冷害風險的實際種植管理模式提供理論依據和技術保障。
1 材料與方法
1.1 試驗品種、設備、時間和地點
試驗品種為中嘉早17(常規超級早稻,當地農業局提供)。實驗設備為智能人工氣候箱HP1500GS(簡稱為氣候箱A)、智能人工氣候箱HP1000G-E(簡稱為氣候箱B)、培養箱。試驗于2016年7月開始在江西省防災減災科技園農業氣象實驗室進行。
1.2 試驗方法
試驗設9個溫度梯度(4、6、8、10、12、14、16、18、25 ℃,其中25 ℃為對照),5個處理時間(1、2、3、4、5 d),2種催芽程度(破胸、芽谷)。氣候箱A用于對照以及低溫處理后恢復生長,設置溫度25 ℃,光照強度3級,相對濕度80%,模擬白天光照時長9 h,黑暗時長15 h。氣候箱B用于低溫處理,除溫度外,其余設置與A一致。
1.2.1 破胸期處理 稱取經曬種、風選后的水稻種子,間歇浸種24 h(前12 h用25%咪鮮胺溶液浸種消毒),后置于培養箱中 35 ℃ 催芽,達到破胸(種子露白)后,選取催芽程度一致的180粒種子均勻播于規格為21 cm×15 cm×9 cm、裝有7 cm厚土層的塑料盒中(塑料盒提前24 h放入氣候箱,保持土溫與氣溫一致),每盒60粒,共30盒;其中15盒為對照組,其余為低溫組(以4 ℃為例)。低溫組在氣候箱B中處理1~5 d后,各移出3盒(1盒用于成苗率調查,另2盒用于取樣)于氣候箱A中,對照組相對應處理繼續在氣候箱A中生長。待各處理恢復生長7 d后,從氣候箱A中取出,然后進行成苗率及秧苗素質調查。其他低溫處理與4 ℃處理方法一致。
1.2.2 芽谷期處理 除在培養箱中催芽至芽谷半長(芽長約為谷粒長度一半,根長與谷粒等長)外,其余處理與“1.2.1”節一致。
1.3 測定指標及方法
1.3.1 成苗率調查 成苗率=經低溫處理移至25 ℃氣候箱恢復7 d后仍存活且苗高≥2 cm的幼苗數/60×100%。
1.3.2 秧苗素質調查 常溫恢復生長7 d后,隨機選取10株長勢一致的幼苗,3次重復,測定其株高、第一片完全葉葉長(簡稱第一葉長),然后將根和莖葉分離,放入烘箱105 ℃殺青30 min,80 ℃烘干至恒質量,稱量莖葉干質量。
1.4 數據處理和分析
采用Excel 2010和SPSS 21.0進行數據統計、制圖和方差分析。
2 結果與分析
2.1 不同低溫處理對成苗率的影響
對不同低溫處理下成苗率進行多因素方差分析,結果表明各處理之間以及各處理兩兩互作間均呈極顯著差異(P<0.01),說明不同溫度、不同持續時間和不同催芽程度均對成苗率有極顯著影響,而且不同處理之間的相互作用可進一步降低芽期成苗率。
從圖1可以看出,隨著氣溫降低或持續時間增加,成苗率均呈下降趨勢;相同溫度及持續時間,芽谷成苗率大都較破胸成苗率低。當氣溫≥12 ℃時,各處理成苗率隨氣溫降低以及持續時間增加逐漸下降,但彼此之間差異大都不顯著,平均成苗率大多在90%以上,平均成苗率降幅在0~5.2%之間,變化不明顯。當氣溫為10 ℃時,平均成苗率仍較高(84.2%~87.4%),平均成苗率降幅在8.25%~10.9%之間,其中破胸的成苗率均在82%以上,芽谷在持續低溫處理4~5 d后,成苗率較破胸成苗率下降4.66~8.00百分點,成苗率相對較高。當氣溫為8 ℃時,成苗率下降比較明顯,尤其是低溫處理3 d以上時,成苗率降至71%以下,其中平均成苗率降幅在15.6%~18.5%之間。當氣溫≤6 ℃時,隨處理時間增加,成苗率降幅更大,平均成苗率降幅在20.00%~46.40%之間,至處理5 d,4 ℃ 下芽谷幼苗全部死亡。
