二倍體與四倍體黃毛草莓的光合特性
2018-04-09 05:47:16徐蘇婷陳露茜李鈞敏
江蘇農業科學 2018年5期
徐蘇婷, 陳露茜, 李鈞敏
(1.上海師范大學生命與環境科學學院,上海 200234; 2.浙江省植物進化生態學與保護重點實驗室/臺州學院生態研究所,浙江臺州 318000)
自1937年Blakeslee和Avery用秋水仙素誘導出多倍體曼陀羅后,全世界掀起多倍體育種熱潮,并獲得了大量的農作物多倍體新品種[1-2]。多倍體植物往往在葉片形態上有更大的葉型、更深的葉色、更厚的葉片、更發達的氣孔和柵欄組織、更豐富的酶物質,其抗病性和抗逆性也往往強于二倍體,品質方面也比二倍體存在更多的優勢,近年來成為研究熱點[3]。
黃毛草莓(FragarainilgerrensisSchlecht.)為薔薇科草莓屬多年生二倍體草本植物,原產于我國西南地區,主要分布于我國陜西、湖北、四川、云南、湖南、貴州和臺灣等省,尼泊爾、錫金、印度東部及越南北部地區也有分布[4]。黃毛草莓花期一般為4—7月,果期為6—8月;果實白色,圓球形,有香味,味淡;種子黃綠色,極小,凹陷;宿存萼片緊貼于果實,具有抗旱、耐高溫、抗葉部病害等特點[5-6]。本實驗室通過秋水仙素誘導成功的四倍體黃毛草莓在形態上與二倍體黃毛草莓相比,具有葉片變大、變寬、葉片增厚、葉色變深等特點,而目前有關二倍體黃毛草莓與四倍體黃毛草莓光合作用的差異未見報道。本研究通過比較分析二倍體與同源四倍體黃毛草莓葉片的光合指標、葉綠素含量等生理指標,了解二者的光合特性差異及環境因子對光合特性的影響,以期為四倍體黃毛草莓的光合生理研究、倍性育種及高效人工種植技術的制定提供理論依據。
1 材料與方法
1.1 材料
二倍體黃毛草莓,2012年7月采自西藏自治區林芝縣色季拉山。2015年春季,將二倍體黃毛草莓種子進行萌發,同時以二倍體黃毛草莓種子經秋水仙素誘變,經流式細胞儀鑒定染色體數量,確定成功獲得四倍體黃毛草莓幼苗;2種幼苗均培養于智能溫室中,相對濕度為80%,溫度白天控制在 20 ℃,晚上控制在15 ℃。
1.2 試驗方法
1.2.1光合特性日變化的測定2016年7月上旬07:00—19:00,選取長勢相近的二倍體與四倍體黃毛草莓各3株,取其冠層陽面健康成熟、生長部位基本相同的功能葉,采用便攜式光合作用測定儀LI-6400分別測定葉片的凈光合速率[Pn,μmol/(m2·s)]、蒸騰速率[Tr,mmol/(m2·s)]、氣孔導度[Gs,mol/(m2·s)]、胞間CO2濃度[Ci,μmol/mol]等參數;每2 h測定1次,每次測定3張樣葉,每張葉片重復記錄5個數據組,取平均值。
