蛭石引發對鹽脅迫下番茄種子萌發、幼苗生長及生理特性的影響
2018-03-27 01:36:41楊若鵬李爺福
江蘇農業科學 2018年4期
關鍵詞:生長
楊若鵬, 李爺福, 李 杰
(1.紅河學院生命科學與技術學院,云南蒙自 661199; 2.云南省高校農作物優質高效栽培與安全控制重點實驗室,云南蒙自 661199)
番茄(LycopersiconesculentumMill)是茄科番茄屬一年生或多年生草本植物,株高0.6~2.0 m。番茄的起源中心是南美洲的安第斯山地帶,目前在全球普遍種植,而且種植面積逐年增加。
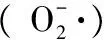
1 材料與方法
1.1 試驗材料
供試番茄(LycopersiconesculentumMill)品種為福寶、東方紅,購買于云南省紅河州蒙自市種子咨詢服務部,蛭石由紅河學院校內實習基地提供。
1.2 試驗設計
1.2.1 種子消毒 用10%次氯酸鈉溶液在大燒杯中浸泡挑選出的飽滿番茄種子進行消毒,消毒后用自來水沖洗3遍。
1.2.2 種子引發 在開始試驗前進行預備試驗,以確定引發各物質的比例,按體積比(蛭石 ∶蒸餾水 ∶種子=1.5 ∶1 ∶1)拌勻后于15 ℃、無光照條件下引發1 d,記作T處理,沒有引發的番茄種子作為對照(CK)。
1.2.3 回干 蛭石引發處理之后用相應的細篩將種子篩出,在鼓風干燥箱中,于(25±2)℃干燥至未引發時種子中的含水量。
1.2.4 種子發芽及鹽脅迫設計 將引發、未引發的種子在光照培養箱內發芽,恒定溫度設為26 ℃,每天光—暗周期為 12 h—12 h,光照度為4 000 lx。將引發的種子(T)和未引發的種子(CK)在加有雙層濾紙消毒的培養皿(直徑120 mm)內發芽,每個處理3個重復,每個重復設150粒種子,分別用0、100、200 mmol/ L NaCl溶液保持發芽盒內濕度恒定。待90%以上種子露白后移栽至裝有復合基質的穴盤內,復合基質由草炭、蛭石、珍珠巖按體積比3 ∶1 ∶1組成。在江河學院溫室大棚內栽培,每次澆水時用相應濃度的NaCl溶液進行脅迫處理。
1.3 指標測定與方法
本試驗中的相應計算公式如下:
發芽率=第7天供試種子的發芽數/供試種子數×100%;
發芽勢=第2天供試種子的發芽數/供試種子數×100%;
發芽指數(GI)=∑Gt/Dt(Gt指在td的發芽數,Dt指發芽時間)。
SOD活性的測定用氮藍四唑(NBT)法,以抑制NBT光化還原的50%為1個酶活單位(U)[13];POD活性的測定用愈創木法,以1 min內D470 nm變化0.01為1個過氧化物酶活性單位(U)[13];植物體內游離脯氨酸的含量用磺基水楊酸法測定[13];用蒽酮比色法測定可溶性糖含量[13];用考馬斯亮藍 G-250 法測定可溶性蛋白質含量[13];CAT活性用紫外-可見分光光度法測定,以1 min內D240 nm減少0.1的酶量為1個酶活單位(U)[14];用硫代巴比妥酸(TBA)法測定丙二醛含量[14]。
1.4 數據處理
試驗得到的數據采用SPSS軟件進行方差分析,并對平均數進行Duncan’s多重比較.
