多效唑和干旱脅迫對毛竹實生苗活力、光合能力及非結構性碳水化合物的影響
2018-04-19 06:34:38楊麗芝潘春霞邵珊璐陶晨悅應葉青
生態學報 2018年6期
關鍵詞:植物
楊麗芝,潘春霞,邵珊璐,陶晨悅,王 威,應葉青,*
1 浙江農林大學省部共建亞熱帶森林培育國家重點實驗室, 臨安 311300 2 安吉縣林業技術推廣中心, 湖州 313300
隨著全球氣候變暖,世界范圍內的干旱問題日益嚴重。植物因蒸騰作用導致失水量多于根系的吸水量時會產生干旱脅迫,引起植物生理生化過程的變化。目前干旱已經成為影響植物健康生長的主要逆境因子之一[1- 4]。根系是植物與土壤直接密切接觸的重要器官,最先感知干旱脅迫,并迅速產生化學信號向上傳遞,促使葉片氣孔關閉,以降低蒸騰作用,減少水分流失[5],其與地上部植株的生長及植株最終產量形成密切相關[6],已成為衡量植物抗旱能力的指標[7]。干旱脅迫使葉綠體形態結構發生改變,葉綠素含量下降[8],植物的光合作用被顯著抑制,光合速率降低,影響光合電子傳遞等,進而影響碳同化[9]。有研究發現,植物的抗旱性與非結構性碳水化合物含量(NSC)有密切關系,NSC作為糖源為植物的生長發育提供能量,反應植物的碳供應狀況,還能通過滲透調節作用平衡細胞的滲透勢[10],從而調節植物生理來適應環境變化[11- 13]。
多效唑(paclobutrazol;PP333),不僅具有調節植物生長和抑菌的雙重作用,又具有廣譜、低毒,對人畜安全、施用方便等特點[14]。近十幾年來,國內外對多效唑在果樹、蔬菜、花卉、作物等植物上的調節作用[15],主要表現為:增強根系活力,促進葉綠素合成[16],增強植物抗逆性[17- 18],影響植物的光合作用、呼吸作用、礦質營養代謝、碳水化合物的的代謝與分配,延緩植物衰老等生理效應[19- 20]。王競紅等[21]研究認為,對黑麥草施加多效唑能明顯提高草坪草的干旱適應性;Stephen發現高溫和干旱雙重脅迫下,PP333能顯著提高匍匐剪股穎(Agrostisstolonifea)的葉片相對含水量和土壤含水量,降低匍匐剪股穎在脅迫下的蒸騰作用,在草地早熟禾上的研究得到了類似的結論[22- 23];在應對惡劣的生存環境時,狗牙根除了調整自身生存策略外,還可人為施加PP333來提高對逆境的適應能力[24]。
毛竹是禾本科(Gramineae)竹亞科(Bambusoideae)剛竹屬植物,是我國最重要的生態經濟竹種[25],主要分布在南方山地地區。近年來,毛竹主產區夏季持續高溫導致季節性干旱頻發,毛竹的正常生長受到嚴重威脅。多效唑作為一種植物生長調節劑能否提高毛竹抗旱性未見報道,其相關作用機制也尚不明確。本試驗通過不同程度干旱脅迫條件下施加不同濃度的多效唑,研究毛竹實生苗光合特性及非結構性碳水化合物對干旱和多效唑及其互作的響應,以期為季節性干旱頻發的背景下如何提高毛竹抗旱能力、科學管理毛竹林地提供理論依據。
1 材料與方法
1.1 試驗材料
2016年1月6日將毛竹種子(于2015年10月份,從廣西桂林生長健壯的單株毛竹上采集)播種于培養缽中,每盆播10顆種子,出苗率大約40%—50%,最終每盆保留4株實生苗。盆規格為上口直徑15.5cm、下口直徑11cm、高13.0cm。基質按泥炭∶珍珠巖∶蛭石按1∶1∶1比例混合,pH值在5.5—6.0之間,并混入復合肥為底肥,每20盆為一組。置于浙江農林大學智能實驗樓的栽培室中進行培育(30°23′N,119°72′E),日溫/夜溫為(23±2)℃/(17±2)℃,濕度為45%—70%,進行常規的水肥管理。待毛竹生長健壯穩定后,于2016年7月9日選取生長一致(實生苗平均株高達到15cm)的盆栽實生苗進行試驗。試驗中所用的PP333(純度99%),由上海生物工程有限公司生產,該處理溶液均現配現用。
1.2 試驗設計