以上分析表明不同催芽程度中破胸較芽谷更耐低溫;氣溫穩定通過10 ℃時,仍可保持較高的成苗率,為保證直播稻適宜的基本苗數,其播種量較穩定通過12 ℃(假定12 ℃下的成苗數為適宜基本苗數,根據10 ℃和12 ℃下的最高和最低成苗率進行換算)需增加3.4%~6.4%。

2.2 不同低溫處理對莖葉干質量的影響
多因素方差分析表明,不同催芽程度對單株莖葉干質量無明顯影響(P>0.05),不同溫度及持續時間對單株莖葉干質量有極顯著影響(P<0.01)。隨著溫度降低,同一溫度不同持續時間處理平均單株莖葉干質量逐漸下降,各溫度梯度之間呈極顯著差異。
從圖2可以看出,氣溫在18~25 ℃時,隨著持續時間延長,不同催芽程度單株莖葉干質量逐漸增加;10~16 ℃時,隨著持續時間延長,不同催芽程度單株莖葉干質量呈先增后減規律,大都在3 d或4 d時達到最大值,說明持續時間超過3 d或4 d以后,地上部分干物質消耗大于積累,使得干物質總量逐漸減少;4~8 ℃時,隨著持續時間延長,不同催芽程度單株莖葉干質量逐漸減少,溫度越低,減少幅度越大,且芽谷處理下降幅度大于破胸處理。
在氣溫≤10 ℃且持續時間≥4 d或氣溫≤8 ℃且持續時間≥3 d或氣溫≤4 ℃且持續時間≥2 d,破胸處理地上部分干物質積累量大于芽谷處理,氣溫越低,差異越明顯。可能原因是隨著溫度降低和持續時間延長,有效抑制了淀粉酶活性及表達時間[11],貯藏在胚乳中的能量不能及時有效輸送至生長中心,使得在相同時間內地上部分干物質積累較少;而破胸處理的穎殼開裂程度較芽谷處理的小,且葉鞘還未長出,受低溫的影響相對較小,因此破胸程度干質量積累較芽谷強,較芽谷更能耐受低溫。
2.3 不同低溫處理對第一葉長的影響
不同催芽程度第一葉長均隨處理時間增加或溫度降低而減小,呈極顯著差異,且芽谷處理的第一葉片均較破胸短(表1),表明不同低溫及處理時間對第一葉長的生長有明顯抑制作用,對芽谷處理的抑制作用更強。
相同溫度、不同持續時間,隨持續時間增加(25 ℃破胸處理除外),第一葉長逐漸縮短且日均縮短幅度隨溫度降低逐漸增大。當氣溫≥14 ℃時,第一葉長較對照縮短幅度較小,平均降幅在6.21%~18.28%之間;氣溫為12 ℃時,平均降幅為21.01%~22.89%之間;氣溫為10 ℃時,平均降幅為23.68%~32.89%之間,氣溫≤8 ℃時,第一葉長降幅在33.32%~72.73%之間。
相同持續時間、不同溫度,隨氣溫降低,第一葉長逐漸變小;低溫處理時間≥3 d時,各處理溫度均與對照(25 ℃)存在顯著或極顯著差異;表明持續時間越長對第一葉長的抑制作用越大。
2.4 不同低溫處理對株高的影響
從圖3可以看出,在16~25 ℃下,隨著處理時間增加,不同催芽程度的株高逐漸增加; 10~14 ℃時, 隨著處理時間增加,株高呈先增后減規律,基本上在4 d達到最大值;4~8 ℃ 時,隨著處理時間增加,株高逐漸降低,表明在長期低溫條件下,不利于水稻幼苗莖葉生長。


表1 不同低溫處理對第一葉長的影響

2.5 不同低溫處理下成苗率與幼苗形態特征相關分析
相關分析表明,不同低溫處理下成苗率與單株莖葉干質量、第一葉長和株高均存在極顯著正相關(表2),說明成苗率與幼苗生長狀況密切相關,單株莖葉干質量越大、第一葉長越長、株高越高,則越有利于成苗率提升。綜上所述,為確保適宜基本苗,有較好的秧苗素質,并盡早播種,充分利用光熱資源(表1、圖1、圖2、圖3),實際生產中可在氣溫穩定通過 10 ℃ 時進行直播,單位面積播種量較12 ℃增加3.4%~6.4%。
3 小結與討論
3.1 不同低溫及持續時間對水稻芽期成苗率的影響
