水稻紋枯病菌RsPhm基因的克隆及其表達分析
2018-04-09 08:34:37江紹鋒王陳驕子舒燦偉周而勛
中國水稻科學 2018年2期
關鍵詞:水稻
江紹鋒 王陳驕子 舒燦偉 周而勛
?
水稻紋枯病菌基因的克隆及其表達分析
江紹鋒 王陳驕子 舒燦偉 周而勛*
(華南農業大學 農學院/廣東省微生物信號與作物病害防控重點實驗室, 廣州 510642;)
【目的】為了闡明苯酚2-單加氧酶基因()在水稻紋枯病菌(AG-1ⅠA)黑化中的功能,【方法】采用常規PCR和RT-PCR技術對該基因進行克隆和生物信息學分析,通過熒光定量PCR(qRT-PCR)技術檢測在兒茶酚脅迫下該基因的相對表達量。【結果】生物信息學分析結果表明,基因的DNA和cDNA全長序列分別為2 628 bp和1 983 bp,編碼660個氨基酸。系統進化樹分析顯示,基因在立枯絲核菌()不同融合群中具有較近的親緣關系,在不同真菌之間在進化上具有一定保守性。通過qRT-PCR技術分析了在不同濃度兒茶酚脅迫下水稻紋枯病菌基因的轉錄表達情況。外源兒茶酚能提高基因的表達量,在12.5 μg/mL濃度下表達量最高,極顯著上調35.7倍,在25 μg/mL和50 μg/mL濃度下表達量分別上調19.1倍和28.4倍,但在100 μg/mL濃度下表達量僅上調2.1倍。【結論】獲得了基因全長序列,了解了其基本生物學信息,明確了其在兒茶酚脅迫下的表達模式。研究結果為科學、系統地闡明水稻紋枯病菌基因調控黑色素形成機制奠定了理論基礎。
水稻紋枯病菌;苯酚2-單加氧酶基因;基因克隆;表達分析
由立枯絲核菌(Kühn) AG-1 ⅠA融合亞群引起的水稻紋枯病是世界范圍內分布最為廣泛的水稻三大真菌病害之一。立枯絲核菌()是一種經濟上非常重要的土傳性病原真菌,其分布范圍廣泛,寄主類型多種多樣,除為害水稻外,還可為害馬鈴薯、玉米、小麥、大豆和甘藍等260多種植物[1-4]。
加氧酶指能催化氧分子加入到有機物質中的酶,是芳香化合物降解過程中的重要酶類,它廣泛存在于各種生命活動的新陳代謝過程中。加氧酶按照催化氧原子的個數可以劃分為兩類:將兩個氧原子都催化反應進入底物的雙加氧酶和只催化一個氧原子進入底物的單加氧酶[5]。苯酚2-單加氧酶(phenol 2-monooxygenase,Phm)是一種單加氧的氧化還原酶,能催化苯酚、NADPH和H+在有氧條件生產兒茶酚、NADP+和水[6]。微生物對芳香族化合物的降解可以分為苯環的激活以及對被激活苯環的開環兩個過程,加氧酶參與芳香族化合物、多環芳烴、農藥等多個種類的污染物(如六六六、甲苯、二甲苯、萘和蒽等)的降解代謝途徑[6-8]。苯酚2-單加氧酶是苯酚降解中的關鍵芳環加氧酶。首先,通過苯酚羥基化作用下生成兒茶酚;然后兒茶酚在C12O和C23O作用下分別通過鄰位開環和間位開環形式被進一步降解[9]。
黑色素是植物病原真菌的主要毒力因子和抗逆因子。許多植物病原真菌的菌絲、分生孢子和菌核都能產生黑色素,從而以多種方式增強真菌的毒力和抗逆性[10-11]。大多數致病真菌可通過內源性前體物質代謝合成黑色素,但新生隱球菌()需要添加外源性左旋多巴(-DOPA)才能形成黑色素[12]。兒茶酚是合成黑色素的前體物質,已經在玉蜀黍黑粉菌()的厚垣孢子內發現[13]。本實驗室在前期研究中發現在培養基中添加兒茶酚有利于促進水稻紋枯病菌黑色素的生物合成和積累,從而加快細胞壁的黑化和增厚[14]。目前,尚未見到關于水稻紋枯病菌苯酚2-單加氧酶()基因的研究報道。本研究通過對基因的克隆及其在兒茶酚誘導下的表達分析,為深入研究基因調控水稻紋枯病菌黑色素形成機制奠定理論基礎。