試驗采用隨機區組設計,試驗設2個多效唑濃度,分別為0mg/L(P1)、40mg/L(P2);設3個水分梯度,分別為75%相對田間持水量(W1)、50%相對田間持水量(W2)、35%相對田間持水量(W3),共6個處理,分別記作P1W1(0mg/L,75%)、P1W2(0mg/L,50%)、P1W3(0mg/L,35%)、P2W1(40mg/L,75%)、P2W2(40mg/L,50%)、P2W3(40mg/L,35%),其中P1W1為對照組。盆栽毛竹幼苗于7月9日到7月12日每天下午17:00,分別用100mL濃度為40mg/L多效唑根施處理,對照加等量的水,每處理設5個重復,每重復4盆竹苗,共計120盆竹苗。通過稱重法每天確定土壤含水量,7月19日盆栽苗基質含水量達到中度干旱(土壤含水量占相對田間持水量45%—50%),7月24日盆栽苗基質含水量達到重度干旱(土壤含水量占相對田間持水量30%—35%),采用稱重法控制土壤水分,保持盆栽苗基質含水量在中度和重度干旱下,于每天的18:00對整盆竹苗進行稱重并補充水分使之達到相應的處理標準。于8月14日測定毛竹幼苗葉片光合參數,8月15日7:00采集從上往下第2—3節位成熟葉片,測定葉綠素含量,根葉里的可溶性總糖、淀粉、氮、蔗糖、果糖含量等生理生化指標。9月15號8:00,每個處理選取5株未采集過葉片的完整植株測定全株的生物量。
1.3 測定方法
1.3.1葉綠素含量測定采用直接浸提法[26]
1.3.2光合參數的測定
選擇晴朗的天氣,采用Li- 6400XT便攜式光合測定系統(美國Li-cor公司;測定毛竹葉片的氣體交換參數),測定時選用標準葉室(表面積6cm2,含內置PAR傳感器)。光合有效輻射(PAR)設定為1000μmol m-2s-1、CO2濃度(Ca)為450μmol/mol、流速為500μmol/s、溫度設定為(29±1)℃。主要測定和計算指標:凈光合速率Pn(μmol m-2s-1)、氣孔導度Gs(mmol m-2s-1)、胞間CO2濃度Ci(mmol/mol)蒸騰速率Tr(mmol m-2s-1)、氣孔限制值Ls、水分利用效率WUE(μmol/mmol)等。測定前用1000μmol m-2s-1光強對葉子進行光誘導,時間控制在15min左右,穩定后讀數,重復五次,取平均值作為測定結果。
1.3.3根系活力測定采用TTC法[27]
1.3.4毛竹葉片和根系可溶性糖含量采用蒽酮比色法測定[28]、淀粉含量采用蒽酮比色法測定[29]
1.3.5碳氮比=(可溶性糖+淀粉含量)/全氮含量[30],N含量采用凱氏定氮法測定植株全氮含量[31]
1.3.6果糖含量、蔗糖含量,采用蘇州科銘生物技術有限公司的試劑盒。
1.4 數據分析
采用Microsoft Office Excel 2007等軟件進行試驗數據的整理及計算,采用SPSS分析軟件對數據進行統計分析,采用Sigmaplot 12.5作圖。
2 結果與分析
2.1 多效唑對干旱脅迫下毛竹光合色素的影響
由表1可知,隨著干旱程度的加深,光合色素含量顯著下降(P<0.05),在重度干旱P1W3達到最小值,多效唑顯著提高了其含量(P<0.05),在P2W1達到最大值。施加外源PP333顯著增加了葉綠素a含量,P2W3較P1W1提高10.2%,P2W1較P1W1提高15.4%;顯著增加了葉綠素b含量,P2W1比P1W1提高74.3%;顯著增加了胡蘿卜含量,P2W3較P1W3提高40.8%;顯著增加了葉綠素總含量,P2W3較P1W3提高35.9%,P2W1較P1W1提高35.4%。干旱和多效唑對毛竹實生苗葉片的光合色素含量具有顯著的交互作用,干旱提升了多效唑的效應。
2.2 多效唑對干旱脅迫下毛竹實生苗氣體交換參數的影響