1.2.2光響應曲線的測定采用自帶紅藍光光源的LI-6400測定儀,測定光合有效輻射強度(PAR)分別為1 500、1 200、1 000、800、600、400、200、150、100、30、20、0 μmol/(m2·s) 時的凈光合速率;利用直角雙曲線修正模型[7]對光響應曲線進行擬合,計算光飽和點(LSP)、光補償點(LCP)、最大凈光合速率(Pnmax)、表觀量子效率(AQY)、暗呼吸速率(Rd)。
1.2.3相對葉綠素含量的測定取二倍體與四倍體黃毛草莓各3株冠層陽面健康成熟、生長部位基本相同的功能葉,采用SPAD-502葉綠素儀測定相對葉綠素含量,每張葉片從里到外測定3個點,取平均值。
1.3 數據分析
采用Excel 2010、SPSS 18.0軟件對試驗數據進行統計分析,采用Origin 8.5軟件進行輔助作圖,采用單因素方差分析比較二倍體和四倍體黃毛草莓光合指標數據間的差異顯著性。
2 結果與分析
2.1 不同倍性黃毛草莓光合作用的日變化
2.1.1凈光合速率(Pn)由圖1可見,自然光照下,黃毛草莓葉片的Pn日變化呈“雙峰”曲線;07:00時,由于氣溫相對較低,光強較弱,凈光合速率相對較低;隨著氣溫升高、光合輻射增強,11:00時Pn出現第1個峰,此時四倍體黃毛草莓的Pn值為4.15 μmol/(m2·s),而二倍體黃毛草莓為 2.89 μmol/(m2·s);13:00時,Pn出現峰谷,說明黃毛草莓葉片存在光合“午休”現象;后隨光合有效輻射、葉片表面溫度的下降,17:00時Pn出現第2個峰,此時四倍體黃毛草莓的Pn值為1.86 μmol/(m2·s),而二倍體黃毛草莓為 1.22 μmol/(m2·s)。

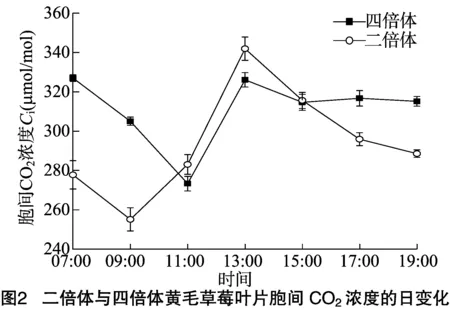
2.1.2胞間CO2濃度(Ci)由圖2可見,Ci日變化與Pn日變化的趨勢基本相反;07:00時,黃毛草莓的Ci相對較高,四倍體黃毛草莓的Ci值為327.04 μmol/mol,二倍體黃毛草莓為277.78 μmo/mol;11:00時,四倍體黃毛草莓Ci出現最低值,后逐漸增大;13:00時,四倍體黃毛草莓的Ci值上升為326.10 μmol/mol,二倍體黃毛草莓的Ci值上升幅度相對更大,為 341.92 μmol/mol;后逐漸下降。