2 結果與分析
2.1 番茄種子萌發特性的變化
由表1可知,引發處理的番茄種子和未引發處理的番茄種子的各項指標均隨著NaCl濃度的升高而顯著降低,說明番茄種子的萌發在受到NaCl脅迫后受阻,而且隨著脅迫濃度的升高,迫害更加明顯。但是經蛭石引發處理的番茄種子的各指標與未引發處理(CK)相比有顯著提高,表明在一定程度上引發后緩解了NaCl的脅迫。福寶番茄的發芽勢、發芽指數、活力指數等高于東方紅番茄,但差異不明顯。所以可以看出,通過蛭石引發處理可以顯著改善番茄種子在鹽堿脅迫逆境中的生長狀況,提高其發芽率及出苗速度。

表1 蛭石引發對NaCl脅迫下番茄種子萌發特性的影響
注:T為蛭石引發種子處理,CK為非蛭石引發處理;同一品種不同處理間標有不同小寫字母表示差異達5%水平。下表同。
2.2 番茄幼苗生長情況的變化
由表2可知,隨著NaCl濃度的增加,番茄幼苗長勢均逐漸減弱,其各項指標都有顯著的降低,尤其在NaCl濃度為200 mmol/L時,幼苗的生長基本被抑制。但是經過蛭石引發后的番茄幼苗根系長、株高、鮮質量、干質量等大都顯著高于未引發的。但在200 mmol/L NaCl濃度處理下,兩者的根長、幼苗鮮質量、干質量無顯著差異。以上結果表明,通過蛭石引發處理可以從一定程度上改善番茄幼苗應對不良環境的能力,可以顯著提升番茄幼苗對NaCl脅迫的抵抗能力。
2.3 番茄幼苗保護性酶活性的變化
由表3可知,在沒有NaCl脅迫時,蛭石引發處理和未引發處理的CAT活性和丙二醛含量無顯著差異。隨著NaCl濃度升高,兩者的CAT活性和MDA含量均隨之升高(在0~100 mmol/L NaCl濃度范圍內),但是引發處理的CAT活性和MDA含量均顯著低于未處理的CAT活性和MDA含量。產生CAT的速率及MDA含量可以體現植物體在受到逆境脅迫時膜脂的過氧化程度。上述結果表明,經過蛭石引發后可以有效地緩解番茄幼苗膜脂化的程度,有效抵制了NaCl脅迫對細胞膜的傷害,提升了番茄種子的耐鹽性。
由表3還可以看出,番茄幼苗葉片中的超氧化物歧化酶活性均隨NaCl脅迫濃度的增大而顯著提升,當NaCl濃度達到200 mmol/L時又降低,在NaCl濃度為100 mmol/L時,活性達到最大值。在0~200 mmol/L NaCl濃度下,引發處理的番茄幼苗葉片SOD活性均顯著高于相應對照。隨著NaCl濃度的增加,引發處理和未引發處理番茄葉片POD活性均逐漸增大,幼苗葉片POD活性在200 mmol/L NaCl處理后開始降低,在0~200 mmol/L NaCl濃度下,引發處理的葉片POD活性均顯著高于相應對照。結果表明,通過引發處理可以顯著提高番茄幼苗各保護酶的活性,使其在受鹽堿的脅迫時能更好地消除對自身不利的因素,從而體現出較強長勢。