由表2可知,隨著干旱程度的加深,氣體交換參數值顯著下降(P<0.05),在重度干旱P1W3達到最小值,多效唑顯著提高了其含量(P<0.05)。施加外源PP333顯著提高毛竹實生苗Pn,重度干旱下提高幅度最大,達到146.9%;顯著提高Gs,P2W2較P1W2提高53.7%,P2W3較P1W3提高157.9%;顯著提高Ci,P2W2較P1W2提高22.0%,P2W3較P1W3提高68.6%;顯著提高Tr,P2W1較P1W1提高16.9%,P2W2較P1W2提高40.7%,P2W3較P1W3提高110.1%;顯著提高WUE,在P2W1達到最大值,P1W3下最小值。多效唑顯著降低Ls,在P2W1達到最小值,在重度干旱下P1W3達到最大值。干旱和多效唑對毛竹實生苗葉片的氣體交換參數具有顯著的交互作用,干旱提升了多效唑的效應。
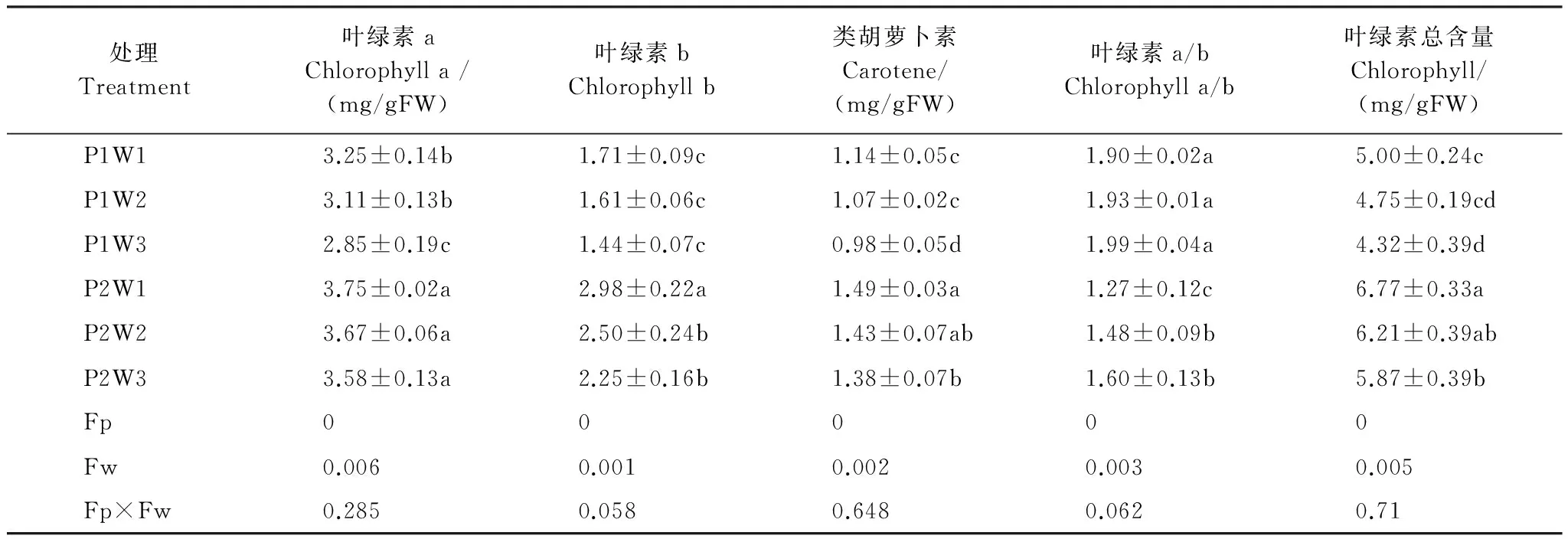
表1 多效唑對干旱脅迫下毛竹實生苗光合色素的影響
P1:0mg/L多效唑濃度,0mg/L level of PP333treatment;P2:40mg/L多效唑濃度,40mg/L level of PP333treatment;W1:75%相對田間持水量,well-watered with 75% field capacity;CK;W2:50%相對田間持水量,中度干旱water stress with 50% FC;W3:35%相對田間持水量,重度干旱water stress with 35% FC。表中誤差值為平均值的標準差,n=5;小寫字母表示該指標在同一次測定時不同處理之間是否具有顯著差異(P<0.05)
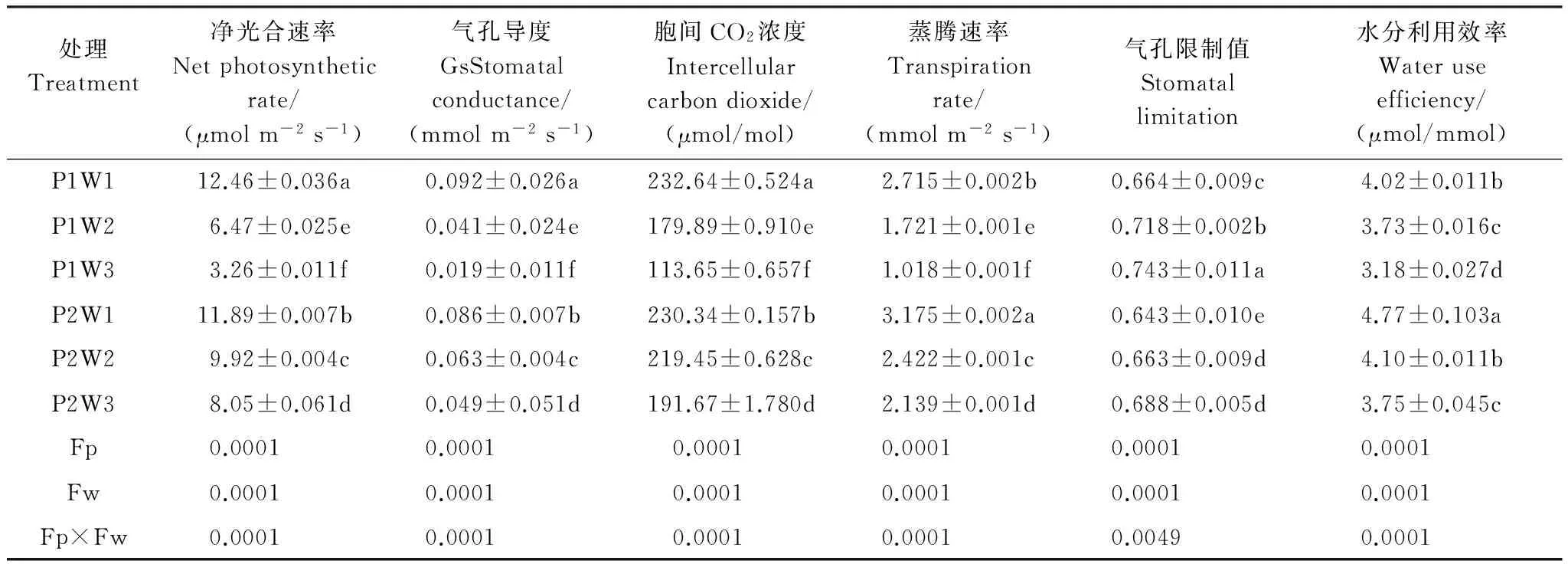
表2 多效唑對干旱脅迫下毛竹實生苗氣體交換參數的影響
FP:多效唑處理間差異,differences in PP333treatment; Fw:干旱處理間差異,differences in drought treatment
2.3 多效唑對干旱脅迫下毛竹實生苗根系活力的影響
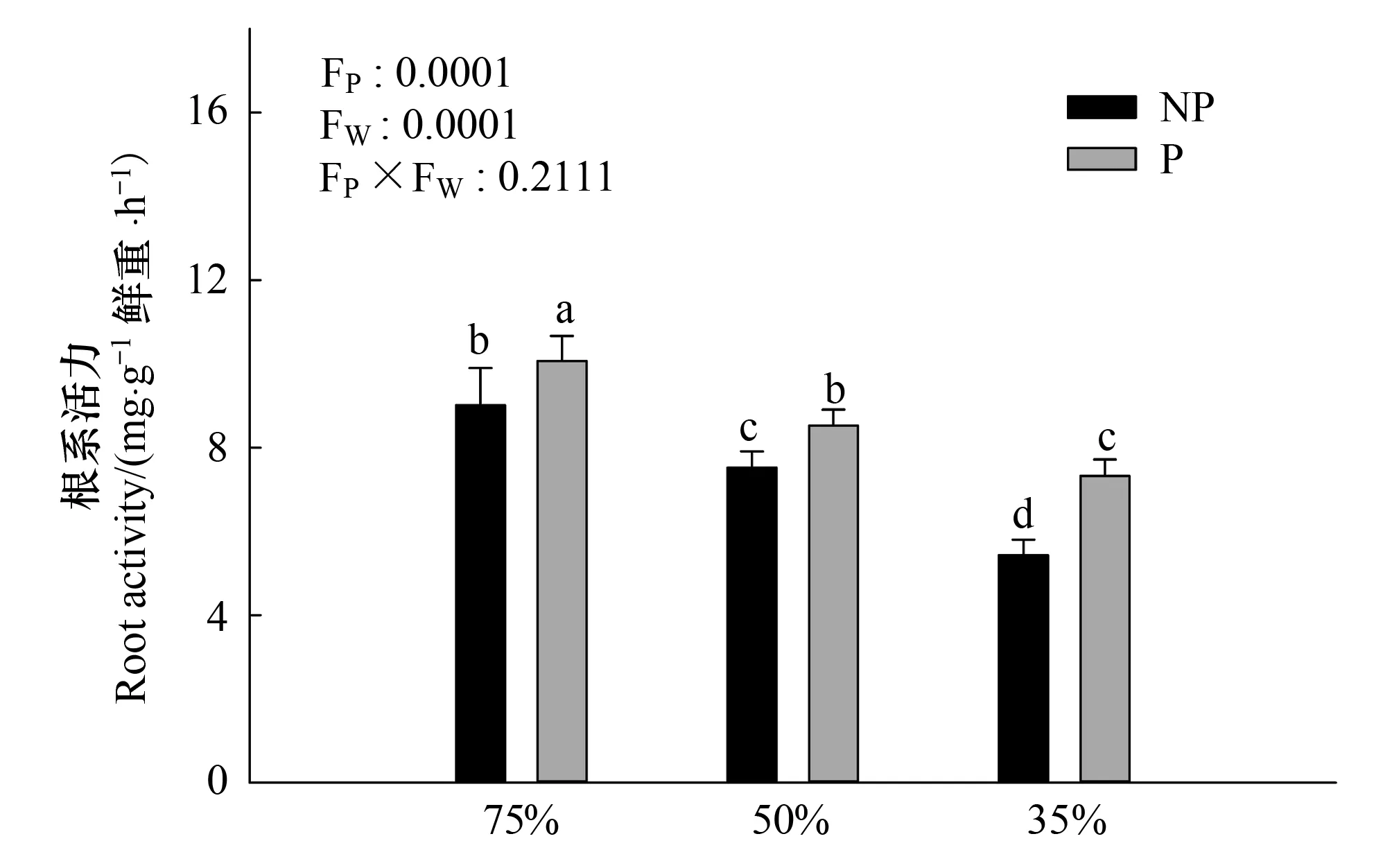
圖1 多效唑對干旱脅迫下毛竹實生苗根系活力的影響 Fig.1 The influence of PP333 on root activity of Phyllostachys edulis′ seedlings under drought stressNP:不加多效唑處理,the treatment without PP333;P:加多效唑處理,the treatment with PP333;FP:多效唑處理間差異,differences in PP333 treatment; FW:干旱處理間差異,differences in drought treatment;圖中小寫字母表示該指標在同一次測定時不同處理之間是否具有顯著差異(P<0.05)
由圖1可知,隨著干旱程度的加深,根系活力顯著減弱(P<0.05),在重度干旱情況下,根系活力最弱。相對來說,施加PP333后,顯著提高了根系活力,中度干旱情況下多效唑處理與正常供水處理差異不顯著(P>0.05),重度干旱下,多效唑提高根系活力的幅度最大,達到34.8%。干旱和多效唑對毛竹實生苗葉片的根系活力的影響具有顯著的交互作用,干旱提升了多效唑的效應。
2.4 多效唑對干旱脅迫下毛竹苗非結構碳水化合物的影響
碳水化合物在自然界中分布十分廣泛,且含量豐富,是植物光合作用的主要產物,也是植物維持生命活動的基礎物質[32],按照其存在形式,可分為結構性碳水化合物(structural carbohydrate,SC)和非結構性碳水化合物(non-structure carbohydrate,NSC),結構性碳水化合物主要包括木質素和纖維素,是植物形態建成的主要物質;非結構性碳水化合物主要包括果糖、蔗糖、淀粉、葡萄糖等參與植物生命代謝的重要物質。
2.4.1多效唑對干旱脅迫下毛竹苗可溶性總糖含量的影響
由圖2可知,隨著干旱程度的加深,毛竹葉片可溶性總糖含量顯著增加(P<0.05),在重度干旱P2W3下達到最大值3.41mg/g,施加PP333顯著增加溶性總糖的含量,在P1W1下達到最小值2.28mg/g;根系中可溶性總糖含量與葉片中變化趨勢一致。干旱和多效唑對毛竹實生苗葉片以及根系中可溶性總糖含量具有顯著的交互作用,干旱提升了多效唑的效應。
