不同小白菜品種硝酸鹽含量、氮代謝關鍵酶活性及NRT1表達和亞細胞定位
2018-04-24 12:01:26李彥華徐衛紅周鑫斌王衛中遲蓀琳張春來趙婉伊秦余麗王正銀謝德體
食品科學 2018年7期
李彥華,楊 蕓,徐衛紅*,周鑫斌,王衛中,遲蓀琳,李 桃,張春來,趙婉伊,秦余麗,王正銀,謝德體
蔬菜極易累積硝酸鹽,在人體所攝入的硝酸鹽中,其貢獻高達72%~94%[1-2]。雖然過量的硝酸鹽對植物無害,但大量的NO3-通過食物鏈進入人體腸胃后,可經細菌的作用還原為亞硝酸或與胺反應生成亞硝胺,從而可能使血液載氧能力降低、誘發人體消化系統癌變等[3-6]。目前我國不少大城市葉類蔬菜的硝酸鹽含量偏高,部分地區高達3 000 mg/kg[7]。氮肥的不合理施用被認為是作物硝酸鹽積累的原因,蔬菜中的硝酸鹽含量會隨氮肥施用量的增加迅速升高[8]。而且,不同形態的氮肥(如銨態氮和硝態氮)及其比例是影響蔬菜硝酸鹽含量的重要因素[9-10]。有研究表明,增加銨態氮含量可以顯著降低葉類蔬菜莖葉中硝酸鹽的含量[11-13]。
不同種蔬菜及同種蔬菜的不同品種間硝酸鹽含量有差異[13],且硝酸鹽積累量的差異變幅可高達1 600 mg/kg[14]。硝酸還原酶(nitrate reductase,NR)活性、硝酸鹽轉運蛋白(nitrate transporter,NRT)等遺傳因子差異是造成積累差異的主要原因;其中,NRT基因對于氮素吸收、運輸和同化有著重要作用,是提高作物氮素利用效率,解決硝酸鹽積累的重要研究對象[11]。NO3-—N主要是通過低親和的NRT1和高親和的NRT2而被高等植物吸收,且這兩類轉運體基因已在擬南芥、水稻和番茄中得到克隆[15]。而目前有關蔬菜NRT基因的表達和亞細胞定位,以及硝銨比、氮代謝酶與蔬菜的硝酸鹽含量相關性的研究報道較少,且結果不一致[16-18]。
小白菜(Brassica chinensis L.)是我國最重要的葉類蔬菜之一。都韶婷等[7]檢測了我國多個城市的主要蔬菜硝酸鹽含量,發現小白菜是硝酸鹽含量最高的蔬菜之一,其中北京和長沙地區的硝酸鹽含量分別高達3 615 mg/kg和3 514 mg/kg,超出了葉類蔬菜的硝酸鹽限量標準;因此,本研究采用液體培養實驗,以低硝酸鹽富集品種‘香港特選奶白菜’和高硝酸鹽富集品種‘揭農四號春白菜’為供試材料,系統探討了硝銨比對2 種小白菜硝酸鹽含量、氮代謝關鍵酶活性的影響及NRT基因的表達和亞細胞定位在小白菜不同品種間的差異,初步明確硝酸鹽積累的生理及分子機理;同時也為蔬菜生產中的氮肥運籌及栽培品種合理選擇提供理論依據。
1 材料與方法
1.1 材料與試劑
供試作物為小白菜,品種為前期實驗中篩選出的低硝酸鹽富集品種‘香港特選奶白菜’和高硝酸鹽富集品種‘揭農四號春白菜’[19]。
小量RNA抽提試劑盒 華舜生物公司;Taq DNA聚合酶 上海鼎國公司;RNase Free-DNaseⅠ、SYBR Premix Ex TaqTMⅡ試劑盒、高保真酶Pfu、dNTP 日本TaKaRa公司;Trizol RNA提取試劑盒 美國Invitrogen公司;逆轉錄酶(M-MLV) 美國Fermentas公司;PCR合成試劑 上海生工生物工程公司;膠回收試劑盒美國Axygen公司;酶解液、PEG4000溶液、W5溶液、MMG溶液 武漢伯遠生物技術公司。