1 材料與方法
1.1 供試菌株和培養基配制
水稻紋枯病菌(AG-1ⅠA) GD-118菌株是本實驗室分離和保存的強致病力菌株[15]。
馬鈴薯葡萄糖瓊脂培養基(potato dextrose agar,PDA):馬鈴薯200.0 g,葡萄糖20.0 g,瓊脂粉18.0 g,雙蒸水1 000.0 mL。馬鈴薯葡萄糖液體培養基(potato dextrose broth,PDB):馬鈴薯200.0 g,葡萄糖20.0 g,雙蒸水1 000.0 mL。
LBA固體培養基(luria-bertani agar,LBA):胰蛋白胨10.0 g,酵母提取物5.0 g,NaCl 10.0 g,調pH至7.4,瓊脂粉15.0 g,雙蒸水1 000.0 mL。LBB液體培養基(luria-bertani broth,LBB):胰蛋白胨10.0 g,酵母提取物5.0 g,NaCl 10.0 g,調pH值至7.4,雙蒸水1 000.0 mL。
1.2 主要試劑
真菌DNA提取試劑盒、RNA提取試劑盒、克隆載體-T1、-T1感受態細胞、RT-PCR試劑盒、普通酶和高保真酶均購自北京全式金生物技術有限公司。兒茶酚、氨芐青霉素鈉、IPTG和X-Gal購自Sigma公司。引物(表1)合成和測序由生工生物工程(上海)股份有限公司完成。
1.3 菌絲體培養與收集
水稻紋枯病菌GD-118菌株在PDA平板上培養2 d后,用內徑5.0 mm的打孔器在菌落邊沿打取菌絲塊,在每個裝有150 mL PDB培養液(分別含0,12.5,25,50,100 μg/mL兒茶酚)的三角瓶中接入10塊菌絲塊,28℃恒溫下搖床振蕩培養3 d后,用無菌濾紙過濾收集菌絲體,無菌水將菌絲體洗滌2~3次,菌絲體用多層滅菌濾紙吸干表面水分后,放入―20℃冰箱中保存備用。
1.4 DNA和RNA的提取
水稻紋枯病菌GD-118 DNA和RNA的提取按照真菌DNA提取試劑盒和RNA提取試劑盒的說明書進行。在采用瓊脂糖凝膠電泳檢測其完整性、紫外分光光度測定其純度和含量,提純后的DNA和RNA樣品分別保存于―20℃冰箱和―80℃超低溫冰箱中,備用。
1.5 RsPhm基因DNA和cDNA克隆
根據水稻紋枯病菌基因組和轉錄組數據庫[16],用Vector NTI 軟件設計一對水稻紋枯病菌基因特異性引物,引物序列為DF01905和DR01905(表1)。以總RNA為模板,利用RT-PCR試劑盒進行RT-PCR擴增得到cDNA。以基因組DNA和cDNA為模板,分別采用北京全式金生物技術有限公司的普通酶和高保真酶進行PCR擴增,擴增體系如下:PCR混合液25 μL,10 μmol/L DF01905和DR01905引物各1 μL,模板1 μL,普通酶或高保真酶1 μL,再加雙蒸水22 μL。PCR程序如下:94℃下預變性3 min;94℃下變性30 s,55℃下退火30 s,72℃下延伸3 min,35個循環后72℃下延伸10 min。PCR產物保存于4℃冰箱。PCR產物經回收、連接,化學法轉入到-T1感受態細胞,經過藍白斑篩選及菌液PCR電泳鑒定,篩選陽性克隆。取5 μL PCR產物在1%瓊脂糖凝膠上進行電泳,加入1 μL 6×上樣緩沖液,在5~8 V/cm電壓條件下電泳30 min,于凝膠成像系統下觀察分析并拍照留存。陽性克隆的測序由生工生物工程(上海)股份有限公司完成。測序結果利用Conting Express軟件進行拼接,并利用NCBI在線工具對水稻紋枯病菌基因進行序列基本結構和特征分析。
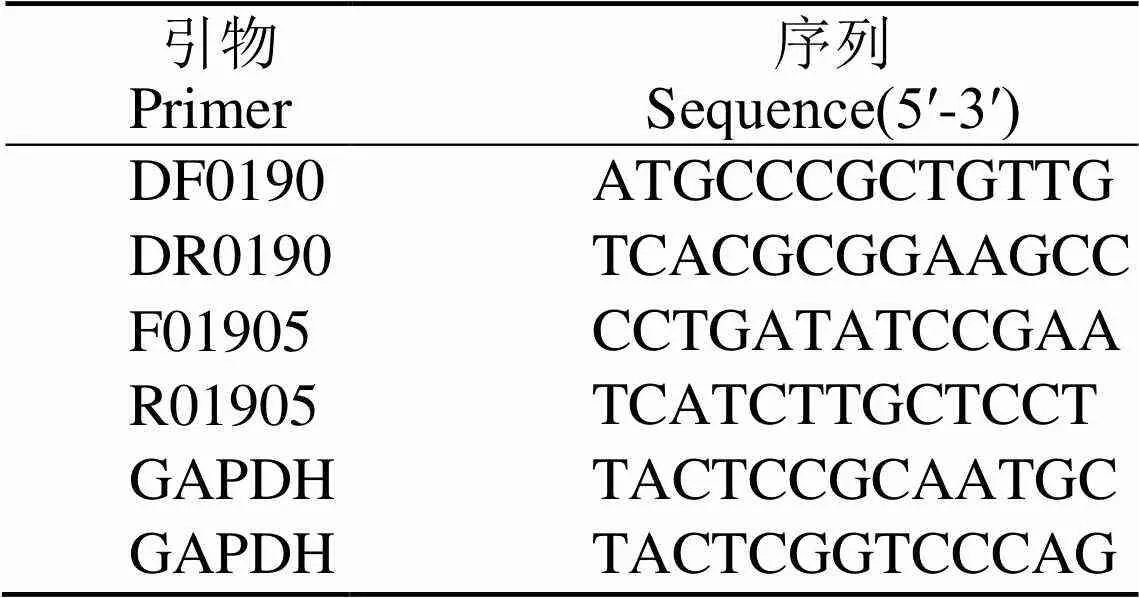
表1 本研究使用的引物序列