2.4.2多效唑對干旱脅迫下毛竹苗淀粉含量的影響
由圖3可知,隨著干旱程度的加深,毛竹葉片淀粉含量顯著下降(P<0.05),在P1W3達到最小值,施加PP333顯著提高了淀粉含量(P<0.05),在P2W1達到最大值31.36mg/g,在中度干旱下淀粉含量的下降幅度最大,達到26.9%;根系中淀粉含量與葉片中變化趨勢一致。干旱和多效唑對毛竹實生苗葉片以及根系中淀粉含量具有顯著的交互作用,干旱提升了多效唑的效應。
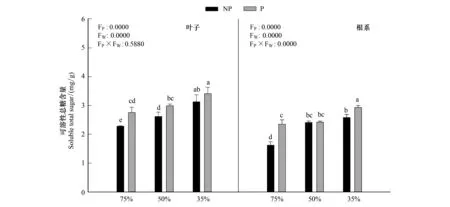
圖2 多效唑對干旱脅迫下毛竹實生苗可溶性總糖含量的影響Fig.2 The influence of PP333 on soluble total sugar of Phyllostachys edulis′ seedlings under drought stress圖中小寫字母表示該指標在同一次測定時不同處理之間是否具有顯著差異(P<0.05)
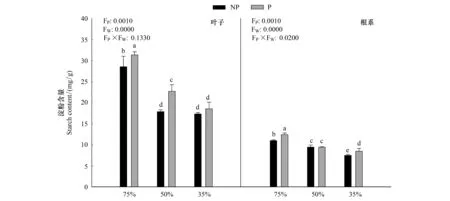
圖3 多效唑對干旱脅迫下毛竹實生苗淀粉含量的影響Fig.3 The influence of PP333 on starch content of Phyllostachys edulis′ seedlings under drought stress
2.4.3多效唑對干旱脅迫下毛竹苗蔗糖含量的影響
根據圖4可知,隨著干旱程度的加深,葉片中蔗糖含量顯著增加(P<0.05),在重度干旱下,施加PP333后,蔗糖含量顯著增加(P<0.05),P2W3較P1W3上升24.0%,P2W3較P2W1增加40.1%,在重度干旱下,達到最大值P2W3為14.64mg/g;根系中蔗糖含量與葉片中變化趨勢一致。干旱和多效唑對毛竹實生苗葉片以及根系中蔗糖含量具有顯著的交互作用,干旱提升了多效唑的效應。

圖4 多效唑對干旱脅迫下毛竹實生苗蔗糖含量的影響Fig.4 The influence of PPP333 on sucrose content of Phyllostachys edulis′ seedlings under drought stress
2.4.4多效唑對干旱脅迫下毛竹苗果糖含量的影響
由圖5可知,隨著干旱程度的加深,毛竹葉片中果糖含量顯著上升(P<0.05),在重度干旱下達到最大值P2W3為8.16mg/g,施加PP333后,果糖含量顯著增加(P<0.05),在重度干旱下上升幅度最大,P2W3較P2W1上升85.03%;根系中果糖含量與葉片中變化趨勢一致。干旱和多效唑對毛竹實生苗葉片以及根系中果糖含量具有顯著的交互作用,干旱提升了多效唑的效應。
2.5 多效唑對干旱脅迫下毛竹實生苗碳氮比的影響
由圖6可知,隨著干旱程度的加深,葉片中碳氮比值顯著增加(P<0.05),在重度干旱P1W3下達到最大值0.35,施加PP333后,在中度干旱下,碳氮比值顯著降低(P<0.05),其值達到了正常生長下的水平,重度干旱下,碳氮比差異不顯著(P>0.05)。干旱和多效唑對毛竹實生苗葉片以及根系中碳氮比的影響具有顯著的交互作用,干旱提升了多效唑的效應。
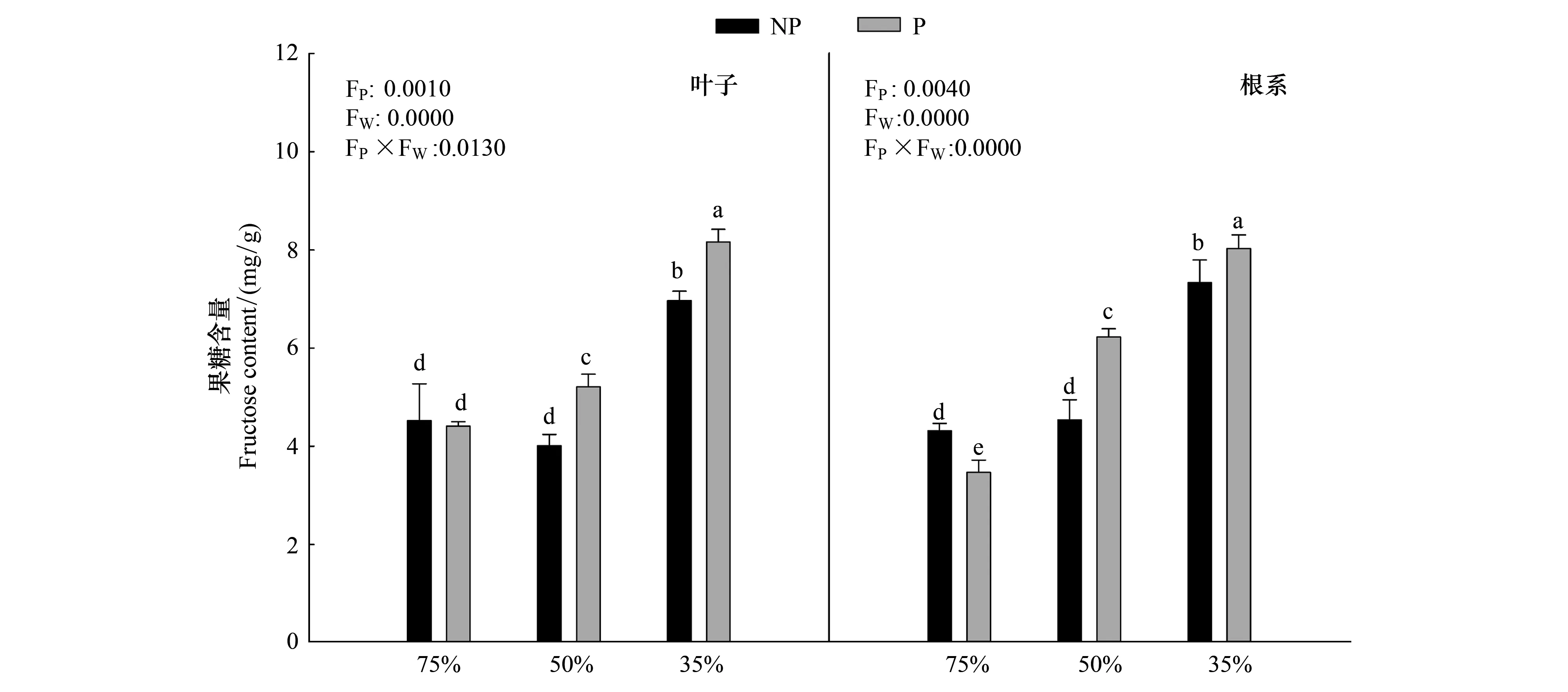
圖5 多效唑對干旱脅迫下毛竹實生苗果糖含量的影響Fig.5 The influence of PPP333 on fructose content of Phyllostachys edulis′ seedlings under drought stress圖中小寫字母表示該指標在同一次測定時不同處理之間是否具有顯著差異(P<0.05)
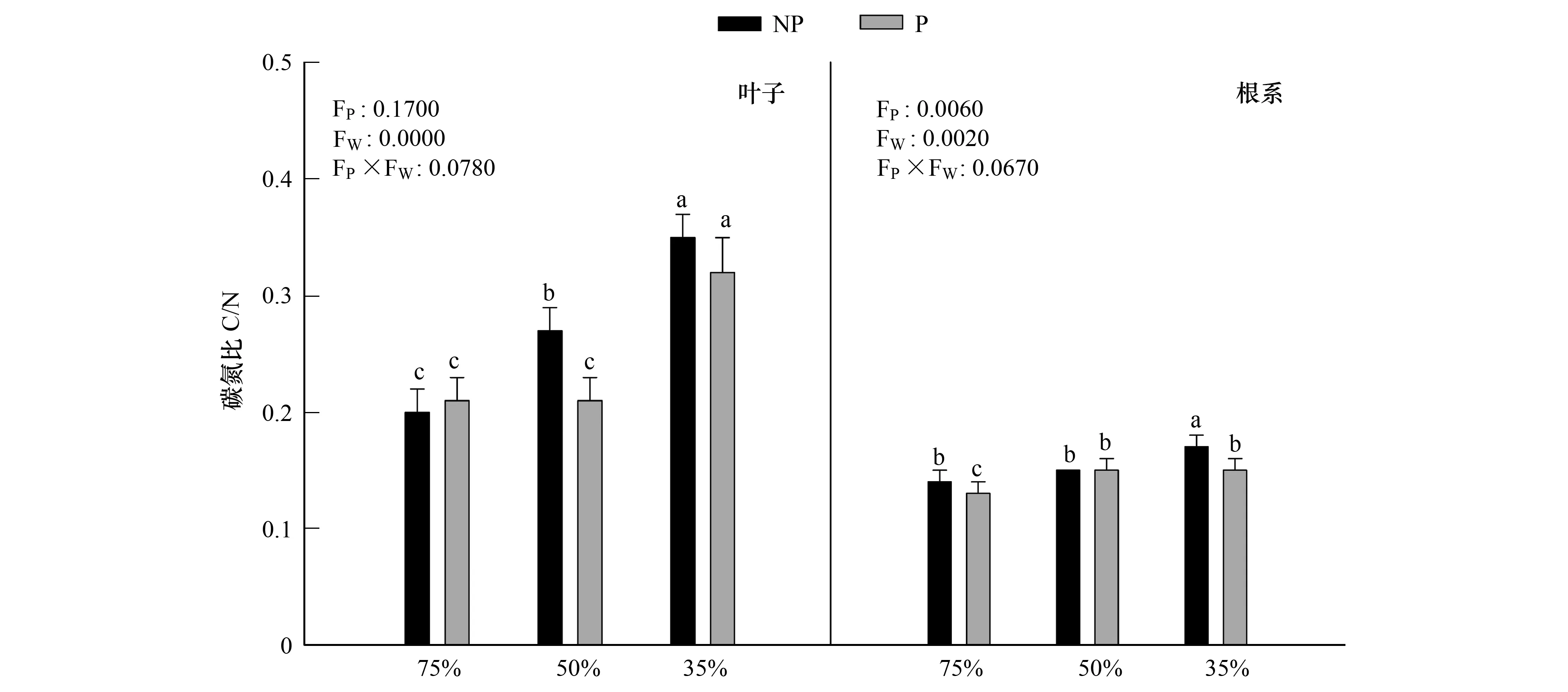