1.2 儀器與設備
ABI-9700聚合酶鏈式反應(polymerase chain reaction,PCR)儀 美國ABI公司;Nanodrop2000紫外-可見分光光度計 美國Thermo Scientific公司;HE99電泳儀 美國GE公司;WD-9403C紫外透射儀 北京市六一儀器廠;LSM700激光共聚焦顯微鏡 德國Zeiss公司。
1.3 方法
實驗共設3 個硝銨比處理組(c(NO3-)∶c(NH4+)分別為50∶50、75∶25和100∶0),營養液按霍格蘭營養液配方配制,氮濃度為16 mmol/L,調節pH值至5.8~6.0,具體操作參見文獻[19]。
1.3.1 植株硝酸鹽含量及氮代謝關鍵酶活力測定
硝酸鹽含量的測定采用GB 5009.33—2010《食品中亞硝酸鹽與硝酸鹽的測定》方法;NR活力測定采用活體法[20-21];亞硝酸還原酶(nitrite reductase,NiR)活力測定參照Ozawa等[22]的方法;谷氨酰胺合成酶(glutamine synthetase,GS)活力參照植物生理學實驗指導[23]測定;谷氨酸合成酶(glutamate synthase,GOGAT)和谷氨酸脫氫酶(glutamate dehydrogenase,GDH)活力參照葉利庭等[24]的方法測定。
1.3.2 熒光定量PCR
1.3.2.1 總RNA提取與檢測
采用華舜生物公司的小量RNA抽提試劑盒提取,并通過電泳結果以及RNA濃度綜合評價RNA的質量。
1.3.2.2 RNA的純化與反轉錄
使用PrimeScript? RT reagent Kit with gDNA Eraser進行RNA的反轉錄,并采用RNase Free-DNaseⅠ除去RNA中微量的DNA。將所得的cDNA置于-20 ℃冰箱保存備用。
1.3.2.3 引物設計與合成
本實驗研究硝酸鹽低親和轉運蛋白基因NRT1。由武漢伯遠生物技術公司設計并合成NRT1基因的反轉錄(reverse transcription,RT)-PCR特異引物,內參引物為actin-F/R(表1)。
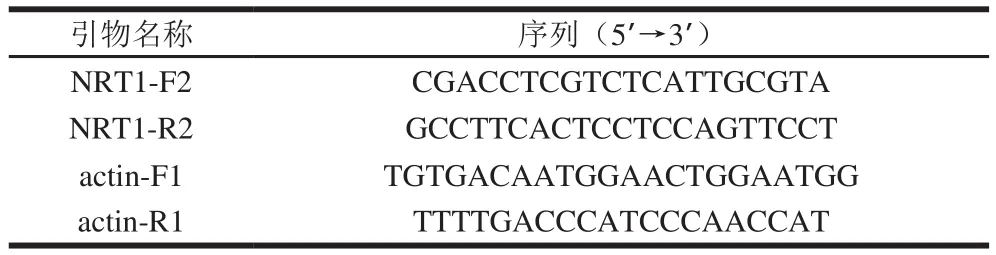
表1 NRT1基因的RT-PCR引物Table 1 Primers used for RT-PCR of NRT1
1.3.2.4 cDNA的PCR擴增
取獲得的cDNA,用ABI-9700 PRC儀進行特異擴增。反應體系的總體積為25 μL,包括2.5 μL 10×Buffer(含Mg2+)、0.5 μL dNTP(10 mmol/L)、正反向引物(10 μmol/L)各0.5 μL、Taq DNA聚合酶(5 U/μL)0.3 μL、cDNA模板0.25 μL,剩余部分用ddH2O補足。PCR程序為:94 ℃預變性2 min,94 ℃變性40 s,退火(溫度為55 ℃)1 min,72 ℃ 45 s,30 個循環,72 ℃ 10 min。PCR產物在1.5%的瓊脂糖凝膠中電泳檢測,并用紫外透射儀拍照。
