溫度和鹽度對(duì)波紋巴非蛤胚胎、幼蟲(chóng)及稚貝發(fā)育的影響
2018-03-27 00:33:34宋志民唐賢明王國(guó)福
水產(chǎn)科學(xué) 2018年2期
司 和,宋志民,唐賢明,王國(guó)福
(海南省海洋與漁業(yè)科學(xué)院,海南 海口 570100)
波紋巴非蛤(Paphiaundulata)屬軟體動(dòng)物門、雙殼綱、簾蛤目、簾蛤科、巴非蛤?qū)伲饕植加谖覈?guó)及東南亞沿海,在澳大利亞及紅海沿岸也有分布[1]。棲息在低潮線下0.5~44.0 m的泥沙底質(zhì)或軟泥底質(zhì)中,營(yíng)埋棲生活,環(huán)境適應(yīng)性廣,繁殖力強(qiáng)、生長(zhǎng)速度快,是淺海增養(yǎng)殖的新品種。同時(shí)波紋巴非蛤含有小分子多肽、生物活性肽、糖胺聚糖等活性成分[2-7],其提取物具有較好的抗氧化性、降血脂等藥用及保健作用[8-15]。由于波紋巴非蛤的養(yǎng)殖苗種主要依靠自然采捕,致使野生苗種逐年減少,苗種短缺成為制約我國(guó)波紋巴非蛤養(yǎng)殖發(fā)展的瓶頸之一。本文探討了溫度及鹽度對(duì)波紋巴非蛤胚胎、幼蟲(chóng)及稚貝發(fā)育的影響,為波紋巴非蛤的苗種繁育提供參考。
1 材料與方法
1.1 材料
智能人工氣候箱(PRX-350B),顯微鏡(XSP6C),細(xì)胞計(jì)數(shù)板,50 mL燒杯,250 mL燒杯,培養(yǎng)皿,篩絹。
1.2 試驗(yàn)方法
試驗(yàn)親貝殼長(zhǎng)(47.7±2.2) mm,殼寬(26.3±1.3) mm,體質(zhì)量(13.4±2.3) g,2014—2016年連續(xù)引自廣西北海。親貝清洗后在室外催熟,主要投喂亞心形扁藻(Platymonassubcordiformis)為主,輔以牟氏角毛藻(Chaetocerosmuelleri)和微綠球藻(Nannochlropsisoculata)。人工催產(chǎn)時(shí)先陰干4 h,再流水刺激,之后清洗并收集受精卵進(jìn)行試驗(yàn)。
1.2.1 溫度對(duì)波紋巴非蛤發(fā)育的影響
通過(guò)光照培養(yǎng)箱調(diào)節(jié)不同的溫度來(lái)研究溫度對(duì)波紋巴非蛤胚胎、幼蟲(chóng)及稚貝發(fā)育的影響,設(shè)置溫度梯度為15、20、25、30、35 ℃,鹽度30,光照度1000 lx,晝夜光周期為12 h∶12 h。