1.6 RsPhm基因生物信息學分析
通過在線程序和軟件對水稻紋枯病菌基因所編碼的蛋白質氨基酸序列進行生物信息學預測和分析,預測和分析參數有RsPhm蛋白質理化性質、信號肽、跨膜結構、二級結構、三維結構和系統進化樹分析,具體分析軟件如下:
1)蛋白質理化性質預測
http://web.expasy.org/protparam/;
2)信號肽的預測
http://www.cbs.dtu.dk/services/SignalP/;
3)跨膜結構預測與分析
http://www.ch.embnet.org/software/TMPRED_form.html;
4)蛋白質二級結構預測
http://imtech.res.in/raghava/apssp/;
5)蛋白質三維結構的預測
https://swissmodel.expasy.org/interactive;
6)系統進化樹分析
采用MEGA 4對水稻紋枯病菌RsPhm蛋白質與其他真菌RsPhm蛋白質的進化關系進行分析。
1.7 RsPhm基因定量表達分析
以水稻紋枯病菌甘油醛3-磷酸()基因作為內參基因,檢測水稻紋枯病菌在不同濃度兒茶酚處理下基因在轉錄水平上的相對表達量。RNA樣品用DNaseⅠ進行預處理后,用反轉錄試劑盒合成單鏈cDNA作為模板。基于已克隆的水稻紋枯病菌基因的cDNA序列,設計的一對引物為F01905和R01905;以基因為內參,引物為GAPDH和GAPDHR(表1)。qRT-PCR體系如下:SYBR Green PCR Master Mix為10 μL、10 μmol/L F01905和R01905引物各1 μL、不同樣本的cDNA(稀釋5倍)為2.0 μL,再加雙蒸水6 μL,包括不加cDNA模板為陰性對照。在Bio-Rad CFX儀器上進行實時定量(qRT)PCR,反應程序如下:預變性95℃ 2 min;95℃變性15 s;60℃退火15 s,72℃延伸30 s,40個循環收集熒光。內參基因qRT-PCR體系,除引物不同外,其余參數同基因。數據收集和分析采用Bio-Rad CFX Manager Sofware 2.0進行分析,每個樣品3次重復。
2 結果與分析
2.1 RsPhm基因核苷酸和氨基酸序列分析
將PCR產物純化回收、連接轉化、測序,對測序結果進行拼接并去掉載體序列,得到DNA和cDNA全長序列分別為2 628 bp和1 983 bp,與瓊脂糖凝膠電泳結果一致(圖1)。用DNAStar軟件分析篩選獲得的基因cDNA序列及其推導的氨基酸序列,以ATG為起始密碼子,TAA為終止密碼子,共編碼660個氨基酸(圖2)。經生物信息學分析,基因編碼蛋白質分子式為C3226H5056N908O970S23;相對分子質量為72 818.51 ku,理論等電點為5.94,含堿性氨基酸74個,酸性氨基酸83個,疏水性氨基酸230個,極性氨基酸149個,屬于酸性親水性蛋白。RsPhm蛋白不含信號肽,預測其為非分泌蛋白。RsPhm蛋白存在5個跨膜螺旋結構域,由3個內到外和2個外到內的雙向跨膜區域組成,屬于跨膜蛋白。利用SWISS-MODEL網站對基因編碼的蛋白質進行三維結構預測,發現RsPhm蛋白的三維結構主要由α-螺旋、β-折疊和不規則卷曲組成,并呈較均勻分布(圖3)。
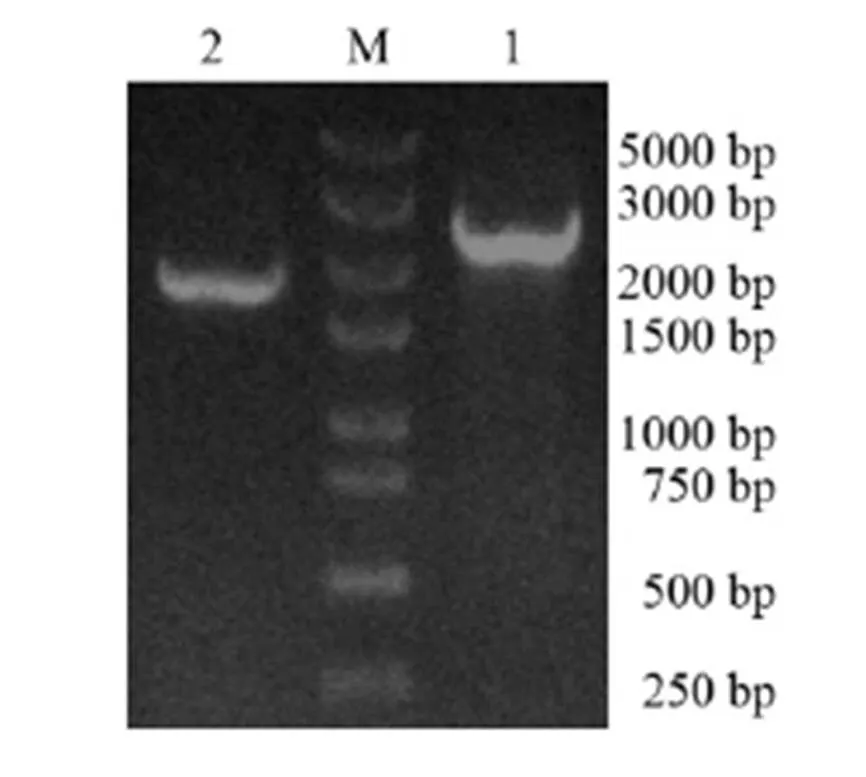
M―250 bp DNA標記; 1―以DNA為模板的RsPhm基因PCR產物; 2―以cDNA為模板的RsPhm基因PCR產物。
Fig. 1. PCR products ofgene in.