1.3.2.5 實時熒光定量PCR
將反轉錄得到的cDNA用ddH2O稀釋25 倍,采用SYBR Premix Ex TaqTMⅡ試劑盒,進行qRT-PCR,檢測其特異性。操作方法參照使用說明書進行,每個試樣重復3 次。
1.3.3 NRT1亞細胞定位
菌種:大腸桿菌用作質粒的轉化宿主;質粒:pBWA(V)H-YFPNR1;融合表達載體的構建:參照楊學東等[25]的NRT1序列設計引物,以NRT1克隆為模板用高保真酶Pfu進行PCR擴增,PCR產物經電泳回收后連接到pBWA(V)H-YFPNR1載體上,獲得最終表達載體(圖1)。

圖1 原生質體亞細胞定位載體圖譜Fig. 1 Map of the protoplast subcellular localization vector
質粒定位原生質體:取小白菜葉用水沖洗表面污垢,在干凈塑料/玻璃板上用鋒利的刀片切成碎段(長度小于0.5 mm),總質量5~10 g左右。加入5~10 mL酶解液全部浸泡組織,28 ℃緩慢振蕩(100 r/min)酶解5~6 h。將原生質體用40 μm濾網進行過濾,然后50×g離心10 min,可見渾濁沉淀。直接倒去上清液,用預冷W5溶液10 mL洗滌2 次,每次50×g離心5 min,可見管底渾濁沉淀。根據需要加入500 μL MMG溶液稀釋原生質體得到原生質體原液。使其在40 倍下顯微鏡檢測每個視野20~40 個左右。取200 μL原生質體懸液原液,等體積的PEG溶液及500 ng以上的純化質粒進行輕柔均勻混合,室溫靜置30 min,用1 mL W5稀釋,混合均勻終止反應,100×g離心5 min收集原生質體,加入1 mL W5洗1~2 次,最后加入1 mL W5溶液重懸,轉移到2 mL EP管中,28 ℃暗培養24~48 h,激光共聚焦顯微鏡觀察。
1.4 數據處理
采用Excel 2010和SPSS 23.0軟件進行統計處理,利用Duncan’s新復極差法做顯著性分析。
2 結果與分析
2.1 硝銨比對不同品種小白菜各部位硝酸鹽含量的影響

圖2 硝銨比對不同品種小白菜葉(A)、葉柄(B)、根(C)硝酸鹽含量的影響Fig. 2 Effect of NO3-/NH4+ ratio on nitrate contents in leaves (A),petioles (B) and roots (C) of two varieties of pakchoi
從圖2可以看出,在不同硝銨比處理下,小白菜硝酸鹽含量為葉柄>葉>根。隨NO3-比例的增加,小白菜葉柄、葉和根硝酸鹽含量總體呈增加趨勢。在硝銨比75∶25和100∶0處理中,‘香港特選奶白菜’和‘揭農四號春白菜’葉片硝酸鹽含量分別較硝銨比50∶50處理增加了7.9%~13.2%和29.1%~30.4%,葉柄硝酸鹽含量分別較硝銨比50∶50處理增加了0.3%~14.3%和3.3%~4.9%,根硝酸鹽含量分別較硝銨比50∶50處理增加了45.4%~73.8%和9.8%~63.3%。相同硝銨比處理下,‘香港特選奶白菜’葉、葉柄和根中的硝酸鹽含量均明顯低于‘揭農四號春白菜’。