早稻直播后遭遇低溫冷害的風險較大,待溫度回升后,部分出苗的種子會出現爛種、爛芽情況,因此成苗率成為衡量直播稻播種質量好壞和苗期耐冷性強弱的重要標志[6,8]。孫世臣等認為,隨溫度降低,水稻種子發芽勢、發芽率逐漸減低,且呈極顯著差異;10 ℃的發芽勢、發芽率和發芽系數作為評價寒地水稻芽期耐寒性的主要指標較合理[12]。潘孝武等研究表明,隨著處理溫度下降和處理時間延長,芽期死苗率呈上升趨勢;8 ℃下低溫處理7 d作為秈稻早稻芽期耐寒性鑒定方式比較合適[13]。本研究表明,不同溫度、持續時間以及不同催芽程度均對水稻成苗率有極顯著影響,隨著溫度降低及其持續時間增加,不同催芽程度的成苗率均呈下降趨勢,芽谷處理的耐寒性較破胸處理的差。在氣溫≥10 ℃時,不同催芽程度均有較高的成苗率;當氣溫低至8 ℃時,平均成苗率降至80%以下,與前人研究結果比較吻合。由于中嘉早17是冷敏感品種[14],耐冷等級為最低級,因此依據成苗率,將10 ℃低溫處理5 d及以內的作為雙季早稻最低播種溫度是切實可行的。
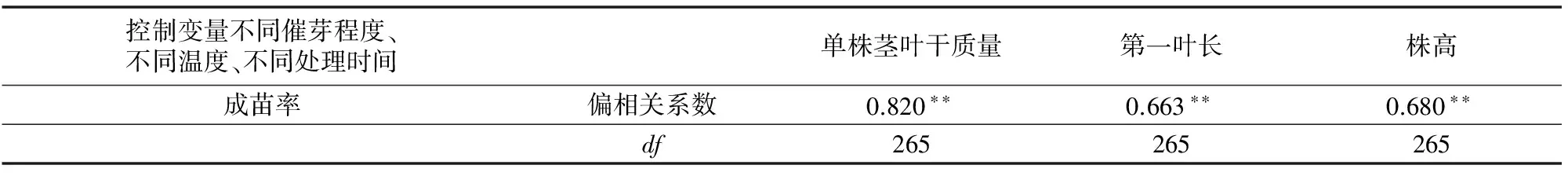
表2 不同低溫處理下成苗率與幼苗生長指標相關分析
注:“**”表示差異極顯著,“*”表示差異顯著。
3.2 不同低溫及持續時間對幼苗生長狀況的影響及原因分析
關于低溫處理后對幼苗生長狀況的影響,周新橋等和潘圣剛等認為,低溫處理會抑制水稻秧苗期生長發育,使得株高、莖葉干質量降低,明顯抑制葉片生長速度[5,15];同時葉齡、株高和倒一葉長相對值與苗期耐冷性存在極顯著正相關[2,8]。本研究認為,低溫處理會抑制莖葉干質量增加、株高和第一葉伸長;隨著溫度降低和處理時間增加,對芽谷抑制作用更強;成苗率與單株莖葉干質量、株高和第一葉長成極顯著正相關,與前人研究結果較一致。在氣溫≥10 ℃時,干物質積累、第一葉長以及株高下降幅度仍較小,在氣溫降至8 ℃時,秧苗素質出現明顯下降,不利于早稻早生快發以及后期產量提高。
造成上述現象可能與芽期營養物質供應有關。淀粉是水稻萌發的能量來源,淀粉水解需要通過淀粉酶作用才能完成。水稻干種子中僅存少量α-淀粉酶[16],可保證萌發初期物質和能量代謝的快速啟動,后期所需淀粉酶需要在盾片或糊粉層細胞重新合成,而低溫及處理時間累加可抑制淀粉酶活性及表達時間[11]。溫度越低,持續時間越長,對淀粉酶抑制作用越強,單位時間內水解淀粉就少,用于供應生長中心的能量和物質相對較少。在氣溫≥12 ℃時,對淀粉酶活性及表達沒有明顯抑制,供應的營養物質可滿足正常生長發育需要,當氣溫逐步降低及持續時間增加時,淀粉酶活性開始下降,甚至會進一步延遲酶的合成,因此低溫處理較對照的干物質積累量、苗高顯著降低。
水稻解剖觀察表明,萌發時胚中已分化形成胚芽鞘、不完全葉、第一葉和第二葉原基,在稻谷發芽出葉過程中,外部每長出1片葉,內部又分化出1個新的葉原基[17]。由于在低溫處理后7 d恢復期內,第一葉已分化形成,但受前期低溫影響,淀粉酶活性較低,不利于能量供應和化合物合成,因而對處生長中心的第一葉生長發育有很大的影響,明顯抑制了葉片生長,使得第一片葉葉長較短。
3.3 持續低溫導致不同催芽程度差異原因分析
除低溫可抑制淀粉酶活性外,谷粒開裂程度也會進一步降低胚活力,尤其是降低淀粉酶活性[18],由于芽谷的裂穎程度大于破胸,低溫狀況下淀粉酶活性下降更明顯,因此芽谷的低溫耐受性弱于破胸,成苗率、干物質積累在較低溫度下都弱于破胸處理;再加之芽谷葉鞘完全裸露在外,受到低溫影響更大,導致第一葉葉長較破胸處理更短。
4 結論