1.2.1.1 溫度對(duì)波紋巴非蛤胚胎發(fā)育的影響
將收集的受精卵按100個(gè)/mL的密度放入50 mL小燒杯中,每個(gè)燒杯放入35 mL滅菌的海水,微波充氣,每個(gè)溫度梯度設(shè)置3個(gè)平行。12 h后計(jì)數(shù)統(tǒng)計(jì)發(fā)育成D形幼蟲(chóng)的孵化率及畸形率。
1.2.1.2 溫度對(duì)波紋巴非蛤幼蟲(chóng)發(fā)育的影響
用400目篩絹收集D形幼蟲(chóng),計(jì)數(shù)后按100個(gè)/mL的密度放入250 mL的燒杯中,每個(gè)燒杯放入200 mL滅菌的海水,下覆載玻片方便觀察,微波充氣,每個(gè)溫度梯度設(shè)置3個(gè)品行。每隔12 h觀察一次。
1.2.1.3 溫度對(duì)波紋巴非蛤稚貝生長(zhǎng)的影響
將發(fā)育的稚貝放入培養(yǎng)皿中進(jìn)行觀察,培養(yǎng)皿鋪設(shè)1~2 mm厚的細(xì)砂。設(shè)置密度設(shè)為5個(gè)/mL。每隔一周計(jì)數(shù)一次。設(shè)置3個(gè)平行。
1.2.2 鹽度對(duì)波紋巴非蛤發(fā)育的影響
通過(guò)光照培養(yǎng)箱調(diào)節(jié)鹽度來(lái)研究鹽度對(duì)波紋巴非蛤胚胎、幼蟲(chóng)及稚貝發(fā)育的影響,鹽度梯度為15、20、25、30、35,溫度30 ℃,光照度1000 lx,晝夜光周期為12 h∶12 h。試驗(yàn)方法同相應(yīng)的溫度試驗(yàn),測(cè)定孵化率、畸形率、變態(tài)率、附著率及存活率。
1.3 數(shù)據(jù)分析
試驗(yàn)數(shù)據(jù)利用Excel 2007及SPSS 20.0進(jìn)行數(shù)據(jù)處理和統(tǒng)計(jì)分析,采用單因素方差分析或T-test分析,設(shè)顯著水平為0.05。
2 結(jié)果與分析
2.1 溫度對(duì)波紋巴非蛤發(fā)育的影響
2.1.1 對(duì)胚胎發(fā)育的影響
