鱘源遲鈍愛德華氏菌的分離鑒定與藥敏特性研究
2018-03-27 01:11:59李小義
水產科學 2018年2期
孔 杰,李小義,周 洲,關 梅,石 婧
(1.貴州省水產研究所,貴州 貴陽 550025;2.貴州省疾病預防控制中心,貴州 貴陽 550004)
鱘魚是一類古老的軟骨硬鱗魚類,素有“活化石”之稱,具有巨大的經濟和科研價值。鱘魚幾乎全身是寶,肉質鮮美,富含多種人體必需氨基酸;魚鰾含有高級膠質可配制上等漆料,還可入藥,而鱘魚卵做成的魚子醬更有“黑色黃金”的美名。強大的市強需求刺激著我國鱘魚養殖市場的發展與擴大,然而由于沒有規范的引種與養殖制度,鱘魚養殖環境不斷惡化,使鱘魚細菌性疾病發生率不斷增加。已有報道的主要致病菌包括:氣單胞菌(Aeromonas)屬革蘭氏陰性菌,如嗜水氣單胞菌(A.hydrophila)、維氏氣單胞菌(A.veronii)、豚鼠氣單胞菌(A.carvia)和類志賀鄰單胞菌(Plesimonasshigelloides)[1-3];革蘭氏陽性菌鏈球菌(Streptococcus),如停乳鏈球菌(S.dysgalactiae)、海豚鏈球菌(S.iniae);此外還有魯氏耶爾森氏菌(Yersiniaruckeri)、弗氏檸檬酸桿菌(Citrobacterfreundii)[4-7]等。
遲鈍愛德華氏菌(Edwardsiellatarda),屬腸桿菌科愛德華氏菌屬,本屬細菌可分為3個種,即遲鈍愛德華氏菌、鲇魚愛德華氏菌(E.ictaluri)和保科愛德華氏菌(E.hoshinae)。遲鈍愛德華氏菌和鲇魚愛德華氏菌已被大量研究證實為重要的魚類病原微生物,而保科愛德華氏菌不能引起魚類疾病。遲鈍愛德華氏菌最初于1962年被日本學者Hoshina在鰻鱺(Anguillajapancia)中發現[8],隨后又分別在人類的排泄物和爬行類中分離到,是一種人魚共患的條件致病菌,也是愛德華氏菌屬里唯一感染人的病原菌,能引起患者腹瀉、腸胃炎、敗血癥及黃疸等疾病。遲鈍愛德華氏菌是水產養殖業中危害極大的革蘭氏陰性菌之一,它能引起鰻鱺、褐牙鲆(Paralichthysolivaceus)、大菱鲆(Scophthalmusmaximus)、大鱗大麻哈魚(Oncorhynchustshawytscha)[9-12]等數十種水產動物疾病。