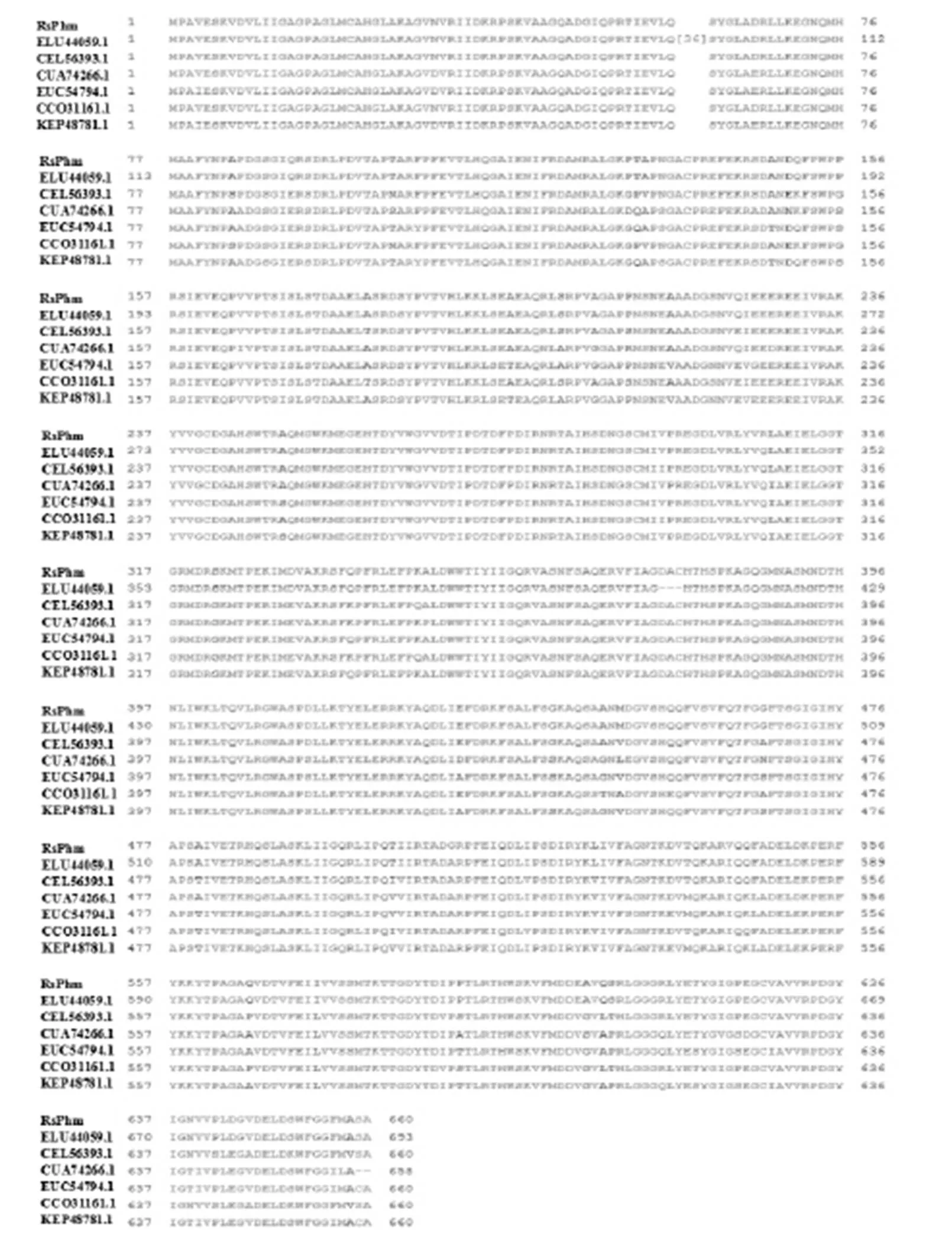
RsPhm為本研究結果,R. solani AG-1ⅠA、R. solani AG-1ⅠB、R. solani AG 22ⅢB、R. solani AG-3、R. solani AG-1ⅠB和R. solani 123E的NCBI GenBank注冊號分別為ELU44059.1、CEL56393.1、CUA74266.1、EUC54794.1、CCO31161.1和KEP48781.1。
Fig. 2. Comparison of amino acid sequences of RsPhm proteins in different anastomosis groups of
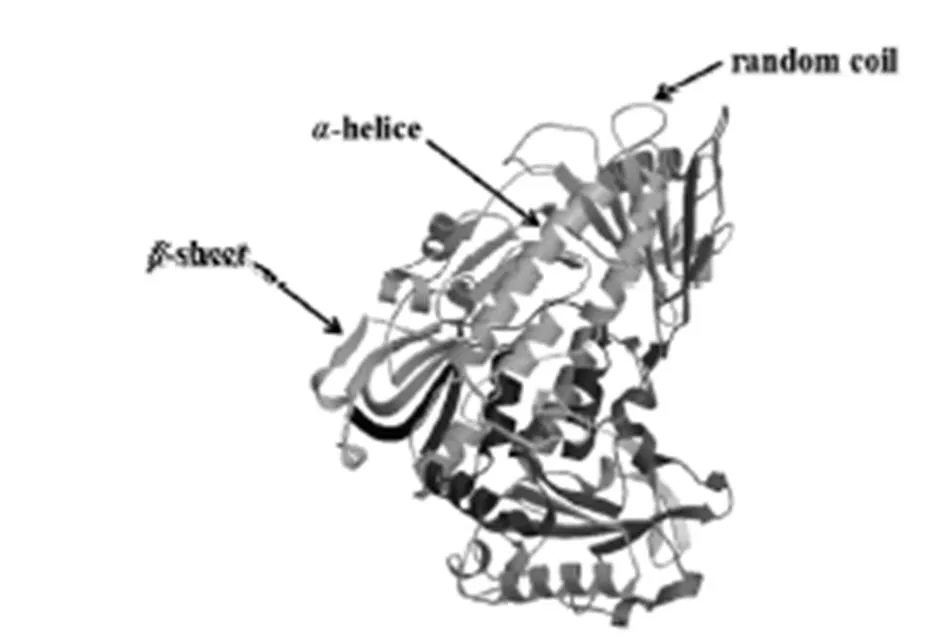
圖3 水稻紋枯病菌RsPhm蛋白三維結構預測
Fig. 3. Three-dimensional structure prediction of RsPhm protein inAG-1 IA.