2.2 硝銨比對不同品種小白菜氮代謝相關酶活性的影響


圖3 不同硝銨比對2 個品種小白菜葉片NR(A)、NiR(B)、GS(C)、GOGAT(D)、GDH(E)活力的影響Fig. 3 Effect of NO3-/NH4+ ratio on the activities of NR (A), NiR (B),GS (C), GOGAT (D) and GDH (E) in leaves of two varieties of pakchoi
由圖3可知,隨著硝銨比的增加小白菜葉片NR活力呈現降低趨勢,在硝銨比75∶25和100∶0處理中,‘香港特選奶白菜’和‘揭農四號春白菜’葉片NR活性分別較硝銨比50∶50處理降低了15.5%~26.6%和21.9%~26.5%。‘香港特選奶白菜’和‘揭農四號春白菜’葉片NiR活力隨著硝銨比的增加有所增加,硝銨比75∶25和100∶0處理分別較硝銨比50∶50處理增加了38.9%~57.3%和5.1%~74.9%。小白菜葉片GOGAT活力隨著硝銨比的增加呈現降低趨勢,硝銨比75∶25和100∶0處理‘香港特選奶白菜’和‘揭農四號春白菜’葉片GOGAT活力分別較硝銨比50∶50處理降低了10.5%~15.5%和0.2%~24.6%。隨著硝銨比的增加小白菜葉片GDH活力呈先增加后降低趨勢,‘香港特選奶白菜’和‘揭農四號春白菜’葉片GDH活力在75∶25時達到最大值,分別較硝銨比50∶50處理增加了2.1%和20.0%。而不同硝銨比對2 個品種小白菜的GS活力沒有顯著影響。不同硝銨比處理中,除硝銨比50∶50處理下的NiR、GDH活力外,其他氮代謝相關酶活力在兩個小白菜品種之間達到顯著性差異。
2.3 小白菜NRT1基因表達分析
高等植物有兩種細胞質膜NRT,即NRTl和NRT2。NRT2是高親和硝酸鹽轉運系統(high-affinity transport system,HATs),而大部分NRTl是低親和硝酸鹽轉運系統(low-affinity transport system,LATs)。根據以上實驗結果分別選擇高硝酸鹽富集品種‘揭農四號春白菜’和低硝酸鹽富集品種‘香港特選奶白菜’為材料,對其葉片部位的RNA進行實時熒光定量PCR。如圖4所示,NRT1在‘香港特選奶白菜’和‘揭農四號春白菜’葉片中均有表達;以‘香港特選奶白菜’NRT1的平均相對表達量為1.00,‘揭農四號春白菜’的NRT1的平均相對表達量為2.47,‘揭農四號春白菜’的NRT1相對表達量顯著高于‘香港特選奶白菜’,且2 個白菜品種葉片中NRT1表達量組內差異均較大。但未檢測出NRT2表達量。
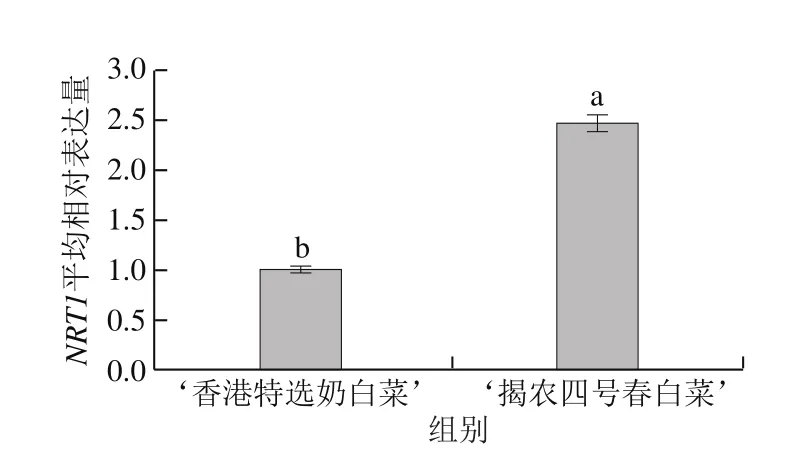
圖4 小白菜葉片中NRT1實時定量熒光PCR分析Fig. 4 Real-time PCR analysis of NRT1-associated genes in leaves of pakchoi
2.4 小白菜NRT1亞細胞定位