2013年7月,貴州省惠水縣鱘魚養殖場養殖的雜交鱘[施氏鱘(Acipenserschrenckii♀)×達氏鰉(Husodauricus♂)]發生大量死亡,病魚的主要癥狀為腹部出現血點,肛門紅腫,鰓絲變白,部分尾部出現白色潰爛。剖檢發現內臟器官無血色,肝臟與腎臟腫大,嚴重的腸壞死穿孔。筆者從具有明顯發病癥狀的雜交鱘中分離得到一株高致病性病原菌,經過生理生化鑒定、回歸感染、16S rDNA序列分析和系統發育分析對該菌進行了分類鑒定,確定該致病菌為遲鈍愛德華氏菌,并研究了其藥敏特性,以期為鱘魚防治遲鈍愛德華氏菌提供理論參考依據。
1 材料與方法
1.1 材料
2014年7月采集瀕死病魚于貴州省水產研究所惠水鱘魚繁育基地;回歸感染用的健康雜交鱘也來自該基地,健康雜交鱘魚放水族箱中暫養7 d,無異常后進行試驗。
1.2 主要試劑
細菌基礎固體培養基、水解酪蛋白瓊脂按常規方法自配,細菌基因組DNA提取試劑盒、taq DNA聚合酶、dNTP等購自TAKARA公司,藥敏試紙片、細菌生化微量鑒定管均購自杭州濱和微生物試劑有限公司。
1.3 方法
1.3.1 病原菌的分離純化
2014年7月,貴州省惠水鱘魚養殖基地部分養殖池內發生鱘魚規模性疾病,病死魚頭腹朝天,腹部腫脹并出現明顯的血點,選取具有典型癥狀瀕死鱘魚,用70%酒精對魚體表面進行消毒滅菌,無菌環境下取病魚腎組織用于細菌分離,用經過灼燒的接種環取少量器官組織劃線于細菌基礎培養基平板上,置28 ℃下培養24 h后挑取形態大小、顏色等菌落特征一致的優勢菌落進一步純化,純化后的菌株用含30%甘油的LB培養液于冰箱-80 ℃保存備用。
1.3.2 理化特性測定
選取純化分離菌株,測定溫度5、15、20、28、35、42 ℃下的生長情況;測定鹽度(即NaCl質量分數)為0~7%下的生長情況;測定pH值為3.0~10.0下的生長情況。應用細菌生化微量鑒定管進行細菌的常規生理生化測定。
1.3.3 16S rDNA基因序列的測定與分析