表2 蛭石引發對NaCl脅迫下番茄幼苗生長的影響
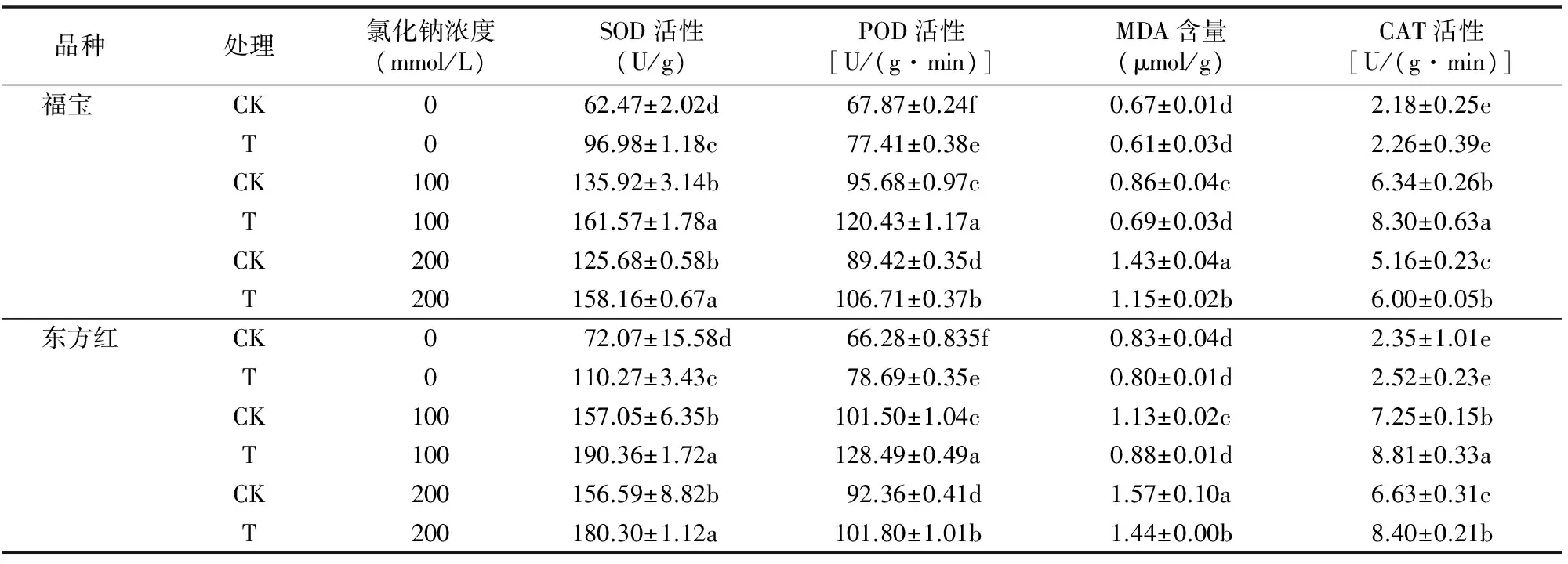
表3 蛭石引發對NaCl脅迫下番茄幼苗保護性酶活性的影響
注:SOD、POD活性與MDA含量均是對于鮮質量而言的。
2.4 番茄幼苗滲透調節物質的變化
由表4可知,在沒有NaCl脅迫時,番茄幼苗葉片中的游離脯氨酸含量及可溶性糖含量無顯著差異,而番茄幼苗葉片中的可溶性蛋白質含量在經蛭石引發處理后有顯著提高。游離脯氨酸、可溶性糖及可溶性蛋白質3種滲透調節物的含量在NaCl濃度增大時都顯著升高(僅NaCl濃度為100 mmol/L時游離脯氨酸含量升高不顯著)。當NaCl濃度在 200 mmol/L 時可溶性蛋白質含量又顯著低于100 mmol/L NaCl處理,且蛭石引發處理過的番茄幼苗葉片中3種滲透調節物含量均顯著高于相應未引發的。由上述結果可得出,經過蛭石引發能夠顯著提高番茄幼苗葉片內的滲透調節物質含量,使得細胞內形成較低的水勢,提高幼苗細胞在高濃度的鹽堿環境中的吸水能力,有效地維持細胞內水分平衡,減輕了鹽害,從而提高了耐鹽性。
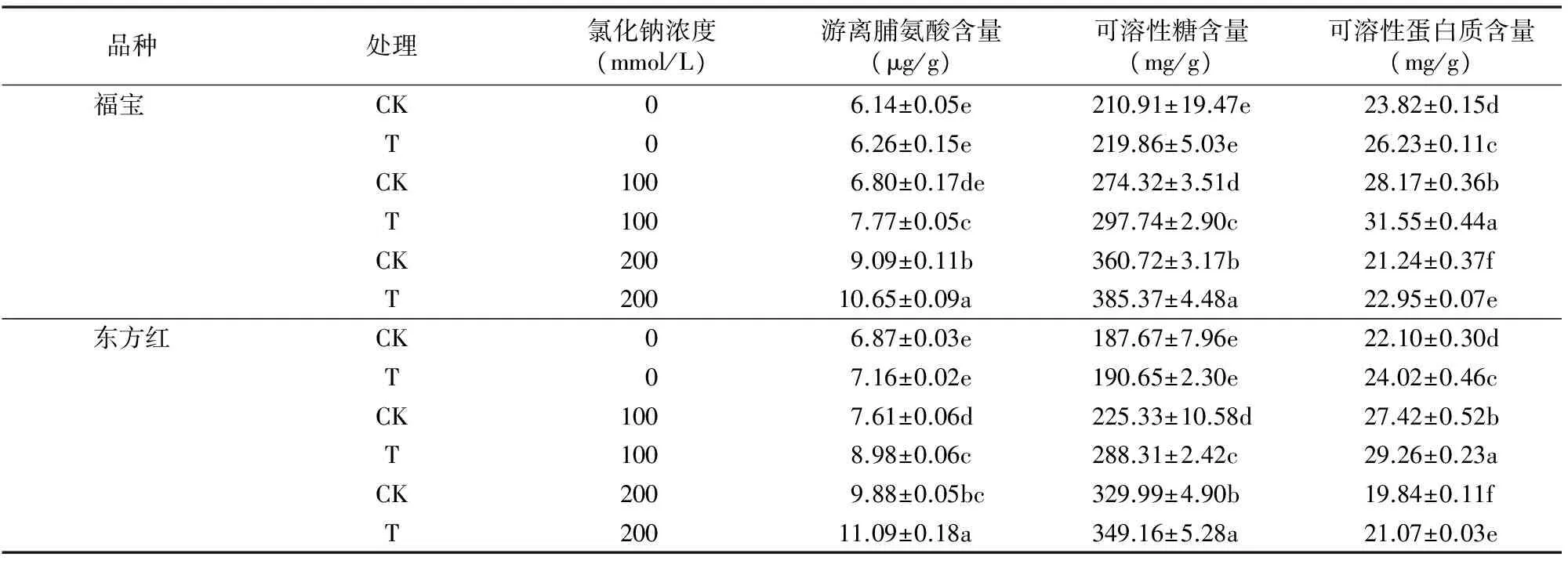
表4 蛭石引發對NaCl脅迫下番茄幼苗滲透調節物質的影響
注:游離脯氨酸、可溶性糖、可溶性蛋白質均為鮮質量含量。
3 討論
3.1 蛭石引發對NaCl脅迫下番茄種子萌發和幼苗生長的影響
鹽脅迫對種子萌發的抑制,是由土壤中水勢降低造成的[15]。鹽脅迫下,如果種子外界水分濃度過高,會導致吸水困難,細胞膜透性增大,細胞內溶質外滲而阻礙了種子的萌發[16]。本試驗結果表明,通過蛭石引發可以提高番茄種子在NaCl脅迫下的發芽能力和幼苗生長指標。本試驗中番茄的4個萌發特性及4個幼苗生長指標都隨著NaCl濃度的升高而顯著降低,而蛭石引發處理后各指標均有顯著提高。因此可以看出,引發可以大大提高NaCl脅迫下番茄種子的出苗率,可以有效緩解NaCl脅迫對種子萌發及幼苗生長的影響,從而提高種子的出苗率[17-18]。
3.2 蛭石引發對NaCl脅迫下番茄種子幼苗保護酶活性的影響