2.1.3氣孔導度(Gs)由圖3可見,黃毛草莓氣孔導度自09:00時出現谷值后,11:00時出現峰值,此時四倍體黃毛草莓Gs值為0.088 mol/(m2·s),二倍體黃毛草莓為 0.068 mol/(m2·s);后逐漸降低。
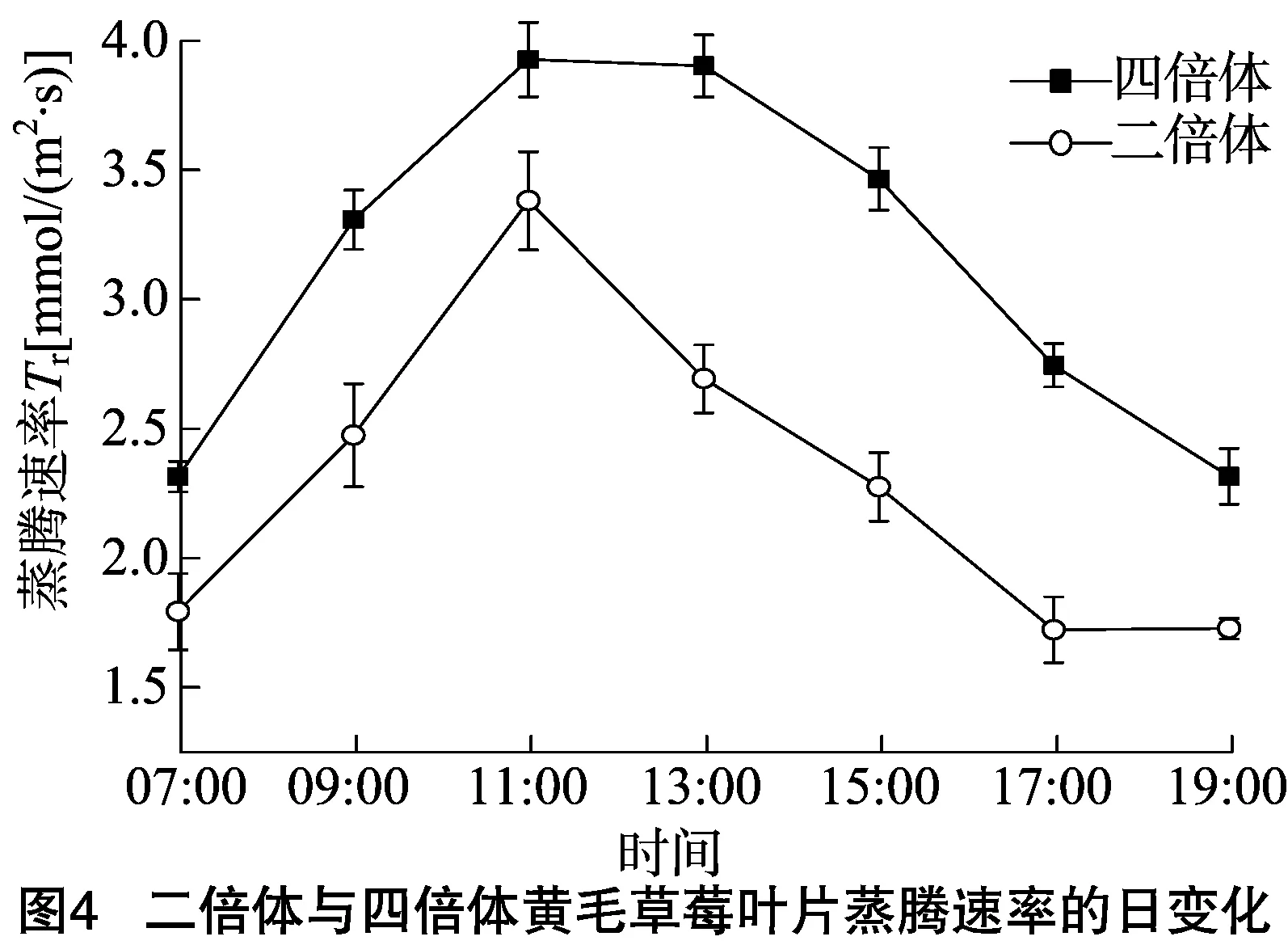
2.1.4蒸騰速率(Tr)由圖4可見,黃毛草莓的Tr日變化呈“單峰”曲線;Tr值在11:00正午時達到峰值,四倍體黃毛草莓的Tr值為3.92 mmol/(m2·s),二倍體黃毛草莓為 3.38 mmol/(m2·s);后逐漸降低。

2.2 四倍體與二倍體黃毛草莓的光響應曲線
圖5為2種不同倍性黃毛草莓的光響應曲線,對其光響應曲線進行擬合發現,二倍體黃毛草莓的最大凈光合速率、光飽和點、光補償點、暗呼吸速率分別為3.451、885.6、23.83、0.527 μmol/(m2·s),表觀量子效率為0.024,四倍體黃毛草莓的最大凈光合速率、光飽和點、光補償點、暗呼吸速率分別為4.223、1 663.6、41.87、1.601 μmol/(m2·s),表觀量子效率為0.051,其中四倍體黃毛草莓的光響應曲線中光飽和點、光補償點、暗呼吸速率這3個指標極顯著大于二倍體黃毛草莓(P<0.01)。

2.3 相對葉綠素含量
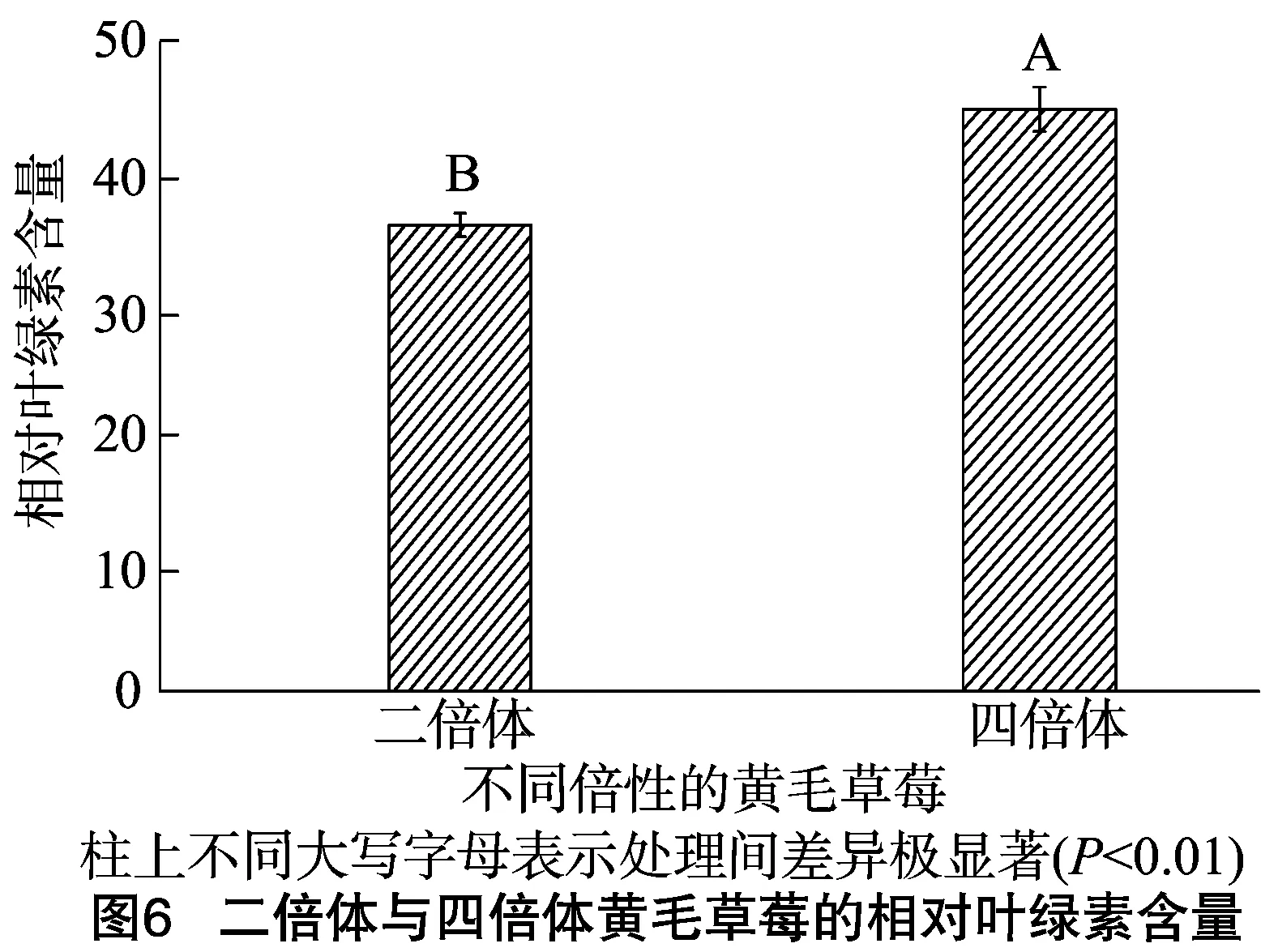
由圖6可見,四倍體黃毛草莓的相對葉綠素含量值極顯著高于二倍體黃毛草莓(P<0.01)。
3 結論與討論
光合作用是植物最基本的生命活動,是植物合成有機物質、獲得能量的根本源泉[8]。通過研究發現,四倍體黃毛草莓的凈光合速率(Pn)、氣孔導度(Gs)、蒸騰速率(Tr)均高于二倍體黃毛草莓,1天中四倍體黃毛草莓的最大凈光合速率、氣孔導度、蒸騰速率分別為4.15 μmol/(m2·s)、0.088 mol/(m2·s)、3.92 mmol/(m2·s),分別比二倍體黃毛草莓高43.6%、29.4%、16.0%,與鄭思鄉等的研究結論[9-10]一致;夏季黃毛草莓葉片的Pn日變化呈“雙峰”曲線,并出現光合“午休”現象;四倍體和二倍體黃毛草莓的Tr、Gs規律基本相似,而胞間CO2濃度(Ci)的日變化與Pn相反;中午出現“午休”現象時Ci和Gs同時升高,說明該現象的發生可能是由葉肉阻力、葉肉自身羧化酶活性發生變化等非氣孔限制因素造成。