純化培養的菌株,按照DNA提取試劑盒說明書提取細菌基因組DNA。PCR擴增引物使用細菌16S rDNA通用引物(27f:5′-AGAGTTTGATC-CTGGCTCAG-3′,1492r:5′-GGTTACCTTGTTA-CGACTT-3′);PCR擴增產物由生工生物工程上海(股份)有限公司進行測序。用BLAST在線同源性查詢軟件查詢菌株的16S rDNA序列屬性。從GenBank數據庫獲得同源性為98%的8株菌的16S rDNA序列,并選擇鲇魚愛德華氏菌模式菌株、保科愛德華氏菌模式菌株共4株、與愛德華氏菌進化關系較近的阪崎腸桿菌(Enterobactursakazakii)的16S rDNA序列進行同源性序列比對分析,然后構建菌株系統發育進化樹。
1.3.4 回歸感染試驗
選取體長10~12 cm的健康雜交鱘,設試驗組、平行試驗組、對照組和平行對照組,每組10尾。將純培養的菌落制成菌懸液,用麥氏比濁法測定并調節菌密度為1×109cfu/mL,分別對試驗組與平行試驗組鱘魚進行胸鰭基部注射,每尾注射0.3 mL,對照組與平行對照組注射相同劑量的無菌生理鹽水。試驗期間,水溫控制在23~25 ℃,充氧,隔日換水,連續觀察11 d至無新增死亡,記錄魚的發病和死亡情況,并對瀕死魚及時剖檢并再次分離純化鑒定致病菌。
1.3.5 藥物敏感性測定
對分離并經鑒定的菌株采用瓊脂擴散紙片法進行常用抗菌類藥物的敏感性測定,以明確這些菌株的耐藥情況并為臨床防治的用藥提供參考。藥敏試驗結果判別按照美國臨床實驗室標準化協會相關標準[13]執行。
2 結 果
2.1 病原菌的分離純化
從病魚的腎中分離到菌落形態相同的優勢菌,28 ℃培養24 h的菌落在LB固體培養基上呈圓形光滑、半透明、邊緣整齊、灰白色菌落。革蘭氏陰性桿菌,將病原菌編號為K9。
2.2 生理生化鑒定
經測定,鹽度為0~30時菌株K9生長正常,40以上鹽度不生長;pH為5~8時生長正常,最適pH為7。其他生理生化特性見表1,菌株ET81126[14]、菌株HC010907-1[10]、菌株HN005[15]均為已鑒定的遲鈍愛德華氏菌,除了對蔗糖的利用與ET81126和HN005不同外,菌株K9的其他特性與遲鈍愛德華氏菌株表現一致。
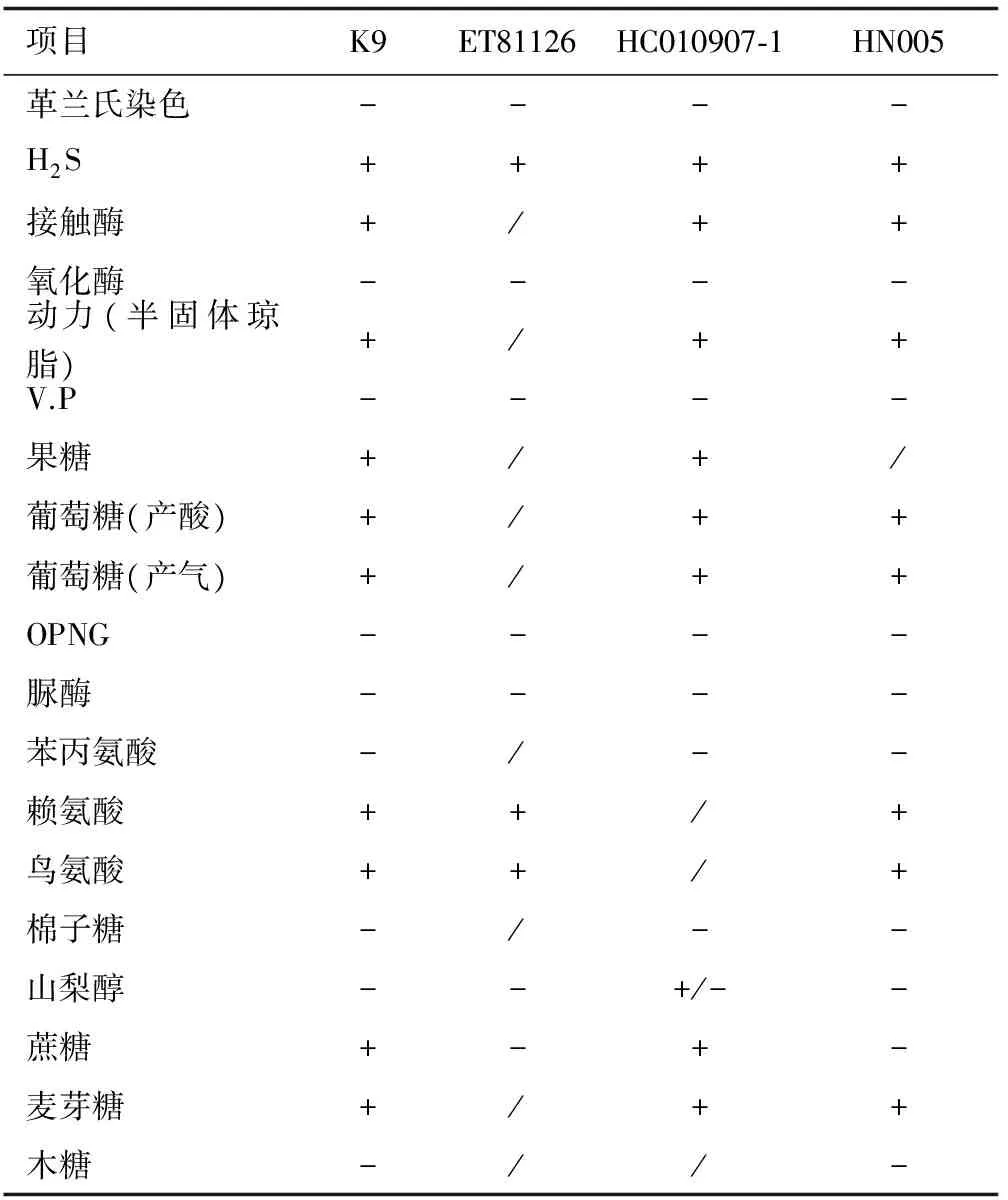
表1 菌株K9的生理生化特性
注:+表示陽性,-表示陰性,+/-表示介于+與-之間.