隨著溫度的升高胚胎孵化率逐漸升高,在30 ℃時(shí)達(dá)到最大值,之后逐漸降低(圖1)。畸形率與孵化率呈負(fù)相關(guān),在低溫和高溫時(shí)均較高,在30 ℃時(shí)達(dá)到顯著性的最低值。
2.1.2 對(duì)幼蟲(chóng)發(fā)育的影響
幼蟲(chóng)存活率隨著溫度的升高先升后降,在30 ℃達(dá)顯著最大值,25 ℃次之,其他3組存活率都不高,三者均無(wú)顯著性差異,15 ℃存活率最低。附著率隨溫度的升高而先升后降,30 ℃附著率最高,20~25 ℃次之,兩者差異不顯著。30 ℃變態(tài)率最高,25 ℃次之,20 ℃變態(tài)率極低,三者均存在顯著性差異,35 ℃及15 ℃不適合幼蟲(chóng)的發(fā)育生長(zhǎng)(圖2)。
2.1.3 對(duì)稚貝生長(zhǎng)的影響
30 ℃稚貝生長(zhǎng)最快,4周后稚貝殼長(zhǎng)達(dá)(2782±489.5) μm(圖3)。25 ℃生長(zhǎng)次之,但在第3周及第4周時(shí)與30 ℃差異不顯著。35 ℃不適合稚貝生長(zhǎng),15 ℃生長(zhǎng)最慢。
2.2 鹽度對(duì)波紋巴非蛤發(fā)育的影響
2.2.1 對(duì)胚胎發(fā)育的影響
隨著鹽度的增高,孵化率逐漸提高,在30時(shí)最高,35雖略有下降,但與鹽度30時(shí)無(wú)顯著性差異(圖4)。畸形率隨鹽度的升高先降后升,鹽度30時(shí)最低。
2.2.2 對(duì)幼蟲(chóng)發(fā)育的影響
幼蟲(chóng)存活率、附著率及變態(tài)率均隨鹽度的升高先升后降(圖5)。在鹽度30時(shí)存活率、附著率及變態(tài)率都最高。鹽度35與30,幼蟲(chóng)附著率差異不顯著。鹽度15不適合幼蟲(chóng)生長(zhǎng)。
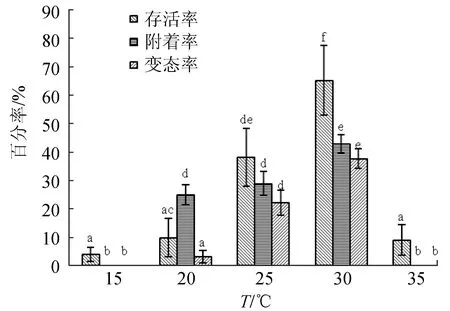
圖2 溫度對(duì)波紋巴非蛤幼蟲(chóng)發(fā)育的影響
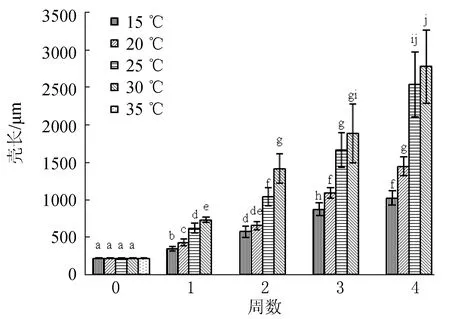
圖3 溫度對(duì)波紋巴非蛤稚貝生長(zhǎng)的影響
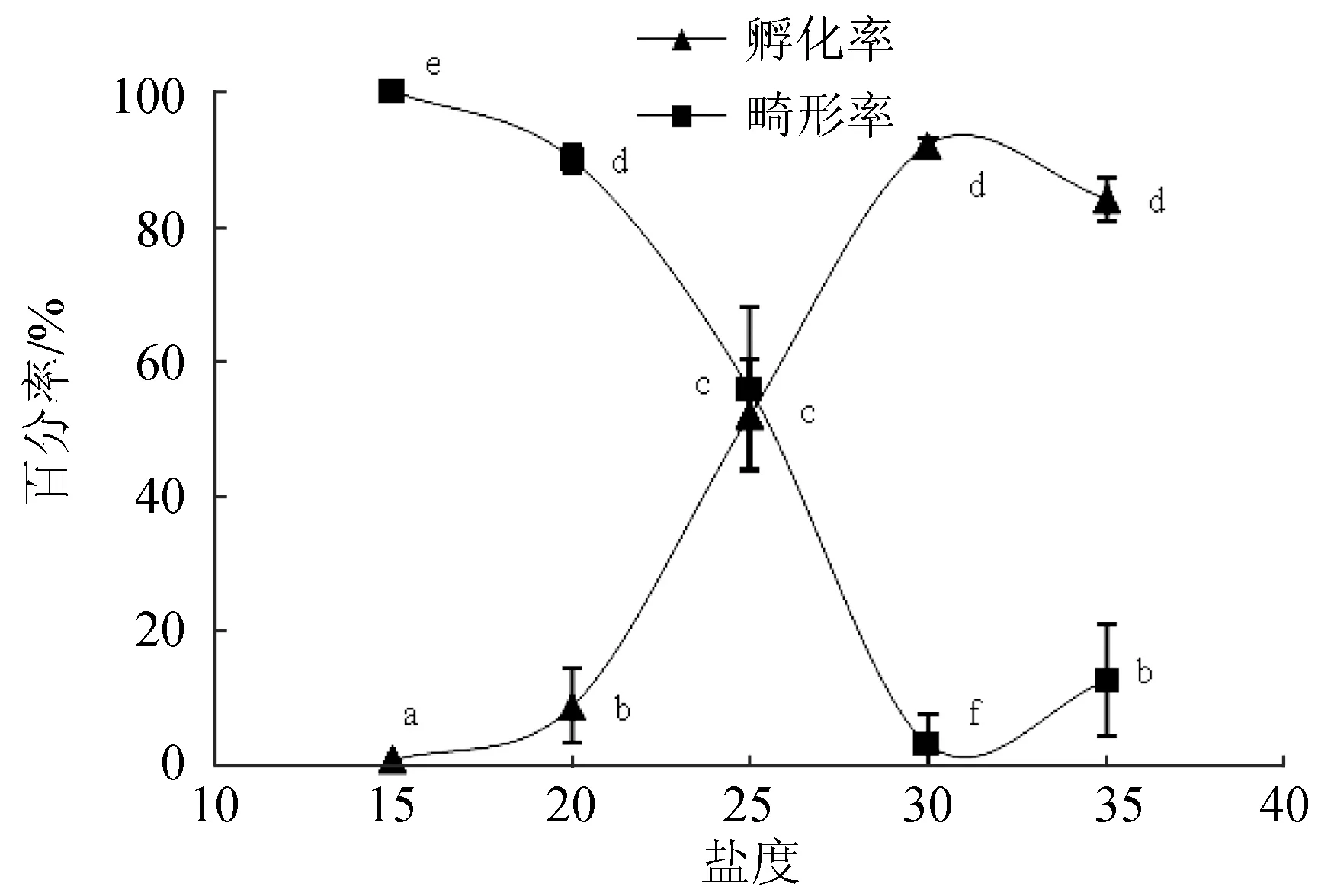
圖4 鹽度對(duì)波紋巴非蛤胚胎發(fā)育的影響

圖5 鹽度對(duì)波紋巴非蛤幼蟲(chóng)發(fā)育的影響