光飽和點(LSP)、光補償點(LCP)、表觀量子效率(AQY)及暗呼吸速率(Rd)是植物光響應的重要指標,能夠反映植物葉片對弱光和強光的適應能力,有助于判定植物光合作用能力、光合作用效率、光合機構的運轉狀況及植物受到環境變化的影響程度[11-14]。光飽和點與光補償點分別代表植物可利用光合有效輻射的上限與下限,體現植物對強光和弱光的利用能力和對光照條件的要求[15]。本研究發現,隨著光強的增加,四倍體和二倍體黃毛草莓的凈光合速率均明顯增大,四倍體黃毛草莓的LSP和LCP明顯高于二倍體黃毛草莓,說明同種條件下四倍體黃毛草莓對光環境的適應性較二倍體黃毛草莓強,對光照的適應性也高于二倍體黃毛草莓。另外,四倍體黃毛草莓的Pn、Rd值較二倍體黃毛草莓高,光合產物多、呼吸消耗大,這可保證四倍體黃毛草莓葉片形態上葉型更大、葉色更深、葉片更厚,在生長過程中有更強的活力。
相對葉綠素含量(SPAD)高低直接影響葉片的光合能力,葉綠素含量和組成與光合速率有著密切的聯系[16],而SPAD值與葉片葉綠素含量呈正相關關系,能較好地反應植物葉片葉綠素變化,其值越高,越有利于植物捕獲更多光能用于光合作用[17]。本研究發現,四倍體黃毛草莓的相對葉綠素含量遠遠大于二倍體黃毛草莓,且差異極顯著,這說明四倍體黃毛草莓能夠捕獲更多的光能用于光合作用。
總之,同等條件下四倍體黃毛草莓的光飽和點、光補償點、凈光合速率、氣孔導度、蒸騰速率及相對葉綠素含量均大于二倍體黃毛草莓,四倍體黃毛草莓在栽培過程中對光環境的適應性更強,相應的水分需求更多,更適宜高濕栽培環境。
參考文獻:
[1]康向陽. 林木多倍體育種研究進展[J]. 北京林業大學學報,2003,25(4):70-74.
[2]馬海淵,張金鳳,李志丹. 植物多倍體育種技術方法研究進展[J]. 防護林科技,2008(1):43-46.
[3]于文艷,劉世琦,梁慶玲,等. 大蒜二倍體與四倍體生長和生理指標的比較研究[J]. 山東農業科學,2008(2):42-44,65.
[4]俞德浚,陸玲娣,谷粹芝. 中國植物志[M]. 北京:科學出版社,1985.
[5]晁無疾,鐘新. 秦巴山區野生草莓資源及其研究[J]. 作物品種資源,1988(4):3-5.