2.2 病原菌的16S rDNA鑒定
菌株K9經PCR擴增所獲得的16S rDNA基因序列長度為1468 bp,GeneBank 登錄號為:KY489780;在GeneBank 數據庫上進行同源性比對分析,結果顯示,菌株K9與遲鈍愛德華氏菌的同源性為98%。在EZTaxon上進行同源序列比對,結果顯示,K9菌株與愛德華氏菌ET833(T) (JRGQ01000134)、鲇魚愛德華氏菌ATCC_33202(T) (AFJI01000002)、保科愛德華氏菌NBRC105699T (BAUC01000062)、遲鈍愛德華氏菌NBRC_105688(T) (BANW01000030)的相似性分別為98.34%、97.99%、97.91%、97.71%。選擇以上序列以及腸桿菌科中的阪崎腸桿菌構建進化樹,結果見圖1,菌株K9與遲鈍愛德華氏菌聚為1支。細菌分類學上,通常將16S rRNA基因同源性大于97.5%的菌株定為同種菌株[16-17],因此綜合生化指標、16S rDNA基因序列分析以及進化樹的分析結果,可將菌株K9鑒定為遲鈍愛德華氏菌。

圖1 根據菌株K9 16S rDNA基因序列同源性構建的系統發育樹
2.3 回歸感染試驗結果