2.2 RsPhm蛋白氨基酸序列系統進化樹分析
通過NCBI網站分析工具,對立枯絲核菌與其他近緣真菌之間的RsPhm蛋白氨基酸序列同源性進行比較。結果顯示,本研究的水稻紋枯病菌(AG-1ⅠA)與立枯絲核菌()其他融合群或亞群的氨基酸同源性最高,均達到91%以上;其中與同一融合亞群(AG-1ⅠA, ELU44059.1)另一個菌株的同源性達94 %,與其他菌株或融合群的(123E、AG-1ⅠB、AG22ⅢB和AG-3)的同源性分別為93%、93%、91%和91%。而與離心射脈革菌()、多葉奇果菌()、白腐菌()、白環菇(sp. SymC. cos)、真姬菇()的氨基酸同源性分別為54%~59%、57%、55%,56%和54%,同源性較低。
為比較立枯絲核菌RsPhm蛋白氨基酸序列與其他近緣真菌RsPhm蛋白氨基酸序列的親緣關系,本研究從GeneBank中查詢得到立枯絲核菌不同融合群菌株與離心射脈革菌、多葉奇果菌、白腐菌、白環菇和真姬菇的RsPhm蛋白氨基酸序列,與本研究克隆出來的RsPhm蛋白氨基酸序列進行比對,隨后利用MEGA 4軟件N-J法構建系統進化樹,Bootstrap自舉值取1 000 次,顯示不同種屬真菌的進化及親緣關系(圖4)。系統進化樹分析結果表明本研究克隆的RsPhm蛋白氨基酸序列和水稻紋枯病菌AG-1ⅠA其他菌株RsPhm蛋白氨基酸的序列同源性最高,同屬擔子菌門(Basidiomycota),同時立枯絲核菌不同的融合群菌株獨立進化為一支,表現出的親緣關系較其他真菌門的真菌更近,而與親緣關系較遠的其他真菌同源性則較低,與分類學地位及傳統形態學分類結果一致,因而推測不同真菌種類的基因在進化上具有一定的保守性。
2.2 兒茶酚誘導下RsPhm基因相對定量表達分析
對水稻紋枯病菌基因在不同濃度兒茶酚誘導下的轉錄表達情況進行了qRT-PCR分析,結果表明,在12.5~100 μg/mL濃度范圍內的兒茶酚普遍能誘導基因上調表達。與對照相比,在12.5 μg/mL兒茶酚誘導下表達量相對最高,極顯著上調35.7倍;在25 μg/mL和50 μg/mL兒茶酚處理下上調表達量分別為19.1倍和28.4倍;但在100 μg/mL兒茶酚處理下上調表達量僅2.1倍。究其原因,可能是高濃度兒茶酚抑制水稻紋枯病菌生長,進而負調控基因的表達(圖5)。兒茶酚是水稻紋枯病菌黑色素形成的前體合成物質,推測基因表達水平與黑色素形成關系密切。
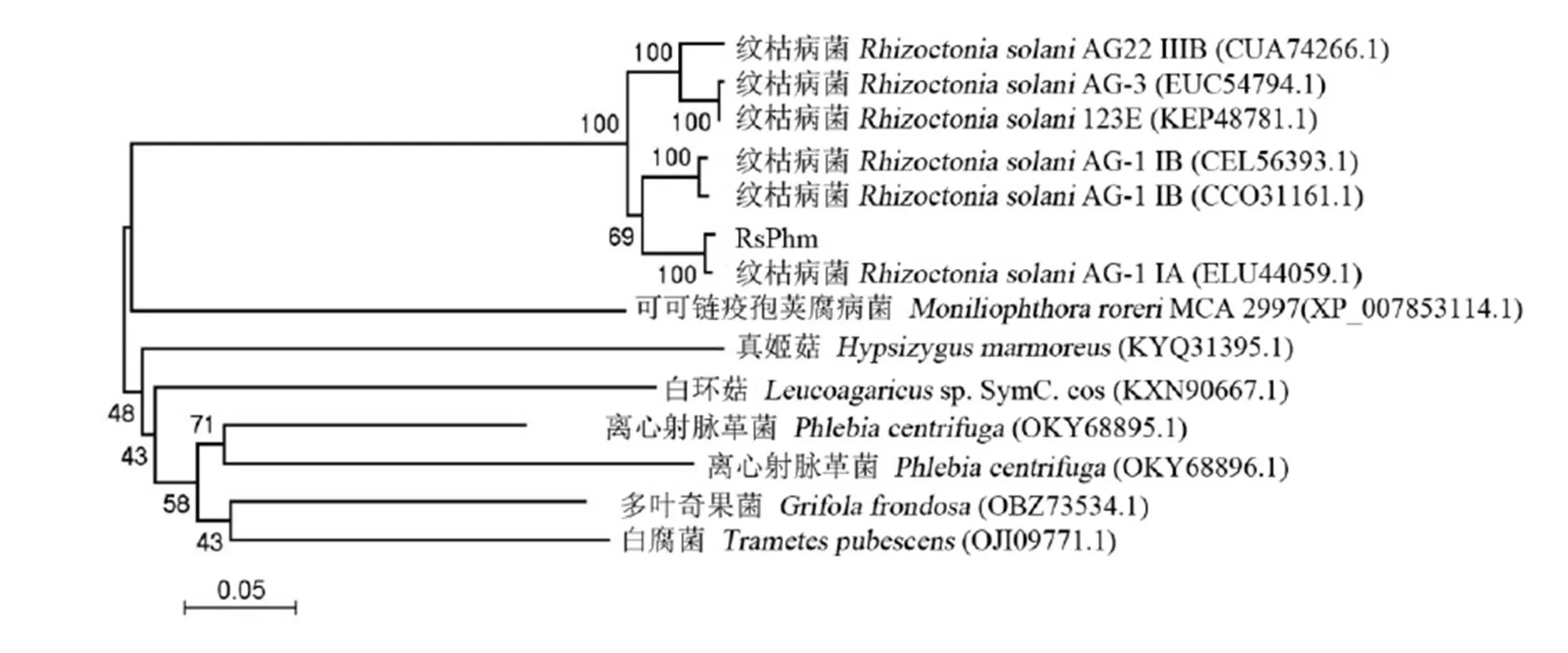
圖4 立枯絲核菌與其他近緣真菌的RsPhm蛋白氨基酸序列的系統進化樹
Fig. 4. Phylogenetic tree of amino acid sequences of RsPhm proteins fromand other related fungi.