圖6 多效唑對干旱脅迫下毛竹實生苗碳氮比的影響Fig.6 The influence of PP333 on C/N of Phyllostachys edulis′ seedlings under drought stress圖中小寫字母表示該指標在同一次測定時不同處理之間是否具有顯著差異(P<0.05)
3 討論與結論
3.1 多效唑對干旱脅迫下毛竹根系活力和光合特性的影響
植物根系對水分反應敏感,其活力大小直接影響植物對礦質營養和水分的吸收利用,從而對植物的生長發育起決定性作用[33- 35],干旱脅迫會不同程度地影響到作物的各個生理過程,其中光合作用是受到最顯著影響的過程之一[36],綠色植物在進行光合作用時,主要通過葉綠體參與光合作用中光能的吸收、傳遞、分配和轉化等過程[37]。在不施加PP333情況下,隨著干旱程度加深,根系活力顯著減弱,在重度干旱下,根系活力最弱,表明重度干旱脅迫嚴重影響了毛竹根系對礦質營養和水分的吸收能力,并導致植物內部代謝紊亂,與吳敏等在栓皮櫟幼苗根系中的研究結果一致[38];葉綠素a、類胡蘿卜素、總葉綠素含量顯著下降,葉綠素b、葉綠素a/b無顯著變化,Pn、Gs、Ci、Tr、WUE顯著下降,Ls顯著上升,表明由于氣孔因素(氣孔關閉引起CO2供應受阻)[39],干旱脅迫下Gs較小,通過氣孔進入葉肉細胞的CO2減少,以至于Ls較高,這與李澤等在油桐幼苗上的研究結果一致[40]。在40mg/L的PP333處理下(P2),隨著干旱程度加劇,根系活力顯著減弱,但下降幅度顯著縮小[38];葉綠素a無顯著變化,葉綠素b、類胡蘿卜素、葉綠素總含量顯著下降,葉綠素a/b顯著上升,Pn、Gs、Ci、Tr、WUE顯著下降,Ls顯著上升,表明多效唑緩解干旱下植物蒸騰速率的下降、氣孔的收縮、氣孔導度降低、保持CO2向葉綠體內輸送,進而保持植物葉片正常的光合作用和蒸騰作用[14]。在不同干旱程度下,毛竹實生苗的根系活力、光合色素、氣體交換參數對PP333的響應規律不同,在正常水分處理下(W1),PP333處理顯著提高了根系活力,在P2W1下達到最大值;顯著提高了光合色素,葉綠素a/b顯著下降,葉綠素總含量在P2W1下達到最大值,Pn、Gs、Ci、Ls顯著下降,Tr、WUE顯著上升,表明說明多效唑能有效提高干旱脅迫下毛竹實生苗的光合同化能力,這與高玲等(2008)在草坪草的研究一致,水分利用效率是植物光合和蒸騰特性的綜合反映,WUE提高,說明植物對水分利用比較充分、抗旱性較強,是植物在逆境環境下長期進化的一種自我保護機制[41]。在中度干旱處理下(W2),PP333處理顯著提高了根系活力,提升幅度較小;顯著提高了光合色素,葉綠素a/b顯著下降,Pn、Gs、Ci、Tr、WUE顯著上升,Ls顯著下降,表明施加外源PP333顯著增加了各參數水平。在重度干旱處理下(W3),PP333處理后,根系活力顯著提高,提高幅度最大;光合色素顯著提高,葉綠素a/b在P1W3處理下達到最大值,Pn、Gs、Ci、Tr、WUE顯著上升,Ls顯著下降,由于光合機構活性破壞降低葉肉細胞中的Ci未能及時同化固定,從而使葉肉細胞中的CO2濃度積累增加,因此Ls逐漸降低。
3.2 多效唑對毛竹干旱脅迫下NSC和C/N的影響
非結構性碳水化合物(NSC)主要包括蔗糖、淀粉、果糖等參與植物生命代謝的重要物質,是植物體內最主要的滲透調節物質[42],其分配格局是多種生態生理過程作用的綜合結果[43],與植物的生存策略密切相關[44],氮是重要的非礦質元素,對維持葉綠素含量和提高葉片光合具有重要作用[45],碳氮代謝是植物最基本的代謝過程,氮代謝需要碳代謝提供能量,而碳代謝需要氮代謝提供光合色素(碳氮比是光合產物分配方向的重要指標)[10]。