分離遲鈍愛德華氏菌K9通過接種感染,顯示對雜交鱘有強致病力。在11 d的感染中,試驗組感染24 h后開始發病死亡,1 d內死亡4尾,之后2 d又死亡3尾,至第11 d共死亡8尾。試驗過程中,死亡魚腹部兩側出現紅點,肛門紅腫,鰓絲變白,在試驗后期死亡的魚背部出現白色潰爛,腸部穿孔。病癥與自然發病鱘魚基本一致。平行試驗組11 d的觀察中,共死亡7尾,而對照組與平行對照組在感染試驗期內,沒有發病死亡病例。從感染死魚中再次分離菌株,經16S rDNA鑒定,與從自然發病鱘魚中所分離的菌株為同一株。
2.4 藥物敏感試驗結果
分離遲鈍愛德華氏菌K9對20種藥物敏感性研究結果表明,遲鈍愛德華氏菌K9對鏈霉素、阿米卡星、左氧氟沙星、阿齊霉素、新霉素、氯霉素(禁藥)等6種抗生素敏感,對頭孢噻肟、阿莫西林、頭孢唑啉、氨芐西林、紅霉素(禁藥)等5種抗生素等中度敏感,對青霉素G、妥布霉素、麥迪霉素等9種抗生素耐藥(表3)。
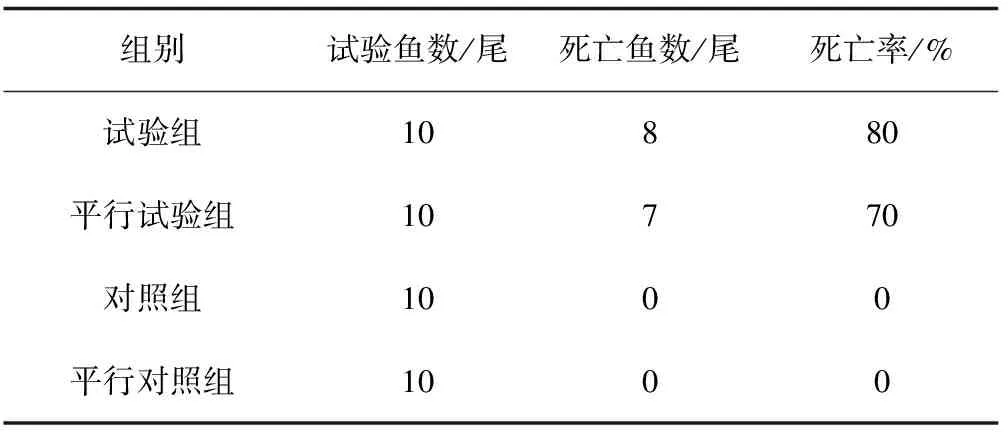
表2 遲鈍愛德華氏菌K9對鱘魚的致性病試驗
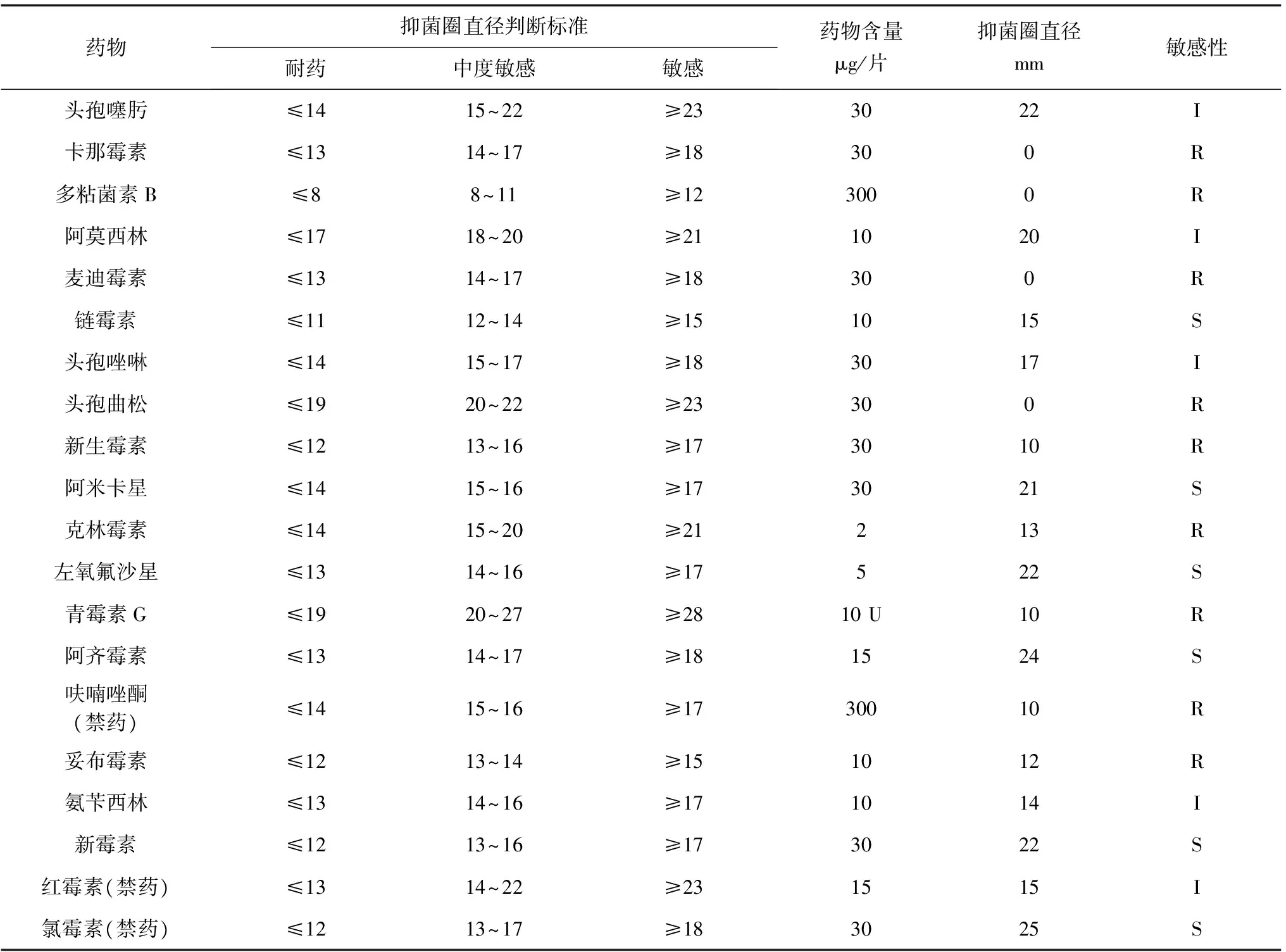
表3 分離遲鈍愛德華氏菌K9藥敏試驗結果
注:S,敏感;I,中度敏感; R,抗性.