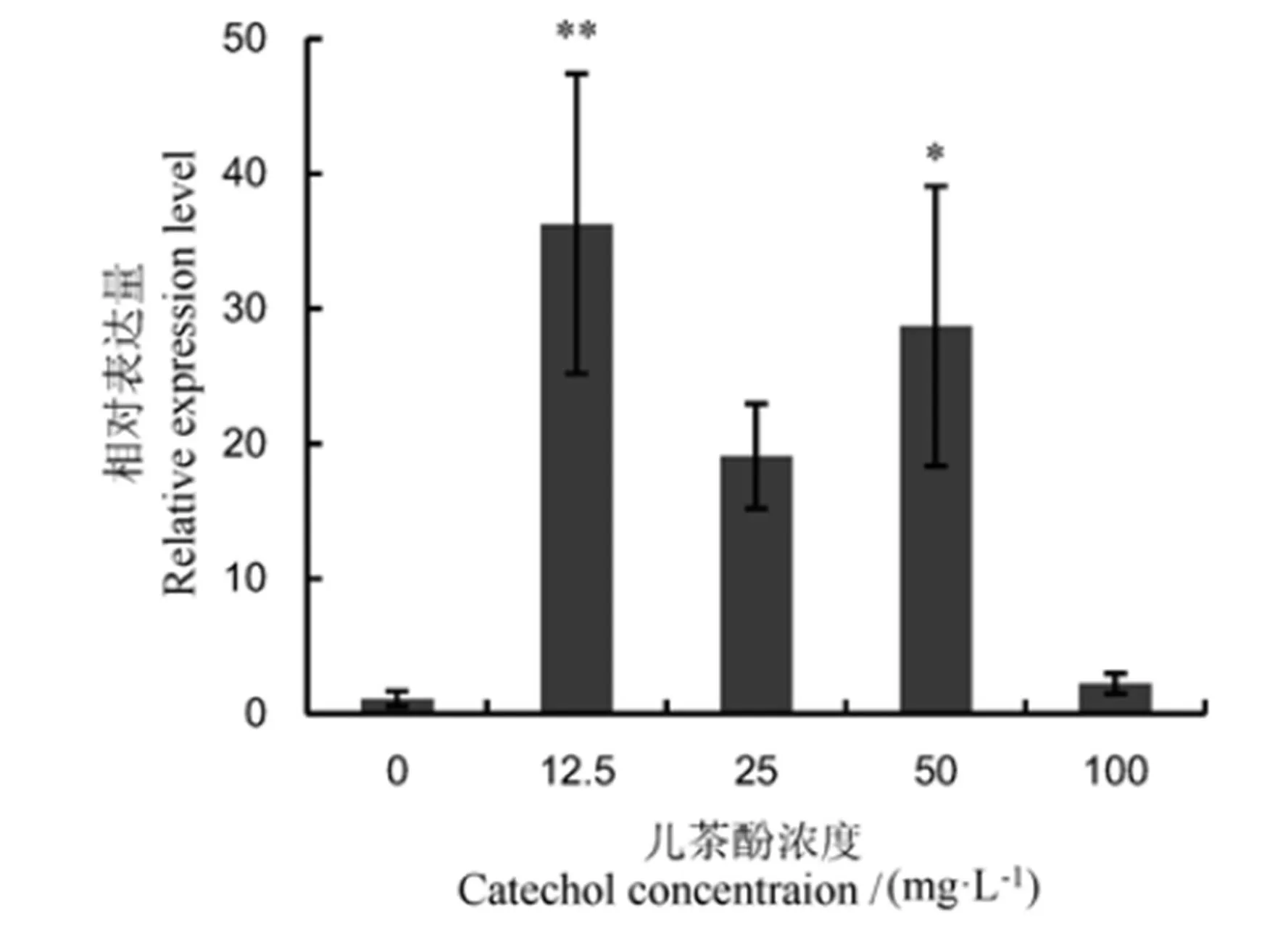
表中數據為3次重復的平均值±標準誤。*和**分別表示相對于對照在5%和1%上差異顯著。
Fig. 5. Expression level ofgeneinAG-1 IA at different concentrations of catechol.
3 討論
大多數的微生物都具有多個單加氧酶和雙加氧酶。苯酚2-單加氧酶屬于加氧酶家族,它是電子傳遞鏈上最關鍵的蛋白,通常含有黃素腺嘌呤二核苷酸(FAD)和鐵硫簇[2Fe-2S],通過輔基負責將電子從NAD(P)H傳遞給羥化酶的雙鐵中心。本研究中RsPhm蛋白具有FAD結構域,跟其他立枯絲核菌的二級結構大致相同(圖2)。在己烷、乙醚和抗壞血酸的存在下,能提高嗜麥芽窄食單胞菌() KB2菌株的苯酚2-單加氧酶的酶活[17]。嗜酸硫化芽孢桿菌()可以將苯酚羥化成低毒的中間物質,因此在降解污染物苯酚中發揮著重要的作用[18]。惡臭假單胞菌菌株DLL-E4(DLL-E4) 的偏三苯酚1, 2-雙加氧酶基因() 能夠對硝基苯酚和對苯二酚進行降解,而敲除菌株DLL-Δ仍然具有利用硝基苯酚和對苯二酚的能力,表明惡臭假單胞菌DLL-E4中除參與PNP降解代謝過程外,還存在另一個雙加氧酶能夠代謝PNP,進而部分恢復了敲除的的功能[19]。惡臭假單胞菌PaW85(PaW85)的包含兒茶酚1,2-雙加氧酶()和苯酚2-單加氧酶()參與苯酚的降解途徑[20]。惡臭假單胞菌()和假單胞菌()都含有兒茶酚1,2-雙加氧酶基因和鄰苯二酚2,3-雙加氧酶基因,能通過多元苯酚羥化酶和鄰位裂解途徑降解長鏈烷基酚[21]。由此可見,單加氧酶和雙加氧酶對微生物代謝有機污染物具有重要作用。水稻紋枯病菌基因在兒茶酚誘導下的表達模式結果表明,隨著兒茶酚濃度升高,基因表達水平提高,同時兒茶酚是RsPhm酶催化的底物,說明基因起到降解兒茶酚的作用。實驗室前期發現兒茶酚和硫脲等抗氧化劑顯著減少了菌核質量和數量[22]。兒茶酚作為菌絲和菌核黑色素形成的前體合成物質,促進黑色素的合成,從而使細胞壁和細胞膜黑化和加厚[14]。這些結果說明基因在水稻紋枯病菌菌核發育和黑色素形成中起到重要作用。
黑色素的合成和形成過程十分復雜。近年對生物體內黑色素合成途徑的研究明確了黑色素起源于4種不同的中間產物:γ-谷氨酰胺酰-3,4-對苯二酚(γ-glutaminyl-3, 4-dihydroxybenzene, GDBH)、多巴(DOPA)、1,8-二羥基萘(DHN)和兒茶酚[23]。核盤菌()能夠不斷產生DHN黑色素,當共同培養核盤菌與白腐菌()時,白腐菌釋放的黑色素降解酶能使核盤菌的黑色素層降解,從而使失去保護層的核盤菌遭受其他溶菌酶的降解,原因是核盤菌黑色素層的存在能幫助其抵御-葡聚糖酶和幾丁質酶的降解[24]。白腐菌產生木質素降解酶,通過過氧化酶代謝途徑可降解黑酵母()黑色素[25]。人類致病真菌新生隱球菌()合成黑色素與毒力密切相關,可以控制黑色素的合成從而降低致病力[26, 27]。立枯絲核菌不同融合群菌絲細胞壁黑色素含量與微生物裂解的敏感性或抗性之間存在相關性,同時也跟致病力密切相關[28, 29]。
本研究結果表明,添加外源兒茶酚可以提高基因的表達量,說明水稻紋枯病菌在兒茶酚脅迫下被誘導表達,但不同的兒茶酚濃度誘導表達量上調倍數不同。因此,水稻紋枯病菌苯酚2-單加氧酶基因表達水平與黑色素形成和菌核發育關系密切。對RsPhm蛋白氨基酸序列的系統進化樹分析表明,立枯絲核菌不同的融合群菌株獨立進化為一支,說明不同的融合群的菌株具有較近的親緣關系,而與離心射脈革菌、多葉奇果菌、白腐菌、白環菇、真姬菇的氨基酸同源性較低,親緣關系的遠近和本身的分類學地位及傳統形態學分類結果一致。因此,不同種屬真菌之間基因在進化上具有一定的保守性。該結果為進一步深入研究水稻紋枯病菌黑色素形成和菌核發育機理奠定了基礎。
[1] Webb K M, Freeman C, Broeckling C D. Metabolome profiling to understand the defense response of sugar beet () toAG 2-2 IIIB., 2016, 94: 108-117.
[2] Hannukkala A O, Rastas M, Laitinen P, Latvala S.injuries in oilseed crops in Finland and impacts of different crop management practices on disease incidence and severity., 2016, 169(2): 257-273.
[3] 鄒成佳, 唐芳, 楊媚, 賀曉霞, 李獻軍, 周而勛. 華南3省(區)水稻紋枯病菌的生物學性狀與致病力分化研究. 中國水稻科學, 2011, 25(2): 206-212.
