外源赤霉素對鹽脅迫下水稻種子萌發及幼苗生長的緩釋效應
2018-04-18 12:37:15張麗麗倪善君張戰趙一洲李鑫毛艇劉研劉福才
中國稻米 2018年2期
關鍵詞:水稻
張麗麗 倪善君 張戰 趙一洲 李鑫 毛艇 劉研 劉福才
(遼寧省鹽堿地利用研究所,遼寧盤錦124010)
土壤鹽漬化是影響土地生產力的重要障礙因子,嚴重制約了糧食生產和農業的可持續發展。據估計,至2050年將有過半的耕地鹽漬化[1]。水稻是我國第一大糧食作物,鹽漬土占水稻栽培總面積的約1/5,且有日益擴大和蔓延的趨勢。因此,如何在當前人口不斷增加、耕地面積日趨減少及水資源日益匱乏的情況下,合理開發、有效利用鹽漬土資源,培育耐鹽作物品種或者施加一些外源物質等措施緩解土壤鹽漬化程度,增強作物耐鹽性,己成為國內外科研工作者迫切解決的重大課題。
植物激素是植物自身產生的調節物質,是植物脅迫響應的信號轉導的主要成員,參與了植物對鹽漬生境的適應性調節過程。研究激素調控植物對環境適應的機制對提高作物耐鹽性具有重要的現實意義[2]。赤霉素(GA)是一個較大的萜類化合物家族,在植物整個生命循環過程中起著重要的調控作用。前人關于GA3在植物伸長生長、細胞活性及蛋白質表達等方面作用做了大量研究,并取得了一些進展[3-5]。本文旨在研究外源赤霉素對鹽脅迫下不同耐鹽性水稻種子萌發及幼苗生長的緩釋作用,探討耐鹽性不同水稻對外源赤霉素響應的差異,以期為進一步揭示激素緩釋機理提供理論依據。
1 材料與方法
1.1 試驗材料
本研究選用耐鹽品種長白9號和鹽敏感品種越光為試驗材料。試驗于2014年進行。
1.2 種子萌發試驗
隨機挑選飽滿的種子,經3%H2O2消毒15 min,并用蒸餾水沖洗干凈,再用赤霉素浸種24 h,赤霉素濃度分別為0、25 mg/L、50 mg/L和100 mg/L 3個梯度。先在直徑為9 cm的培養皿中放2層濾紙并進行編號(Ⅰ-Ⅴ)除Ⅰ號培養皿中加入20 mL蒸餾水外其余4個加入20 mL 100 mM NaCl處理液,然后在每個培養皿中擺放40粒浸種24 h后的水稻種子,每個處理設3個重復,然后把所有處理都放在28℃恒溫培養箱中發芽,每天換1次處理液。
1.3 幼苗培養和處理
前期試驗結果表明,在100 mM NaCl脅迫下,供試材料幼苗生長受到顯著抑制,可以作為研究赤霉素緩解鹽害的脅迫濃度。
種子經消毒后,用去離子水洗凈,25℃浸種48 h,后28℃條件下催芽24 h,然后置于人工氣候箱中沙培,1周后用1/2木村營養液[6]水培,溫度25℃,光/暗時間為14 h/10 h。培養至1葉1心期轉移至裝有1/2木村B營養液的塑料箱中,1周后換成全木村B營養液。幼苗用海綿固定在有孔的苯板上,外部用遮光紙遮光,培養室光/暗溫度28℃/20℃,光/暗時間為14 h/10 h。培養至3葉1心時,加入NaCl并噴施赤霉素物質進行處理。每種材料設5個處理,分別為對照(CK,0 mM Na-Cl+0 mg/L GA3)、G0(100 mM NaCl+0 mg/L GA3)、G1(100 mM NaCl+25 mg/L GA3)、G2(100 mM NaCl+50 mg/L GA3)和G3(100 mM NaCl+100 mg/L GA3),脅迫7 d后取樣進行各指標測定。