2.2.3 對(duì)稚貝生長(zhǎng)的影響
隨鹽度增高生長(zhǎng)速率逐漸加快,鹽度30生長(zhǎng)最快,但相比鹽度35差異不顯著(圖6)。鹽度小于20時(shí)波紋巴非蛤全部死亡。
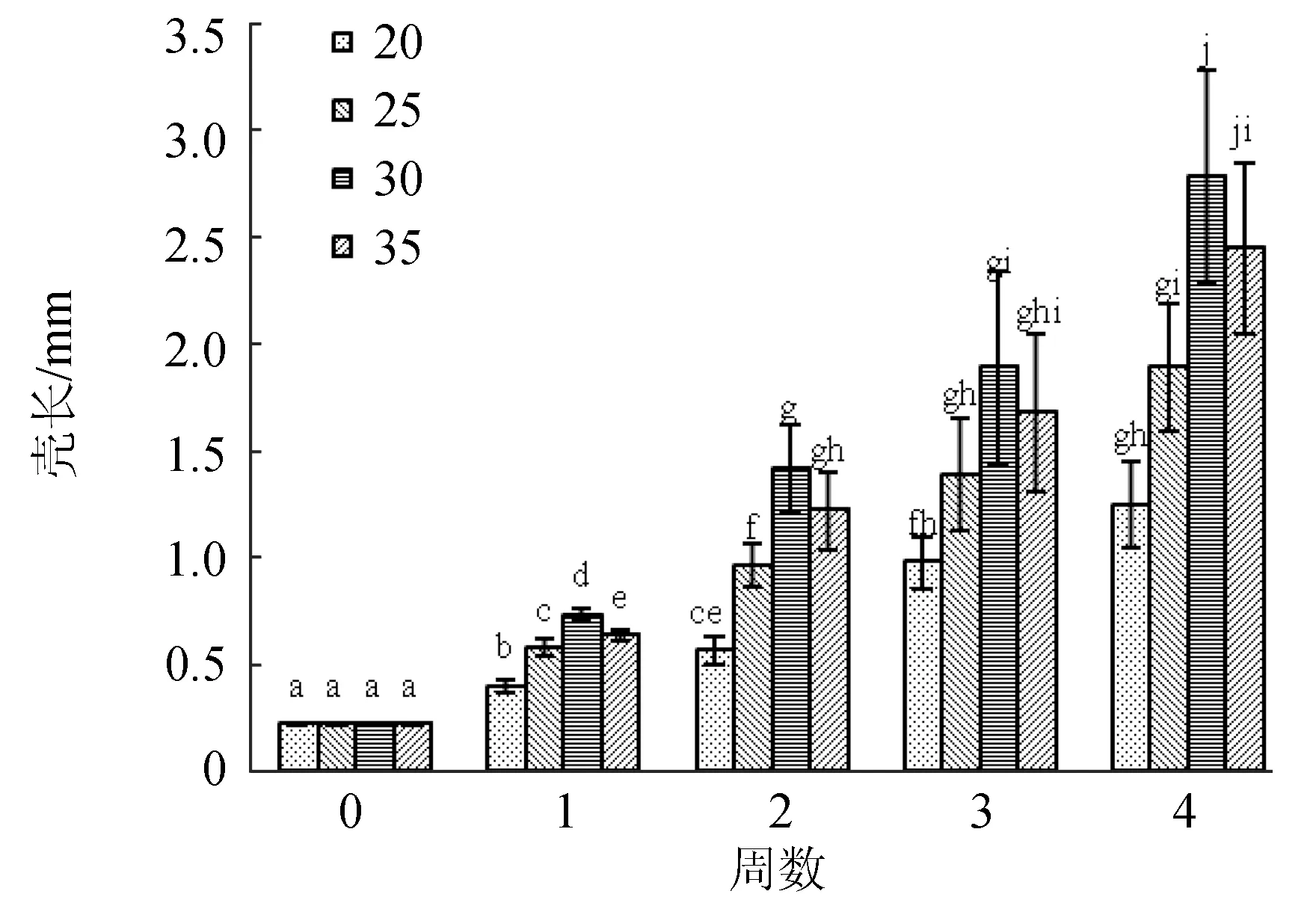
圖6 鹽度對(duì)波紋巴非蛤稚貝生長(zhǎng)的影響
3 討 論
20世紀(jì)80年代起,我國(guó)對(duì)波紋巴非蛤育苗及養(yǎng)殖技術(shù)陸續(xù)開(kāi)展了一系列研究[16-19]。吳洪流等[20-22]觀察了波紋巴非蛤的性腺組織組織學(xué),并對(duì)性腺發(fā)育進(jìn)行了分期,為育苗工作提供了理論依據(jù)及指導(dǎo)。陳何東[23]進(jìn)行了波紋巴非蛤育苗試驗(yàn),在28~29 ℃,海水相對(duì)密度1.021~1.022下育成了大量的波紋巴非蛤稚貝,在室外軟泥底池塘中進(jìn)行了稚貝培育。王萬(wàn)東等[24-26]在育苗過(guò)程中發(fā)現(xiàn),高溫天氣對(duì)育苗產(chǎn)生負(fù)面影響。徐小偉等[27]系統(tǒng)研究了幾種生態(tài)因子對(duì)波紋巴非蛤胚胎發(fā)育的影響,發(fā)現(xiàn)波紋巴非蛤胚胎發(fā)育的適宜溫度為27~30 ℃;適宜鹽度為27~33,與本研究的結(jié)果一致。徐小偉等[28]對(duì)波紋巴非蛤的胚胎發(fā)育進(jìn)行了掃描電鏡觀察,豐富了胚胎發(fā)育的形態(tài)學(xué)內(nèi)容。洪一川等[29]在室外條件下對(duì)波紋巴非蛤進(jìn)行了催產(chǎn)及幼蟲(chóng)培育,發(fā)現(xiàn)14.6~20 ℃時(shí)幼蟲(chóng)發(fā)育緩慢且無(wú)法完成變態(tài),23~29.6 ℃時(shí)則發(fā)育正常,這與本試驗(yàn)的結(jié)果相吻合。陳志等[30]通過(guò)響應(yīng)面法研究了溫度、鹽度和密度交互作用對(duì)波紋巴非蛤稚貝生長(zhǎng)的影響,結(jié)果表明,鹽度對(duì)稚貝的生長(zhǎng)影響最為顯著,其次是溫度。