植物在受到外界鹽脅迫等逆境時,其體內會產生活性氧簇(ROS),使得膜脂過氧化作用在細胞內發生,而作為植物體細胞膜脂過氧化的主要產物MDA,在植物體受到外界鹽堿等逆境時,其含量會升高,而作為清除細胞內ROS的保護酶,SOD、POD、CAT活性也會隨著ROS的增加而升高。在本試驗中,與非鹽脅迫相比,100 mmol/L NaCl下番茄葉片中的SOD、POD和CAT活性升高,但是經過蛭石引發處理后,MDA含量升高程度有所減緩,主要可能是蛭石引發處理后提高了番茄幼苗葉片細胞膜的穩定性和完整性,從而減少了MDA的產生。當NaCl濃度達到200 mmol/L時,番茄葉片中的SOD、POD、CAT活性又有所下降,可能是由于高濃度的鹽環境抑制了番茄的生長,這與張士功等研究水楊酸對小麥種子萌發和幼苗生長過程中鹽害的緩解作用相一致[19]。
3.3 蛭石引發對NaCl脅迫下番茄種子幼苗滲透調節作用的影響
當植物體生長在鹽堿等逆境中時,植物體根系周圍土壤內會形成高濃度的環境,所以導致外界滲透勢降低,造成植物體吸水困難的生理現象。此時植物自身的滲透調節尤為重要,而作為滲透調節指標的可溶性蛋白質、游離脯氨酸和可溶性糖就起著重要作用[22-23]。有關研究表明,鹽脅迫下有關外源多胺能通過抑制淀粉酶和蛋白酶的活性,提高可溶性糖和蛋白質的量,從而影響生長[24]。可溶性蛋白質含量的提高增加了細胞功能性蛋白的含量和滲透勢,有利于提高植物的抗逆性[10]。在本試驗中,番茄幼苗葉片中的滲透調節物含量在NaCl濃度增加時顯著升高,而蛭石引發處理過后也顯著提高了滲透調節物質含量。其結果與前人的研究結果一致,可溶性糖等作為細胞調節物質和細胞的滲透勢密切相關,可溶性糖含量提高,增加了細胞的滲透勢,提高了植物的抗逆性[25]。經引發的番茄幼苗葉片中的可溶性蛋白質、游離脯氨酸和可溶性糖含量與未處理番茄幼苗葉片相比均有顯著提高,尤其在100 mmol/L NaCl脅迫時最顯著,表明蛭石引發提高了NaCl脅迫下番茄幼苗的滲透調節作用,使番茄幼苗更正常地生長。
4 結論
綜上所述,2個品種番茄種子在受到NaCl脅迫時,萌發特性、保護酶活性、滲透調節物質含量通過蛭石引發處理時均表現出顯著優勢,使得植株生長更具耐鹽性,說明蛭石引發顯著增強了番茄種子及幼苗對鹽環境的抗性。蛭石引發顯著提高CAT、SOD、POD保護酶活性與可溶性蛋白質、可溶性糖、游離脯氨酸含量,而MDA含量顯著降低可維持細胞的穩定性,提高番茄種子的萌發率及促進幼苗的生長。2個不同品種間的相同處理間表現也存在一定的差異性,從試驗結果來看,福寶品種的耐鹽性比東方紅強,且效果在100 mmol/L NaCl模擬脅迫時最明顯。因此可以得出,在鹽堿地區2個品種的選擇上,推薦種植福寶。
[1]阮松林,薛慶中. 植物的種子引發[J]. 植物生理學通訊,2002,38(2):198-202.