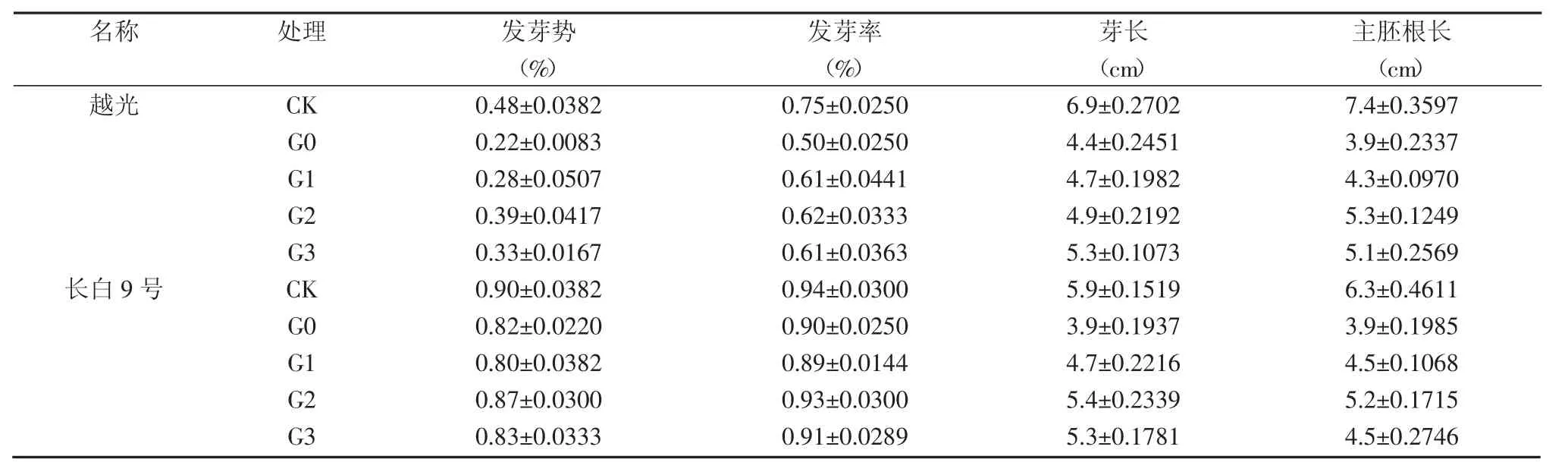
表1 赤霉素對NaCl脅迫下種子發芽的緩解效應
1.4 測定指標及方法
1.4.1 發芽率、發芽勢、芽長和主胚根長
以芽長超過1/2種子長度且根長超過種子長度為發芽標準,每天統計發芽數,連續統計7 d。第4 d和第7 d記錄發芽數,計算發芽率、發芽勢。發芽率(%)=(發芽終期全部正常發芽的種子數/供試種子數)×100,發芽勢(%)=(發芽前4 d全部正常發芽的種子數/供試種子數)×100。10 d后隨機取10粒種子測定主胚根長和芽長。
1.4.2 葉綠素含量
參照張憲政[7]的方法測定葉綠素含量(包括葉綠素總量、葉綠素a和葉綠素b)。
1.4.3 膜脂過氧化
參照張憲政[7]的方法測定葉片丙二醛含量、相對電導率和O2-含量。
1.4.4 數據分析
采用DPS軟件和Excel軟件進行統計分析。
2 結果與分析
2.1 外源赤霉素對鹽脅迫下水稻種子發芽的影響
從表1可見,鹽脅迫使供試品種種子發芽率降低,不同品種間受影響程度不同。越光在G0處理下發芽率與CK相比降低了33.3%,長白9號僅降低了3.6%。外源赤霉素處理提高了鹽脅迫下參試品種的發芽率。越光發芽率在G1、G2和G3處理下均高于G0處理;赤霉素對鹽脅迫下長白9號發芽率的影響不大,G2處理略高。
鹽脅迫對耐鹽性不同品種種子發芽勢的抑制程度有所不同。在G0處理下,越光和長白9號發芽勢均低于CK,分別比CK降低了54.4%和8.9%。赤霉素處理能緩解鹽脅迫對參試品種發芽勢的抑制程度,特別是對鹽敏感品種越光的緩解作用更明顯,與G0處理相比,G1、G2和G3處理發芽勢分別提高了30.7%、80.8%和53.8%。赤霉素對鹽脅迫下長白9號的種子發芽略有緩解作用。
鹽脅迫下,水稻種子芽長和主胚根長生長受到明顯抑制。赤霉素對鹽脅迫下芽長和主胚根長生長存在一定促進作用。G1、G2和G3處理與G0處理相比,長白9號和越光的芽長和主胚根長均有不同程度的提高,且G2處理的效果要好于其他處理。表明外源赤霉素能有效緩解鹽脅迫對幼芽和根長的抑制作用。
2.2 外源赤霉素對鹽脅迫下水稻幼苗葉片葉綠素含量的影響
由圖1所示,參試品種在鹽脅迫下葉綠素總量、葉綠素a含量和葉綠素b含量均不同程度降低。葉面噴施赤霉素后,各參試品種的葉綠素總量和葉綠素a含量不同程度增加,而葉綠素b含量呈降低趨勢。
長白9號葉綠素總量G0處理與CK相比降低了16.4%,G1、G2和G3處理與CK相比分別降低了3.9%、15.7%和11.1%;長白9號葉綠素a含量G0處理與CK相比降低了17.8%,G1、G2和G3處理與CK相比分別降低了6.0%、10.6%和8.9%;葉綠素b含量G0處理與CK相比降低了14.9%,G1、G2和G3處理與CK相比分別降低了21.0%、30.9%和20.4%,以G2處理下降最明顯。