李俊輝等[31-32]通過(guò)研究溫度及鹽度對(duì)波紋巴非蛤耗氧率及排氨率的影響發(fā)現(xiàn),15~30 ℃,耗氧率和排氨率隨著溫度的升高而增大;在鹽度20~28時(shí),耗氧率和排氨率隨著鹽度的升高而升高,28~36時(shí),單位耗氧率與排氨率均隨鹽度的增大而降低。王冬梅等[33]發(fā)現(xiàn),在鹽度為24~32時(shí),波紋巴非蛤?yàn)V水率隨鹽度的升高而增大,鹽度為32~36時(shí)則隨鹽度的增大而減小。陳堅(jiān)等[1,30]的研究發(fā)現(xiàn),波紋巴非蛤成貝隨著鹽度的升高存活率先升后降,適宜鹽度為24~36。張躍平等[34]首次對(duì)波紋巴非蛤胚胎及浮游幼蟲(chóng)的形態(tài)發(fā)育和生長(zhǎng)特點(diǎn)進(jìn)行了報(bào)道,并與簾蛤科另外5種經(jīng)濟(jì)種類進(jìn)行了比較。邵艷卿等[35]利用組織學(xué)方法對(duì)斧文蛤(Meretrixlamarckii)性腺發(fā)育及生殖周期進(jìn)行了研究,在水溫29 ℃、鹽度28、pH 7.8~8.5的條件下由受精卵孵化成D形幼蟲(chóng),并經(jīng)培養(yǎng)最終發(fā)育變態(tài)為稚貝。
溫度和鹽度是決定育苗成敗的關(guān)鍵的重要因子。隨著波紋巴非蛤需求量的逐漸增加,大規(guī)模開(kāi)展育苗工作迫在眉睫。根據(jù)近年公布的研究成果,在室外開(kāi)展波紋巴非蛤育苗工作總是難以持續(xù)的獲得波紋巴非蛤成貝,這與南方各海區(qū)夏秋兩季的海水特點(diǎn)密切相關(guān)。夏季水溫合適,但雨水偏多,加之臺(tái)風(fēng)的影響,海水鹽度變化過(guò)大,各育苗中心受此影響較大。進(jìn)入秋冬季節(jié),臺(tái)風(fēng)雨水減少,水溫下降至偏低的階段,對(duì)波紋巴非蛤的育苗工作不利。尋找合適的溫度及鹽度是育苗成敗的關(guān)鍵之一,本研究確立了育苗過(guò)程中的合適溫度、鹽度,對(duì)波紋巴非蛤育苗工作確立了有利條件。但是,其他因素,如餌料、光照、pH值等影響波紋巴非蛤發(fā)育的環(huán)境因子的研究還不充分,需要深入研究。
[1] 陳堅(jiān),柯愛(ài)英,范景水,等.波紋巴非蛤生物學(xué)性狀及生態(tài)習(xí)性的初步觀察[J].浙江海洋學(xué)院學(xué)報(bào):自然科學(xué)版,2007,26(3):343-346.