[6]雷家軍,楊高,代漢平,等. 我國的草莓野生資源[J]. 果樹科學,1997,14(3):198-200.
[7]葉子飄. 光合作用對光和CO2響應模型的研究進展[J]. 植物生態學報,2010,34(6):727-740.
[8]趙依杰,張小紅,林航,等. 二倍體與四倍體西瓜葉片若干生理指標比較[J]. 中國農學通報,2010,26(16):144-146.
[9]鄭思鄉,胡秀,雷小云,等. 離體培養條件下三色堇多倍體誘導研究[J]. 云南農業大學學報,2003,18(4):397-400.
[10]張杰,張蜀寧,徐偉鈺,等. 二、四倍體青花菜凈光合速率日變化及其影響因子的相關和通徑分析[J]. 江蘇農業科學,2006(6):220-223.
[11]Zeng X M,Yuan L,Shen Y G. Response of photosynthesis to light intensity in intact and detached leaves ofArabidopsisthaliana[J]. Plant Physiology Communications,2002,38(1):25-26.
[13]許殊. 重慶市常見園林植物光合和生理生態特性[J]. 江蘇農業科學,2016,44(8):281-285.
[14]高傳友. 廣西地區15種典型園林觀賞植物的耐陰性及光合特性[J]. 江蘇農業科學,2016,44(4):209-214.
[12]Ye Z P,Yu Q. Comparison of new and several classical models of photosynthesis in response to irradiance[J]. Chinese Journal of Plant Ecology,2008,32(6):1356-1361.
[15柯世省,金則新,林恒琴,等. 天臺山東南石櫟光合生理生態特性[J]. 生態學雜志,2004,23(3):1-5.
[16]尤鑫,龔吉蕊. 葉綠素熒光動力學參數的意義及實例辨析[J]. 西部林業科學,2012,41(5):90-94.
[17]俞世雄,李芬,李紹林,等. 水分脅迫對小麥新品系葉綠素含量的影響[J]. 云南農業大學學報(自然科學版),2014,29(3):353-358.