[2]Yang X H,Jiang W J,Wei M,et al. Review on plant response and resistance mechanism to salt stress[J]. Journal of Shandong Agriculture University,2006,37(2):302-305.
[3]郟艷紅,劉仲齊,金鳳媚,等. 西紅柿耐鹽性研究進展[J]. 天津農業科學,2006,12(2):20-23.
[4]邱倩倩,李 明,姚東偉,等. 蛭石引發對NaCl單鹽脅迫下辣椒種子萌發和幼苗抗氧化特性的影響[J]. 上海農業學報,2009,25(3):47-50.
[5]常 瑤,李 明,姚東偉,等. 蛭石引發對高溫脅迫下小白菜種子萌發和幼苗生長的影響[J]. 上海農業學報,2013,29(6):40-43.
[6]付愛飛. 硒提高西紅柿耐鹽性的生理機制研究[D]. 石河子:石河子大學,2008.
[7]張 毅. 亞精胺對西紅柿幼苗鹽堿脅迫的緩解效應及調控機理[D]. 楊凌:西北農林科技大學,2013.
[8]吳鳳芝,劉 德,奕非時. 大棚土壤連作年限對黃瓜產量及品質的影響[J]. 中國蔬菜,1999,30(1):245-248.
[9]陳 陽. 新疆鹽生植物生理生態適應性及硅提高植物抗鹽作用機制的研究[D]. 蘭州:甘肅農業大學,2002.
[10]高艷明,李建設,張 敏. 茄子種子基質引發試驗[J]. 西北農業學報,2006,15(2):148-151.
[11]李建設,高艷明,馮 艷. 蛭石和珍珠巖基質引發對洋蔥種子發芽率影響[J]. 北方園藝,2006(6):16-17.
[12]陳雙燕,韓建國,王贇文,等. 蛭石引發對結縷草種子發芽率和發芽速度的影響[J]. 草地學報,2007,15(3):254-258.
[13]李關榮,李天俊,馮建成. 生物化學實驗教程[M]. 北京:中國農業大學出版社,2011:127-133.
[14]張志良,瞿偉菁,李小方. 植物生理學實驗指導[M]. 北京:高等教育出版社,2008:227-229.
[15]Netondo G W,Onyango J C,Beck E. Sorghum and salinity: I. Response of growth, water relations,and ion accumulation to NaCl salinity[J]. Crop Science,2004:101-106.
[16]閆先喜,馬小杰,邢樹平,等. 鹽脅迫對大麥種子細胞膜透性的影響[J]. 植物學通報,1995(增刊1):53-54.
[17]楊小環,馬金虎,郭數進,等. 種子引發對鹽脅迫下高粱種子萌發及幼苗生長的影響[J]. 中國生態農業學報,2011,19(1):103-109.
[18]賀長征,胡 晉,朱志玉,等. 混合鹽引發對水稻種子在逆境條件下發芽及幼苗生理特性的影響[J]. 浙江大學學報(農業與生命科學版),2002,28(2):175-178.
[19]張士功,高吉寅,宋景芝. 水楊酸對小麥種子萌發和幼苗生長過程中鹽害的緩解效應[J]. 中國農業科學,1998,31(4):90-91.
[20]侯彩霞,湯章城. 細胞相容性物質的生理功能及其作用機制[J]. 植物生理學報,1999,35(1):1-7.
[21]秦峰梅,張紅香,武 祎,等. 鹽脅迫對黃花苜蓿發芽及幼苗生長的影響[J]. 草業學報,2010,19(4):71-78.
[22]Krishnamurthy R. Amelioration of salinity effect in salt tolerant rice (OryzasativaL.) by foliar application of putrescine[J]. Plant & Cell Physiology,1991,32(5): 699-703.
[23]陳雙燕,韓建國,王赟文,等. 蛭石引發對結縷草種子發芽率和發芽速度的影響[J]. 草地學報,2007,15(3):254-258.
[24]Shan C J. Effect of soil drought on physiological characteristics of winter wheat Luomai 9133 during jointing stage[J]. Journal of Triticeae Crops,2007,27(5):880-883.
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