越光葉綠素總含量G0處理與CK相比降低了18.3%,G1、G2和G3處理與CK相比分別降低了8.4%、12.4%和23.3%,G1、G2處理與G0處理間差異達極顯著(P<0.01)。葉綠素a含量變化趨勢與葉綠素總量基本一致,G0處理與CK相比降低了18.1%,而G1、G2、G3與CK相比分別降低了0.3%、0.5%和14.5%,G1、G2處理與G0處理間差異達極顯著(P<0.01)。越光葉綠素b含量G0處理與CK相比降低了18.5%,G1、G2和G3處理與CK相比分別降低了28.5%、30.1%和47.4%,以G3處理下降最明顯。
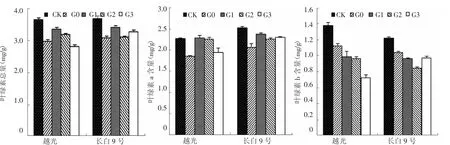
圖1 外源赤霉素對鹽脅迫下水稻葉片葉綠素含量的緩解效應
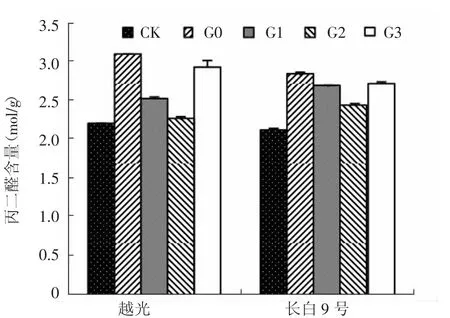
圖2 赤霉素對鹽脅迫下水稻幼苗葉片丙二醛含量的影響
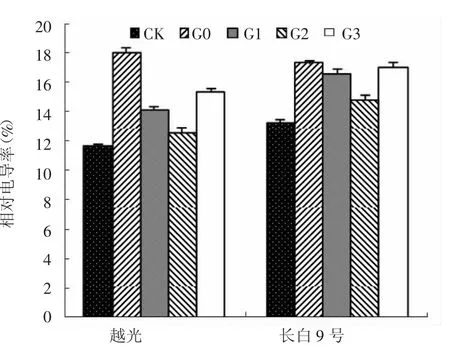
圖3 赤霉素對鹽脅迫下水稻幼苗葉片相對電導率的影響
2.3 外源赤霉素對鹽脅迫下水稻幼苗葉片膜脂過氧化的影響
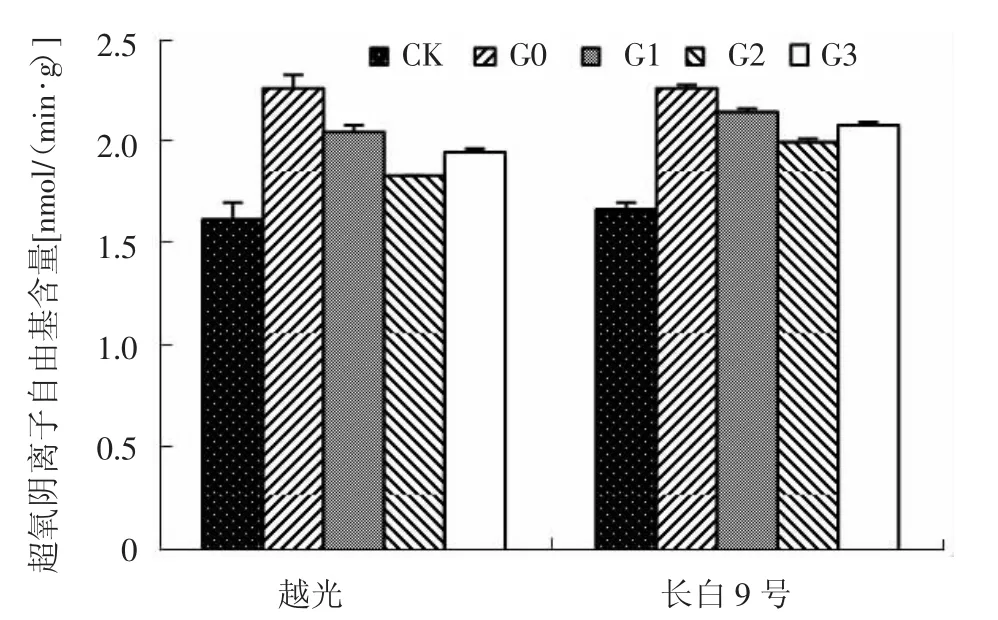
圖4 赤霉素對鹽脅迫下水稻幼苗葉片超氧陰離子自由基含量的影響
丙二醛(MDA)是膜脂過氧化作用的主要產物之一,其含量是反映細胞膜脂過氧化作用強弱的重要指標。如圖2所示,G0處理使耐鹽性不同的2個水稻品種葉片MDA含量均極顯著增加,分別比CK升高41.5%和33.6%。葉面噴施赤霉素后,長白9號和越光葉片MDA含量明顯降低,且二者變化趨勢一致。與G0處理相比,G1、G2和G3處理長白9號葉片MDA含量分別降低了5.2%、14.2%和4.2%,越光則分別降低了18.3%、27.2%和5.4%。表明鹽脅迫下噴施赤霉素具有降低膜脂過氧化作用,減輕了對植物細胞的傷害。
如圖3所示,G0處理下,2個水稻品種葉片相對電導率與CK相比均極顯著增加,分別比CK升高了55.2%和30.6%。外源赤霉素處理可明顯降低葉片相對電導率。與G0處理相比,G1、G2和G3處理長白9號相對電導率分別降低了4.2%、14.3%和1.7%,G2處理與G0處理之間差異極顯著(P<0.01)。越光葉片相對電導率變化趨勢與長白9號基本一致,與G0處理相比,G1、G2和G3處理分別降低了21.9%、30.2%和15.0%。
如圖4所示,未經鹽脅迫處理,參試品種葉片中O2-的產生速率較低;G0處理導致2個水稻品種葉片O2-的產生速率均極顯著增加,分別比CK升高39.4%和35.9%。噴施外源赤霉素后參試品種葉片中O2-的產生速率顯著降低。與G0處理相比,G1、G2和G3處理長白9號葉片中O2-的產生速率分別降低了5.0%、11.4%和7.8%,越光分別降低了9.2%、19.1%和13.7%。表明G2處理效果要好于G1、G3處理,越光降低幅度大于長白9號。
3 討論
赤霉素是指具有赤霉烷骨架,并能刺激細胞分裂和伸長的一類化合物的總稱,其最顯著的生理效應就是促進植物的生長,主要是促進細胞伸長[8]。一定濃度赤霉素可以促進多種作物種子萌發和幼苗生長[9-10]。前人研究結果表明,10 μmol/L GA3處理能顯著緩解一定濃度范圍內的鹽脅迫對水稻種子發芽及幼苗生長的抑制作用[11]。本研究結果表明,本試驗中不同濃度赤霉素均提高了鹽脅迫下水稻種子發芽率,并可以顯著地恢復鹽脅迫下種苗根和芽的伸長生長。其中以G2處理效果最佳。這可能是鹽脅迫使植物的內源赤霉素活性下降,導致植物生長受抑制,而外源赤霉素可以部分抵消這種影響,進而提高水稻耐鹽性,這與前人研究結果一致[2]。也有研究認為,這是由于赤霉素能促進細胞對Ca2+的吸收,提高細胞質中的Ca2+濃度,并活化ER上的Ca2+-ATPase活性,促進Ca2+向ER中運輸,提高ER活性,從而促進種子萌發[12]。