3 討 論
遲鈍愛德華氏菌于1962年首次在鰻鱺中被發現,隨后在大口黑鱸(Micropterussalmoides)[18]、美洲紅點鮭(Salvelinusfontinalis)[19]、羅非魚(Oreochromis)[20]、褐牙鲆[10]、大菱鲆[21]、黃顙魚(Pelteobagrusfulvidraco)[15]、尖吻鱘(Acipenseroxyrinchus)[22]、斑馬魚(Daniorerio)[23]等數十種水產品種中均有報道。本研究從患病的雜交鱘腎臟組織中分離到1株革蘭氏陰性桿菌K9,人工感染試驗表明K9能使健康鱘魚致病死亡,表現出與自然感染下相似的臨床癥狀。遲鈍愛德華氏菌對斑馬魚與透明四帶無須鲃(Puntiustetrazona)[23]的人工感染試驗表明,當菌液密度達到106cfu/mL時,能分別引起90%和80%的死亡率;在遲鈍愛德華氏菌株ET81126在對歐洲鰻(A.anguilla)的致病性試驗中[14],當菌液密度為106cfu/mL時,歐洲鰻的死亡率達到了100%;尖吻鱘源遲鈍愛德華氏菌的死亡率為50%[22];本研究中,遲鈍愛德華氏菌K9的人工感染試驗結果顯示,在菌液密度為109cfu/mL時,引起鱘魚75%的死亡率,表明遲鈍愛德華氏菌對鱘魚的致病性較其他水生動物弱。
本研究的藥敏結果顯示,遲鈍愛德華氏菌K9只對鏈霉素、阿米卡星、左氧氟沙星、阿齊霉素、新霉素、氯霉素等6種藥物敏感,對青霉素G、妥布霉素、麥迪霉素、呋喃唑酮等9種抗生素表現出很強的耐藥性。與分離自斑馬魚中菌株Z1[23]相比,在Z1表現為敏感的藥物中,如青霉素G、妥布霉素、頭孢唑啉、慶大霉素、紅霉素、卡那霉素,K9表現均為中度敏感甚至耐受。與分離自牙鲆[10]中的遲鈍愛德華氏菌相比,K9對該菌株表現為敏感的頭孢曲松、妥布霉素、呋喃唑酮和耐藥的青霉素、克林霉素均表現耐受性,表明K9耐藥性較為強烈。葛慕湘等[24]對35株遲鈍愛德華氏菌的耐藥檢測顯示,大部分菌株耐抗菌類藥物達7~9種。本次試驗分離到的遲鈍愛德華氏菌耐受9種抗生素,與其研究結果相同,進一步表明遲鈍愛德華氏菌的耐藥性嚴重性。本研究中遲鈍愛德華氏菌表現為耐藥性的抗生素,并非意味著該菌株對這些抗生素完全耐受,從刁菁等[25]的研究中可以看出,雖然遲鈍愛德華氏菌在低抗生素含量下表現為耐受,但是隨著抗生素含量的增加細菌的生長受到抑制。因此,遲鈍愛德華氏菌K9表現為耐受的抗生素,也可能會在增加抗生素含量后表現為敏感。需注意的是,在本次藥敏試驗所用抗生素中,氯霉素、紅霉素和呋喃唑酮為國家禁用抗生素,僅供試驗研究用,嚴禁用于生產中。因此,防治遲鈍愛德華氏菌引起的疾病,可以考慮采用其他遲鈍愛德華氏菌K9敏感的抗生素,如阿米卡星、左氧氟沙星、阿齊霉素、新霉素。遲鈍愛德華氏菌感染魚體通常是由水溫升高所致[22],因而建議養殖中注意控制水溫,當疾病發生后,除給藥外,還應同時采取相應的降溫措施。