本研究發現,在PP333處理后,毛竹實生苗的非結構性碳水化合物含量、碳氮比對干旱程度加深的響應規律不同,在0mg/L的PP333處理下(P1),隨著干旱程度加深,葉片和根系的可溶性總糖、蔗糖、果糖含量顯著升高,蔗糖在P1W2、P1W3處理下差異不顯著,果糖在P1W1、P1W2處理下差異不顯著,葉片及根系里的淀粉含量顯著下降,在P1W2、P1W3處理下差異不顯著,表明淀粉轉化為可溶性糖,生理代謝活動增強,來吸收和運輸水分,用于生長需求;葉片及根系里的碳氮比顯著上升,但在P1W2、P1W3處理下差異不顯著,由于植物的光合同化能力降低,光合產物輸出減少,形成了光合作用與呼吸作用之間碳源供應的不平衡,葉片中淀粉水解,糖類等有機物堆積[42],儲存于毛竹實生苗體內NSC含量增加,來抵御逆境,導致碳過剩。在40mg/L的PP333處理下(P2),隨著干旱程度加深,葉片及根系里的可溶性總糖和果糖含量顯著上升,葉子里的蔗糖含量顯著上升,根系里的蔗糖含量先下降后升高,葉片及根系里的淀粉含量顯著下降,由于可溶性糖更多地參與到形態建成,淀粉發生水解,以補充植物體對可溶性糖的需求;葉片及根系里的碳氮比顯著上升。在不同干旱程度下,毛竹實生苗的非結構性碳水化合物含量、碳氮比對PP333的響應規律不同,在正常水分處理下(W1),PP333處理后,葉片及根系里的可溶性總糖、淀粉、蔗糖顯著上升,果糖在葉片中的含量差異不顯著,在根系中顯著下降,由于光合作用的提高,將可溶性糖轉化為淀粉,NSC含量增加,施加外源PP333葉片淀粉含量顯著增加,分解速度下降,葉片總糖含量顯著上升,這結果與鄭日如對百合研究[46],陳曉光等在甘薯上的研究一致[47];碳氮比在葉片中差異不顯著,在根系中顯著下降;在中度干旱處理下(W2),PP333處理后,葉子里的可溶性總糖、淀粉含量下顯著增加,根系里無顯著變化,果糖在葉片和根系中顯著上升,碳氮比在葉片中顯著下降,在根系中差異不顯著,多效唑顯著增加淀粉、糖含量,促進光合作用,提供光合色素含量,顯著降低了碳氮比,在中度干旱下,使碳氮比達到正常生長下的毛竹,說明多效唑有利于維護干旱下毛竹實生苗的正常生長。這結果與陳炫等在荔枝上的研究一致[48];在重度干旱處理下(W3),PP333處理后,葉片里的可溶性糖、淀粉含量差異不顯著,根系中的含量顯著上升,葉片及根系里的蔗糖、果糖含量顯著上升,葉片中碳氮比差異不顯著,根系中顯著下降,光合作用受到抑制,光合產物合成受阻,淀粉加速分解為糖為光合作用提供能量、參與滲透調節,碳升高,葉綠素含量下降,導致氮素含量下降,毛竹葉片以及根系中碳氮代謝顯著上升,碳氮比失衡。
綜上,干旱脅迫下毛竹實生苗幼苗生長受到顯著抑制,主要表現為根系活力減弱,氣孔導度降低導致光合能力下降。施加多效唑(PP333)可顯著提高毛竹抗旱性,,具體表現為:根系活力顯著提高,以獲取更多的水分和礦物元素促進毛竹體內營養物質合成和生命活動調節;通過氣孔導度調節提升光合能力和水分利用效率,協調碳同化,保持碳氮比平衡同時提高毛竹實生苗體內NSC的儲存量進而提升其抗旱能力。
參考文獻(References):
[1]Mueller B, Seneviratne S I. Hot days induced by precipitation deficits at the global scale. Proceedings of the National Academy of Sciences of the United States of America, 2012, 109(31):12398- 12403.
[2]Naika M, Shameer K, Mathew O K, Gowda R, Sowdhamini R. STIFDB2: an updated version of plant stress-responsive transcription factor database with additional stress signals, stress-responsive transcription factor binding sites and stress-responsive genes in Arabidopsis and rice. Plant and Cell Physiology, 2013, 54(2): e8.