葉綠體是植物細胞中對鹽分最敏感的細胞器,易受到鹽脅迫的傷害。當植物遭受鹽脅迫時,葉綠體在葉肉細胞中的排列呈現紊亂,基粒間連接松馳,類囊體內腔膨大,葉綠體雙層被膜部分出現損壞,脂質小球增多[13]。本研究結果表明,鹽脅迫下耐鹽性不同的2個水稻品種葉綠素含量均顯著降低,噴施25~100 mg/L外源赤霉素均可以提高鹽脅迫下水稻葉片中葉綠素總量和葉綠素a含量,其中以50 mg/L效果最為明顯。表明外源赤霉素可以有效地抑制水稻葉片中葉綠素降解,這與徐鍇等[14]研究結果一致。本試驗中鹽脅迫下外施赤霉素導致葉片中葉綠素b含量繼續下降,其原因有待進一步明確。
逆境脅迫可誘發植物細胞內產生過量的活性氧,活性氧積累會造成膜、蛋白質和DNA分子結構等損傷[15],活性氧代謝失調是逆境下需氧生物受害的普遍表現,也是逆境損傷的重要原因之一[16],過多的活性氧還可導致膜脂過氧化,并進一步引起DNA斷裂,導致基因突變[17],可以引起膜透性增大,外滲液中離子濃度升高,相對電導率增大,相對電導率是標志膜受損傷即植物受脅迫傷害程度的指標[18],而MDA是活性氧啟動膜脂氧化過程中產生的主要產物之一,其含量是衡量植物在逆境脅迫下活性氧傷害程度大小的常用指標[19]。本試驗結果表明,在鹽脅迫條件下耐鹽性不同的2個品種幼苗葉片的MDA、超氧陰離子自由基含量和相對電導率均顯著增加,而噴施25~100 mg/L外源赤霉素后,葉片MDA含量、超氧陰離子自由基含量和相對電導率均明顯下降,其中以50 mg/L的處理效果最為明顯。由此可見,外源赤霉素可以緩解鹽脅迫對水稻植株的損害,降低葉片的膜脂過氧化作用,減少活性氧的積累和產生,從而提高植物耐鹽能力,這可能是外源赤霉素處理提高水稻耐鹽性的原因之一。
綜上所述,外源赤霉素在一定濃度范圍內能有效緩解鹽脅迫對水稻種子萌發和幼苗生長的傷害,其中以50 mg/L的濃度緩解效果最為明顯。而耐鹽性不同水稻品種長白9號與越光對赤霉素響應不同,緩解機制也不盡相同,有待進一步明確。
[1] Ashraf M.Breeding for salinity tolerance in plants[J].Crit Rev Plant Sci,1994,13:17-42.
[2] 溫福平,張檀,張超暉,等.赤霉素對鹽脅迫抑制水稻種子萌發的緩解作用的蛋白質組分析[J].作物學報,2009,35(3):483-489.
[3] Konishi H,Yamane H,Maeshima M,et al.Characterization of fructose-bisphosphate aldolase regulated by gibberellin in roots of rice seedling[J].Plant Mol Biol,2004,56:839-848.