低溫及其持續時間會顯著降低水稻芽期單株莖葉干質量、株高、第一葉長和成苗率;且成苗率與芽期單株莖葉干質量、株高和第一葉長呈極顯著正相關,其形態指標亦可作為水稻芽期耐冷性鑒定指標。破胸較芽谷表現出更強的耐低溫性。綜合考慮成苗率、秧苗素質、充分利用光熱資源以及為雙搶提供充裕時間等因素,實際生產上可在氣溫穩定通過10 ℃時以破胸形態進行雙季稻區早稻直播,播種量較12 ℃增加3.4%~6.4%。
[1]聶元元,蔡耀輝,顏滿蓮,等. 水稻低溫冷害分析研究進展[J]. 江西農業學報,2011,23(3):63-66.
[2]傅泰露. 水稻不同生育期的耐冷性及鑒定指標的篩選[D]. 成都:四川農業大學,2008.
[3]朱德峰,陳惠哲,徐一成,等. 我國雙季稻生產機械化制約因子與發展對策[J]. 中國稻米,2013,19(4):1-4.
[4]潘曉華,李木英,曾勇軍,等. 江西雙季稻主要種植方式及其配套栽培對策[J]. 江西農業大學學報,2013,35(1):1-6.
[5]周新橋,陳達剛,李麗君,等. 秧苗期低溫脅迫對華南主推雙季稻生長的影響[J]. 西南農業學報,2013,26(3):936-941.
[6]王 蘭,龍云銘,田 華,等. 水稻苗期抗寒種質的篩選與鑒定[J]. 核農學報,2011,25(2):208-213.
[7]肖輝海,郝小花,吳小輝,等. 不同早秈稻品種幼苗期的耐冷生理鑒定[J]. 湖南文理學院學報(自然科學版),2012,24(1):46-50.
[8]傅泰露,馬 均,李 敏,等. 雜交水稻苗期耐冷性綜合評價及其鑒定指標的篩選[J]. 西南農業學報,2009,22(3):608-614.
[9]李 霞,戴傳超,程 睿,等. 不同生育期水稻耐冷性的鑒定及耐寒性差異的生理機制[J]. 作物學報,2006,32(1):76-83.
[10]Shimono H,Hasegawa T,Iwama K.Response of growth and grain yield in paddyrice to cool water at different growth stages[J]. Field Crop Research,2002,73(2/3):67-79.
[11]江 玲,候名語,劉世家,等. 水稻種子低溫萌發生理機制的初步研究[J]. 中國農業科學,2005,38(3):480-485.
[12]孫世臣,鄒德堂,劉化龍,等. 寒地水稻種質資源萌發期耐寒性鑒定評價[J]. 東北農業大學學報,2007,38(2):145-148.
[13]潘孝武,李小湘,黎用朝,等. 湖南早秈稻品種芽期和苗期耐冷性初步研究[J]. 雜交水稻,2015,30(5):59-64.
[14]趙 楊,魏穎娟,鄒應斌. 低溫脅迫下早稻幼苗葉片和根系的生理指標變化及其品種間差異[J]. 核農學報,2015,29(4):792-798.
[15]潘圣剛,孫杜鵬,雷 舜,等. 低溫處理對精量穴直播水稻幼苗生長特性的影響[J]. 華北農學報,2013,28(4):139-144.
[16]陸定志,楊煌峰,施天生,等. 雜交水稻干種子內存在α-淀粉酶[J]. 植物生理學報,1987,13(4):418-421.
[17]凌啟鴻,張洪程,蘇祖芳,等. 稻作新理論——水稻葉齡模式[M]. 北京:科學出版社,1994:27-30.
[18]劉洪偉,李小林,任萬軍. 裂穎與去殼對水稻種子萌發生理的初步研究[J]. 種子,2009,28(6):81-85.
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