[2] 諶素華,洪鵬志,章超樺.波紋巴非蛤肉及菠蘿汁營(yíng)養(yǎng)口服液的研制[J].食品科技,2004,30(8):59-61.
[3] 董曉靜,范秀萍,吳紅棉,等.響應(yīng)面法優(yōu)化波紋巴非蛤糖胺聚糖提取工藝的研究[J].現(xiàn)代食品科技,2009,25(12):1423-1426.
[4] 陳純馨,陳忻,岑佩強(qiáng),等.波紋巴非蛤生物活性肽的分離與提純研究[J].食品科學(xué),2009,30(20):143-145.
[5] 陳純馨,孫恢禮,陳曉剛,等.風(fēng)味蛋白酶酶解波紋巴非蛤制備小分子肽工藝研究[J].廣東農(nóng)業(yè)科學(xué),2011,38(20):5-8.
[6] Chen X,Sun H,Zhou D.Preparation of low-molecular-mass peptides by enzyme autolysis ofPaphiaundulata[J].Preparative Biochemistry Biotechnology,2010,40(4):286-293.
[7] Wu X,Tan J,Cai M,et al.Molecular cloning,characterization,and expression analysis of a heat shock protein (HSP) 70 gene fromPaphiaundulata[J].Gene,2014,543(2):275-285.
[8] 董曉靜,范秀萍,吳紅棉,等.波紋巴非蛤糖胺聚糖抗氧化作用的研究[J].食品工業(yè)科技,2012,33(6):181-184.
[9] 范秀萍,董曉靜,吳紅棉,等.波紋巴非蛤多糖對(duì)高脂模型小鼠血脂的影響[J].現(xiàn)代食品科技,2014,30(1):7-10.
[10] 范秀萍,吳紅棉,王婭楠,等.波紋巴非蛤糖蛋白的分離提取及體外清除羥自由基活性的研究[J].食品與發(fā)酵工業(yè),2008,34(1):138-140.
[11] 何小慶,曹文紅,章超樺,等.波紋巴非蛤蛋白酶解產(chǎn)物的抗氧活性及分子量分布研究[J].現(xiàn)代食品科技,2014,30(1):74-80.
[12] 林麗云.波紋巴非蛤活性肽的體外抗氧化活性[J].江蘇農(nóng)業(yè)科學(xué),2014,42(10):298-300.
[13] 林麗云,孫恢禮,肖婉娜,等.波紋巴非蛤活性肽的酶法提取及其抗氧化性研究[J].安徽農(nóng)業(yè)科學(xué),2010,38(20):10901-10902.
[14] 吳紅棉,范秀萍,雷曉凌,等.波紋巴非蛤氨基多糖的分離純化及其理化性質(zhì)的初步研究[J].食品與發(fā)酵工業(yè),2005,31(7):133-136.
[15] He X,Cao W,Pan G,et al.Enzymatic hydrolysis optimization ofPaphiaundulataand lymphocyte proliferation activity of the isolated peptide fractions[J].Journal of the Science of Food and Agriculture,2015,95(7):1544-1553.
[16] 黃松木,方火順,吳和平,等.云霄礁美海區(qū)波紋巴非蛤生物學(xué)特性及增殖研究[J].福建水產(chǎn),1984(3):14-19.