[3]Athar H R, Ashraf M. Strategies for crop improvement against salinity and drought stress: an overview // Ashraf M, Ozturk M, Athar H R, eds. Salinity and Water Stress. Netherlands: Springer, 2008: 1- 16.
[4]Cramer G R, Urano K, Delrot S, Pezzotti M, Shinozaki K. Effects of abiotic stress on plants: a systems biology perspective. BMC Plant Biology, 2011, 11(1): 163.
[5]劉靜, 魏開發, 高志暉, 李冰冰, 任慧波, 胡建芳, 賈文鎖. 干旱脅迫下氮素營養與根信號在氣孔運動調控中的協同作用. 植物學通報, 2008, 25(1): 34- 40.
[6]Paez-Garcia A, Motes C M, Scheible W R, Chen R J, Blancaflor E B, Monteros M J. Root traits and phenotyping strategies for plant improvement. Plants, 2015, 4(2): 334- 355.
[7]Ali M A, Abbas A, Niaz S, Zulkiffa M, Ali S. Morpho-physiological criteria for drought tolerance in sorghum (Sorghumbicolor) at seedling and post-anthesis stages. International Journal of Agriculture & Biology, 2009, 11(6): 674- 680.
[8]Bartels D, Sunkar R. Drought and salt tolerance in plants. Critical Reviews in Plant Sciences, 2005, 24(1): 23- 58.
[9]Zhang Y J, Xie Z K, Wang Y J, Su P X, An L P, Gao H. Effect of water stress on leaf photosynthesis, chlorophyll content, and growth of oriental lily. Russian Journal of Plant Physiology, 2011, 58(5): 844- 850.
[10]蔣思思, 魏麗萍, 楊松, 肖迪, 王曉潔, 侯繼華. 不同種源油松幼苗的光合色素和非結構性碳水化合物對模擬氮沉降的短期響應. 生態學報, 2015, 35(21): 7061- 7070.
[11]李東勝, 史作民, 劉世榮,耿麗君. 南北樣帶溫帶區櫟屬樹種種子化學組成與氣候因子的關系. 生態學報, 2012, 32(24):7857- 7865.
[12]于麗敏, 王傳寬, 王興昌. 三種溫帶樹種非結構性碳水化合物的分配. 植物生態學報, 2011, 35(12):1245- 1255.
[13]李娜妮, 何念鵬, 于貴瑞. 中國東北典型森林生態系統植物葉片的非結構性碳水化合物研究. 生態學報, 2016, 36(2):430- 438.
[14]高玲, 劉國道. 植物生長調節劑——多效唑在草坪草上的應用進展. 熱帶農業科學, 2008, 28(4): 98- 102.
[15]劉靜雅, 李紹才, 孫海龍, 宋海鳳, 陳艷華. 多效唑對紫穗槐生長及生理特性的影響. 植物科學學報, 2016, 34(2): 271- 279.