[4] Komatsu S,Konishi H.Proteome analysis of rice root proteins regulated by gibberellin[J].Genomics Proteomics Bioinformatics,2005,3(3):132-142.
[5] Komatsu S,Zang X,Tanaka N.Comparison of two proteomics techniques used to identify proteins regulated by gibberellin in rice[J].J Proteome Res,2006,5(2):270-276.
[6] 許明學,樸昌浩,金瀅完.水稻在不同營養液中生育情況的研究[J].延邊農學院學報,1984,5(1):14-20.
[7] 張憲政.作物生理研究法[M].北京:農業出版社,1990.
[8] 蔡永萍.植物生理學[M].北京:中國農業大學出版社,2008:117-181.
[9] 劉曉冰,曾寒冰.植物生長調節物質對春小麥種子萌發的效應[J].東北農學院學報,1991,22(3):220-224.
[10] Jhorar B S.Effect of gibberellic acid on seedling emergence and early growth of cotton Indian[J].J Plant Physiol,1982,25:423-426.
[11] 尹昌喜,汪獻芳,金莉,等.赤霉素對鹽脅迫下水稻種子發芽及幼苗生長的影響[J].安徽農業科學,2009,37(14):6 389-6 390.
[12] 侯振安,李品芳,龔元石.激素對植物耐鹽性影響的研究現狀與展望[J].石河子大學學報,2000,4(3):239-245.
[13] Barhoumi Z,Djebali W,Abdelly C,et al.Salt impact on photosynthesis and leaf ultrastructure of Aeluropus littoralis[J].J Plant Res,2007,120(4):529-537.
[14] 徐鍇,金芳.外源激素對鹽脅迫下草莓光合性能的影響[J].甘肅農業大學學報,2006,41(2):31-34.
[15] 梁慧敏,夏陽,王太明.植物抗寒凍、抗旱、耐鹽基因工程研究進展[J].草業學報,2003,12(3):1-7.
[16] 陳潔,林棲鳳.植物耐鹽生理及耐鹽機理研究進展[J].海南大學學報:自然科學版,2003,21(2):177-188.
[17] 劉曉麒,曹恩華.脂質過氧化引起DNA損傷研究進展[J].生物化學與生物物理進展,1994,21(3):218-222.
[18] 閻秀峰,孫國榮,那守海,等.鹽脅迫下星星草幼苗的生理Ⅱ--鹽脅迫對星星草幼苗膜透性的影響[J].黑龍江畜牧獸醫,1994(4):1-3.
[19] 李合生.植物生理生化試驗原理和技術[M].北京:高等教育出版社,2000.
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