目前水產養殖中對細菌性疾病的防治仍以抗生素為主,而長期使用抗生素極易產生耐藥菌株,本研究中分離到具有較強耐藥性菌株K9,可能就是由于生產上長期使用抗生素導致。中草藥不易產生耐藥性、毒副作用小,成為研究抗菌藥物的熱點。楊移斌等[26]就曾對鱘源的3種致病菌進行過中草藥研究,結果表明,烏梅、石榴皮、地榆、杞子對防治鱘魚細菌性疾病效果良好。而不同來源的遲鈍愛德華氏菌也開展過許多中草藥抑菌研究,如五倍子與石榴皮對斑馬魚源的遲鈍愛德華氏菌抑菌作用明顯[23],最小抑菌質量濃度和最小殺菌質量濃度均≤6.25 mg/mL;五倍子、訶子、黃芩、秦皮和紅藤對牙鲆源的遲鈍愛德華氏菌抑菌作用明顯[27],最小抑菌質量濃度<12.5 mg/mL,最小殺菌質量濃度<50 mg/mL;五倍子+黃芩、五倍子+大黃、黃連+黃芩和大黃+黃芩對石斑魚源的遲鈍愛德華氏模式菌具有明顯的強力殺菌作用[28],最小抑菌質量濃度和最小殺菌質量濃度均≤3.125 mg/mL;陳言峰等[29]研究表明,20種中草藥中五倍子的殺菌能力極強,最小抑菌質量濃度為1.5626 mg/mL;從以上數據可以看出,五倍子對防治遲鈍愛德華氏菌具有明顯的作用,在防治鱘源遲鈍愛德華氏菌中不僅可以考慮本文中給出的抗生素藥物,還可以考慮中草藥的防治方法。
[1] 楊治國.鱘魚嗜水氣單胞菌的分離鑒定[J].淡水漁業,2001,31(5):40-41.
[2] 楊移斌,夏永濤,邱軍強,等.鱘源維氏氣單胞菌分離鑒定及藥敏特性研究[J].四川動物,2014,33(3):414-418.
[3] 曹海鵬,楊先樂,高鵬,等.鱘細菌性敗血綜合征致病菌的初步研究[J].淡水漁業,2007,37(2):53-56.
[4] 潘厚軍,劉曉勇,常藕琴,等.西伯利亞鱘停乳鏈球菌的分離、鑒定與致病性[J].中國水產科學,2009,16(6):891-903.
[5] 鄧夢玲,耿毅,劉丹,等.西伯利亞鱘海豚鏈球菌的分離鑒定及毒力基因檢測[J].水產學報,2015,39(1):127-135.
[6] 楊移斌,夏永濤,鄭衛東,等.鱘源魯氏耶爾森氏菌的分離鑒定[J].水生生物學報,2013,37(2):393-398.
[7] 楊移斌,夏永濤,趙蕾,等.鱘源弗氏檸檬酸桿菌分離鑒定及藥敏特性研究[J].水生生物學報,2013,37(4):766-771.
[8] Hoshina T.On a new bacterium,Paracolobactrumanguillimortifn.sp.[J].Bull Jpn Soc Sci Fish,1962,28(2):162-164.
[9] Amandi A,Hiu S F,Rohovec J S,et al.Isolation and characterization ofEdwardsiellatardafrom fall chinook salmon (Oncorhynchustshawytscha)[J].Applied & Environmental Microbiology,1982,43(6):1380-1384.
[10] 張曉君,戰文斌,陳翠珍,等.牙鲆遲鈍愛德華氏菌感染癥及其病原的研究[J].水生生物學報,2005,29(1):31-26.