Zou C J, Tang F, Yang M, He X X, Li X J, Zhou E X. Studies on biological characteristics and pathogenicity differentiation of rice sheath blight pathogen from three provinces in South China.,2011, 25(2): 206-212. (in Chinese with English abstract)
[4] Feng S J, Shu C W, Wang C J Z, Jiang S H, Zhou E X. Survival ofAG-1 IA, the causal agent of rice sheath blight, under different environmental conditions., 2017, 165(1): 44-52.
[5] 劉衛東.DLL-E4 1,2,4 -苯三酚1,2-雙加氧酶的晶體結構及兩種農藥降解相關酶的蛋白質晶體學研究. 南京: 南京農業大學, 2010.
Liu W D. Structural analysis of hydroxyquinol 1, 2-dioxygenase fromDLL-E4 and crystallographic study on two pesticide degradation related enzymes. Nanjing: Nanjing Agricultural University,2010. (in Chinese with English abstract)
[6] Neujahr H Y, Gaal A. Phenol hydroxylase from yeast. Sulfhydryl groups in phenol hydroxylase from., 1975, 58(2): 351-357.
[7] Yotinov I, Todorova Y, Schneider I, Daskalova E, Topalova Y. The effect of nanodiamonds on phenol biodegradation bysp. strain isolated from polluted sediments., 2016, 16(7): 7696-7706.
[8] Nakagawa H, Takeda Y. Phenol hydroxylase., 1962, 62(2): 423-426.
[9] Yang R D, Humphrey A E. Dynamic and steady state studies of phenol degradation in pure and mixed cultures., 1975, 17(8): 1211-1235.
[10] Hamada T, Asanagi M, Satozawa T, Araki N, Banba S, Higashimura N, Akase T, Hirase K. Action mechanism of the novel rice blast fungicide tolprocarb distinct from that of conventional melanin biosynthesis inhibitors., 2014, 39(3): 152-158.
[11] Chai L Y A, Netea M G, Sugui J, Vonk A G, van de Sande W W J, Warris A, Kwon-Chung K J, Jan Kullberg B.conidial melanin modulates host cytokine response., 2010, 215(11): 915-920.
[12] Soares A R, de Lourdes Lucio Ferrarese M, de Cássia Siqueira-Soares R, Marchiosi R, Finger-Teixeira A, Ferrarese-Filho O. The allelochemical 1-dopa increases melanin production and reduces reactive oxygen species in soybean roots., 2011, 37(8): 891-898.
[13] Piattelli M, Fattorusso E, Magno S, Nicolaus R A.melanin, a naturally occurring catechol melanin., 1963, 4(15): 997-998.