[16]楊忠義, 范春暉, 郭平毅. 氮肥與多效唑對冬小麥葉片生理功能的調控. 植物營養與肥料學報, 2008, 14(5): 947- 950.
[17]毛軼清, 鄭青松, 陳健妙, 劉兆普, 劉國紅, 姜超強. 噴施多效唑提高麻瘋樹幼苗耐鹽性的生理機制. 生態學報, 2011, 31(15): 4334- 4341.
[18]唐鳳德, 蔡天革, 韓士杰, 黃麗榮. 生物制劑對沙地樟子松苗木成活生長及生理特征的影響. 生態學報, 2009, 29(5): 2294- 2303.
[19]宋海鳳, 李紹才, 孫海龍, 劉靜雅, 陳艷華. 根施不同濃度多效唑對紫穗槐生長特性和相關生理指標的影響. 植物生理學報, 2015, 51(9): 1495- 1501.
[20]呂雙慶, 李生秀. 多效唑對旱地小麥一些生理、生育特性及產量的影響. 植物營養與肥料學報, 2005, 11(1): 92- 98.
[21]王競紅, 多多. 多效唑對6種草坪草苗期抗旱性影響的研究. 草業學報, 2014, 23(6): 253- 258.
[22]McCann S E, Huang B R. Effects of Trinexapac-ethyl foliar application on creeping bentgrass responses to combined drought and heat stress. Crop Science, 2007, 47(5): 2121- 2128.
[23]McCann S E, Huang B R. Drought responses of Kentucky bluegrass and creeping bentgrass as affected by abscisic acid and Trinexapac-ethyl. Journal of the American Society for Horticultural Science, 2008, 133(1): 20- 26.
[24]Chan Z L, Shi H T. Improved abiotic stress tolerance of bermudagrass by exogenous small molecules. Plant Signaling & Behavior, 2015, 10(3): e991577.
[25]莊若楠, 金愛武. 施肥對毛竹稈型特征的影響. 中南林業科技大學學報, 2013, 33(1): 80- 84.
[26]鄒琦. 植物生理學實驗指導. 北京: 中國農業出版社, 2000.
[27]程兆偉, 鄒應斌, 劉武. 水稻根系研究進展. 作物研究, 2006, 20(5): 504- 508.
[28]張振清. 植物生理學試驗手冊. 上海: 上海科學技術出版社, 1985: 134- 138.
[29]姜東, 于振文, 李永庚, 余松烈, 孔蘭靜. 高產小麥營養器官臨時貯存物質積運及其對粒重的貢獻. 作物學報, 2003, 29(1): 31- 36.
[30]Breeman A M. Relative importance of temperature and other factors in determining geographic boundaries of seaweeds: experimental and phenological evidence. Helgol?nder Meeresuntersuchungen, 1988, 42(2): 199- 241.
[31]魯如坤. 土壤農業化學分析方法. 北京: 中國農業科技出版社, 1999.
[32]雷美玉, 李輝亮, 劉立元, 彭世清. 糖在植物中的感知與信號傳導研究進展.西北植物學報, 2007, 27(10): 2118- 2127.
[33]衛星, 王政權, 張國珍,陳海波,王婧. 水曲柳苗木不同根序對干旱脅迫的生理生化反應. 林業科學, 2009, 45(6):16- 21.
[34]李香玲, 馮躍華.水稻根系生長特性及其與地上部分關系的研究進展. 中國農學通報, 2015, 31(6): 1- 6.
[35]Hu H H, Xiong L Z. Genetic engineering and breeding of drought-resistant crops. Annual Review of Plant Biology, 2013, 65: 715- 741.
[36]Hosseinzadeh S R, Amiri H, Ismaili A. Effect of vermicompost fertilizer on photosynthetic characteristics of chickpea (CicerarietinumL.) under drought stress. Photosynthetica, 2016, 54(1): 87- 92.
[37]張棟. 干旱脅迫對蘋果光合作用和葉綠素熒光的影響及葉片衰老特性研究[D]. 楊凌: 西北農林科技大學, 2011.
[38]吳敏, 張文輝, 周建云,馬闖,韓文娟. 干旱脅迫對栓皮櫟幼苗細根的生長與生理生化指標的影響. 生態學報, 2014, 34(15):4223- 4233.
[39]裴斌, 張光燦, 張淑勇,吳芹,徐志強,徐萍. 土壤干旱脅迫對沙棘葉片光合作用和抗氧化酶活性的影響. 生態學報, 2013, 33(5):1386- 1396.
[40]李澤, 譚曉風, 盧錕,張琳,龍洪旭,呂佳斌,林青. 干旱脅迫對兩種油桐幼苗生長、氣體交換及葉綠素熒光參數的影響. 生態學報, 2017, 37(5):1515- 1524.
[41]李磊, 賈志清, 朱雅娟, 綦艷林. 我國干旱區植物抗旱機理研究進展. 中國沙漠, 2010, 30(5): 1053- 1059.
[42]熊炳霖. 干旱脅迫下氮素對玉米葉片衰老和葉片碳氮平衡的影響[D]. 北京: 中國科學院研究生院(教育部水土保持與生態環境研究中心), 2016.
[43]王凱, 雷虹, 夏揚,于國慶. 楊樹幼苗非結構性碳水化合物對增加降水和氮添加的響應. 應用生態學報, 2017, 28(2):399- 407.
[44]Imaji A, Seiwa K. Carbon allocation to defense, storage, and growth in seedlings of two temperate broad-leaved tree species. Oecologia, 2010, 162(2): 273- 281.
[45]張立軍, 劉新. 植物生理學(第二版). 北京: 科學出版社, 2011.
[46]鄭日如. 生長延緩劑對東方百合植株生長和鱗莖養分代謝的影響研究[D]. 杭州: 浙江大學, 2011.
[47]陳曉光, 李洪民, 張愛君, 史新敏, 唐忠厚, 魏猛, 史春余. 不同氮水平下多效唑對食用型甘薯光合和淀粉積累的影響. 作物學報, 2012, 38(9): 1728- 1733.
[48]陳炫, 陶忠良, 吳志祥, 周兆德, 王令霞. 多效唑+乙烯利對妃子笑荔枝內源激素及碳氮營養的影響. 江西農業大學學報, 2012, 34(1): 27- 33.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