[11] 王印庚,秦蕾,張正,等.養殖大菱鲆的愛德華氏菌病[J].水產學報,2007,31(4):487-495.
[12] 陳昌福,吳志新,高漢嬌.日本鰻鱺愛德華氏菌病原菌的分離[J].華中農業大學學報:自然科學版,1998,17(4):382-388.
[13] Clainical and Laboratory Standards Institute.CLSI document M100-S21,Performance Standards for Antimicrobial Susceptibility Testings[S].950 West Valley Road,Suite 1400,Wayne,Pennsylvania 19087,USA: Clinical and Laboratory Standards Institute,2011.
[14] 陳強,龔暉,楊金先.歐洲鰻遲鈍愛德華氏菌的分離鑒定[J].中國人獸共患病學報,2011,27(1):7-10.
[15] 鄧先余,羅文,譚樹華,等.黃顙魚(Pelteobagrusfulvidraco)“紅頭病”病原菌遲鈍愛德華氏菌(Edwardsiellatarda)的分離及鑒定[J].海洋與湖沼,2008,39(5):511-516.
[16] 魯辛辛,黃艷飛,田曉波.細菌rDNA分類鑒定的方法學研究[J].中華檢驗醫學雜志,2006,29(5):435-437.
[17] 蹇文嬰,東秀珠.定向進化同源基因在細菌系統進化研究中的應用[J].微生物學通報,2000,27(5):377-381.
[18] Francis-Floyd R,Reed P,Bolon B.An epizootic ofEdwardsiellatardain largemouth bass (Micropterussalmoides)[J].Journal of Wildlife Diseases,1993,29(2):334-336.
[19] Uhland F C,Helie P,Higgins R.Infections ofEdwardsiellatardaamong brook trout in Quebec[J].Journal of Aquatic Animal Health,2000,12(1):74-77.
[20] Clavijo A M,Conroy G,Conroy D A.First report ofEdwardsiellatardafrom tilapias in Venezuela[J].Bull Eur Assoc Fish Pathol,2002,22(4):280-282.
[21] 秦蕾.遲鈍愛德華氏菌感染大菱鲆的病理學研究[J].中國水產科學,2009,16(3):411-419.
[22] Francis-Floyd R,Dvm M S.Disease History of Cultured Sturgeon in Florida,1990-1999 [G]//Metcalf K,Zajicek P.Proceedings of the Florida Sturgeon Culture Risk Assessment Workshop,Florida: Florida Department of Agriculture and Comsumer Services,2000:33-37.
[23] 劉春,李凱彬,王慶,等.斑馬魚遲緩愛德華氏菌的鑒定、致病性及藥物敏感性[J].華中農業大學學報:自然科學版,2013,32(3):105-111.
[24] 葛慕湘,靳曉敏,張艷英,等.遲鈍愛德華氏菌耐藥表型及4種耐藥基因檢測[J].水產科學,2015,34(5):300-304.
[25] 刁菁,楊秀生,葉海斌,等.一種水產遲鈍愛德華氏菌快速藥敏檢測方法的研究[J].水產科學,2014,33(1):29-34.
[26] 楊移斌,曹海鵬,夏永濤,等.41種中草藥對3種鱘源病原菌的體外抑菌效果[J].淡水漁業,2013,43(4):80-84.
[27] 朱壯春,史相國,張淑杰,等.中草藥對牙鲆病原遲鈍愛德華氏菌的體外抑制作用研究[J].水產科學,2007,26(5):278-281.
[28] 李瑞,白東清,郭永軍,等.9種中草藥及17組方對遲鈍愛德華氏菌的體外抑菌試驗[J].水利漁業,2008,28(4):114-116.
[29] 陳言峰,鄒記興.20種中草藥對遲緩愛德華氏菌的體外抑菌試驗[J].水生態學雜志,2011,32(5):110-112.