[14] Chen J Y, Wang C J Z, Shu C W, Zhu M H, Zhou E X. Isolation and characterization of a melanin from, the causal agent of rice sheath blight., 2015, 142(2): 281-290.
[15] Yang Y Q, Yang M, Li M H, Zhou E X. Cloning and functional analysis of an endo-PG-encoding geneof, the causal agent of rice sheath blight., 2012, 34(3): 1-12.
[16] Zheng A P, Lin R M, Zhang D H, Qin P G, Xu L Z, Ai P, Ding L, Wang Y R, Chen Y, Liu Y, Sun Z G, Feng H T, Liang X X, Fu R G, Tang C Q, Li Q, Zhang J, Xie Z L, Deng Q M, Li S C, Wang S Q, Zhu J, Wang L X, Liu H N, Li P. The evolution and pathogenic mechanisms of the rice sheath blight pathogen., 2013, 4: 1424.
[17] Wojcieszynska D, Gren I, Labuzek S, Respondek M. Substrate specificity and sensitiveness of phenol monooxygenase fromstrain KB2 versus their potential application to bioremediation of the environment., 2007, 2: 181-191.
[18] 李朦. 嗜酸硫化芽孢桿菌苯酚羥化酶還原酶組分的重組表達與性質研究. 廈門: 國家海洋局第三海洋研究所, 2016.
Li M. Expression and characterization of the phenol hydroxylase reductase fromTPY. Xiamen: The Third Institute of Oceanography, State Oceanic Administration, 2016. (in Chinese with English abstract)
[19] 沈文靜, 張靜, 曹慧, 崔中利. 惡臭假單胞菌DLL-E4對硝基苯酚降解途徑關鍵基因的突變分析. 生態與農村環境學報, 2008, 24(4): 77-82.
Shen W J, Zhang J, Cao H, Cui Z L. p-nitrophenol degradation characteristics of hydroxyquinol1, 2-D ioxygenase gene () knock-out mutant ofDLL-E4., 2008, 24(4): 77-82. (in Chinese with English abstract)
[20] Kasak L, H?rak R, Nurk A, Talvik K, Kivisaar M. Regulation of the catechol 1, 2-dioxygenase- and phenol monooxygenase-encodingoperon inPaW85., 1993, 175(24): 8038-8042.
[21] Tuan N N, Hsieh H C, Lin Y W, Huang S L. Analysis of bacterial degradation pathways for long-chain alkylphenols involving phenol hydroxylase, alkylphenol monooxygenase and catechol dioxygenase genes., 2011, 102(5): 4232-4240.
[22] Lu L, Shu C W, Liu C J Z, Zhou E X. The impacts of natural antioxidants on sclerotial differentiation and development inAG-1 IA., 2016, 146(4): 729-740.
[23] Weijn A, Bastiaan-Net S, Wichers H J, Mes J J. Melanin biosynthesis pathway inmushrooms., 2013, 55(6): 42-53.
[24] Butler M J, Gardiner R B, Day A W. Melanin synthesis by, 2009, 101(3): 296.
[25] Butler M J, Day A W. Destruction of fungal melanins by ligninases ofand other white rot fungi., 1998, 159(6): 989-995.
[26] Casadevall A, Rosas A L, Nosanchuk J D. Melanin and virulence in., 2000, 3(4): 354-358.
[27] Eisenman H C, Chow S, Tsé K K, McClelland E, Casadevall A. The effect of L-DOPA ongrowth and gene expression., 2014, 2(4): 329-336.
[28] Hyakumachi M, Yokoyama K, Ui T. Role of melanin in susceptibility and resistance ofto microbial lysis., 1987, 89(1): 27-33.
[29] Kim H T, Chung Y R, Cho K Y. Mycelial melanization ofAG1 affecting pathogenicity in rice., 2001, 17: 210-215.
Cloning and Expression Analysis ofGene inAG-1ⅠA of Rice Sheath Blight Pathogen
JIANG Shaofeng, WANG Chenjiaozi, SHU Canwei, ZHOU Erxun*
(Guangdong Province Key Laboratory of Microbial Signals and Disease Control/College of Agriculture, South China Agricultural University, Guangzhou 510642, China;)
【Objective】In order to elucidate the functions of phenol 2-monooxygenase()gene in melanization ofKühn AG-1ⅠA, the causal agent of rice sheath blight,【Method】the gene was cloned by routine PCR andRT-PCR techniques, and the bioinformatics analysis of this gene was conducted; furthermore, therelative expression levelunder catecholstresswas determinedby using fluorescence quantitative real-time PCR (qRT-PCR) technique.【Result】Bioinformatics analysis showed that the full-lengthDNA and cDNA sequences ofgene were 2628 bp and 1983 bp, respectively, which encode660 amino acids. The phylogenetic tree analysis showed thatgene hada close relationship in different anastomosis groups (AGs) of, and a certain evolutionary conservation among different fungal species. Results of qRT-PCR indicated that the exposure to exogenous catecholcould improvethe expression level ofgene, and which peaked at 12.5 μg/mL of catechol,with a significant increase of 35.7 times, 19.1times and 28.4 timesup-regulated at 25 μg/mL and 50 g/mL, respectively, but only 2.1 times up-regulated at 100 g/mL.【Conclusion】The full-length sequence ofgene was obtained, its basic biological information was understood, and its expression pattern under catechol stress was clarified.Thesefindings will lay a basis for the scientific and systematic elucidation of regulatorymechanism of melanin formation bygene ofAG-1ⅠA.
Kühn AG-1ⅠA;gene; gene clone; expression analysis
Corresponding author, E-mail: exzhou@scau.edu.cn
Q785; S435.111.4+2
A
1001-7216(2018)02-0111-08
2017-08-10;
2017-08-25。
國家自然科學基金資助項目(31271994, 31470247)。
通訊聯系人, E-mail: exzhou@scau.edu.cn
10.16819/j.1001-7216.2018.7094
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