[17] 趙志江,李復(fù)雪.波紋巴非蛤Paphiaundulata精子發(fā)生的超微結(jié)構(gòu)[J].臺(tái)灣海峽,1992,11(3):238-243.
[18] 徐志雄.福建沿岸水域的巴非蛤[J].福建水產(chǎn),1987(2):44-46.
[19] 趙志江,李復(fù)雪,柯才煥.波紋巴非蛤的性腺發(fā)育和生殖周期[J].水產(chǎn)學(xué)報(bào),1991,28(1):1-8,26.
[20] 吳洪流,王紅勇,王珺.波紋巴非蛤性腺發(fā)育分期的研究[J].海南大學(xué)學(xué)報(bào):自然科學(xué)版,2002,20(1):41-47.
[21] 吳洪流.波紋巴非蛤性逆轉(zhuǎn)時(shí)生殖腺的組織學(xué)變化[J].海洋科學(xué),2002,26(1):5-8.
[22] 吳洪流,伍春雨,陳雪芬,等.波紋巴非蛤雄性生殖腺的組織學(xué)觀察[J].海南大學(xué)學(xué)報(bào):自然科學(xué)版,2000,18(2):169-174.
[23] 陳何東.波紋巴非蛤大規(guī)格苗種培育技術(shù)[J].中國(guó)水產(chǎn),2015(5):62-63.
[24] 王萬(wàn)東.東山灣云霄海區(qū)波紋巴非蛤生態(tài)習(xí)性和增養(yǎng)殖技術(shù)[J].漁業(yè)現(xiàn)代化,2007,34(4):33-34.
[25] 王萬(wàn)東.福建省東山灣云霄海區(qū)波紋巴非蛤養(yǎng)殖狀況監(jiān)測(cè)[J].養(yǎng)殖與飼料,2009,8(6):17-21.
[26] 王萬(wàn)東.波紋巴非蛤人工育苗技術(shù)的初步研究[J].養(yǎng)殖與飼料,2010,9(8):14-17.
[27] 徐小偉,張鵬飛,周龍,等.幾種環(huán)境因子對(duì)波紋巴非蛤胚胎發(fā)育的影響[J].福建水產(chǎn),2015,37(6):435-440.
[28] 徐小偉,張鵬飛,黃妙琴,等.波紋巴非蛤早期發(fā)育的掃描電鏡觀察[J].福建水產(chǎn),2015,37(4):263-269.
[29] 洪一川,呂小梅,張躍平,等.波紋巴非蛤人工育苗技術(shù)初探[J].福建水產(chǎn),2010,32(3):61-64.
[30] 陳志,高如承,胡青,等.溫度、鹽度和密度交互作用對(duì)波紋巴非蛤幼貝生長(zhǎng)的影響[J].海洋科學(xué),2013,37(8):55-60.
[31] 李俊輝,劉紅禮,杜曉東,等.鹽度和規(guī)格對(duì)波紋巴非蛤Paphiaundulata耗氧率和排氨率的影響[J].海洋學(xué)研究,2011,29(4):95-99.
[32] 李俊輝,葉尚芳,杜曉東,等.溫度和規(guī)格對(duì)波紋巴非蛤耗氧率和排氨率的影響[J].水生態(tài)學(xué)雜志,2010,3(5):96-99.
[33] 王冬梅,李春強(qiáng),彭明,等.鹽度、pH對(duì)波紋巴非蛤(Paphiaundulata)濾水率的影響[J].海洋通報(bào),2009,28(2):23-27.
[34] 張躍平,呂小梅,洪一川,等.波紋巴非蛤胚胎與浮游幼蟲(chóng)的形態(tài)發(fā)育和生長(zhǎng)特性[J].臺(tái)灣海峽,2011,30(4):546-550.
[35] 邵艷卿,張炯明,方軍,等.人工蓄養(yǎng)斧文蛤的生殖周期及早期發(fā)育[J].中國(guó)水產(chǎn)科學(xué),2017,24(1):82-90.
猜你喜歡
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2022年3期)2022-03-16 05:55:08
當(dāng)代陜西(2021年2期)2021-03-29 07:41:24
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語(yǔ)·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
媽媽寶寶(2017年3期)2017-02-21 01:22:28
紅領(lǐng)巾·萌芽(2016年1期)2016-09-10 07:22:44
中國(guó)塑料(2016年3期)2016-06-15 20:30:00