圖5 NRT1在小白菜葉片原生質體中的亞細胞定位Fig. 5 Subcellular localization of NRT1 in pakchoi protoplasts
通過生物信息學方法分析蛋白可能的信號肽或功能域來確定蛋白在細胞內的定位,有利于高效地探索蛋白的功能。近年來綠色熒光蛋白被廣泛應用于基因表達、蛋白質定位等領域。當其在細胞中表達時,若遇到藍光或紫外光,便會產生明亮的綠色熒光。為了研究NRT1在植物細胞中的作用部位,以高硝酸鹽富集品種‘揭農四號春白菜’和低硝酸鹽富集品種‘香港特選奶白菜’為材料,構建了原生質體表達載體。如圖5所示,NRT1在小白菜葉片原生質體中的亞細胞定位結果表明,NRT1在原生質膜上大量表達,而在原生質體內并無表達,說明NRT1是定位于原生質體細胞膜上的非跨膜蛋白。
3 討 論
艾紹英等[26]研究發現適當增加營養液中銨態氮的比例可有效降低菜心中的硝酸鹽含量。在本實驗中,2 個品種小白菜葉片、葉柄和根的硝酸鹽含量隨著銨態氮比例的增加而降低,隨硝態氮比例的增加而增加。Ullrich[27]認為其可能的原因是銨能夠高速進入細胞,而導致細胞膜去極化,阻止了H+與陰離子的協同運輸,因此抑制了硝酸鹽的吸收;也可能是因為受到了溫度、光照等外部因素的影響[28-31]。供試2 個品種小白菜體內硝酸鹽含量表現為葉柄>葉片>根,這與Chen Wei等[32]報道一致。Gransteck等[33]認為這是由于根和葉主要承擔物質的吸收和同化的作用,進入根部后的NO3-部分被同化為營養物質,而大部分NO3-則在蒸騰拉力的作用下經由木質部輸送到葉片、果實等地上部位,參與各種生理代謝。供試低硝酸鹽富集品種‘香港特選奶白菜’,高富集品種‘揭農四號春白菜’,2 個小白菜品種之間葉片硝酸鹽含量相差1.22~1.46 mg/kg,葉柄硝酸鹽含量相差1.21~1.40 mg/kg,根硝酸鹽含量相差1.80~2.38 mg/kg,這種蔬菜種間、品種間硝酸鹽積累差異原因主要由遺傳因子控制所致。有研究表明,蔬菜硝酸鹽積累較高和較低的兩極類群品種表現穩定,基因型差異可穩定遺傳[11]。
硝酸鹽被特異性的NRT高效運輸到植物根細胞中,儲存在液泡里,在細胞質中被NR降解成亞硝酸鹽。亞硝酸鹽進入根的葉綠體或非綠色組織的質體中,在6-鐵氧還蛋白的幫助下NiR將其進一步還原成銨。隨后在ATP供能下,GS將谷氨酸和NH4+轉換成谷氨酰胺,該過程是在葉綠體、細胞質或根細胞質體中進行的,最終銨參與到氨基酸的合成。當植物細胞中NH4+濃度較高時,NADH提供電子,NH4+還可以由存在于葉綠體和線粒體中的GDH還原為谷氨酸。植物體內,NR和NiR對初級氮的同化起著非常重要的作用,NR調控硝酸鹽降解為亞硝酸鹽,NiR再進一步將亞硝酸鹽降解為銨,最終銨參與到蛋白質的合成中[34]。本實驗中,對小白菜硝酸鹽含量影響較大的是NR活性。有研究報道隨著營養液硝態氮的增加,蔬菜硝酸鹽含量與NR活力隨之升高[35]。而本實驗中,隨著硝銨比的增加,小白菜葉片NR活力呈下降趨勢。該結果與胡承孝等[36]報道小白菜、番茄的硝酸鹽含量與NR活力間呈負相關相似。NR作為是植物氮代謝中一個重要的限速酶和調節酶,在高硝態氮下其活性降低,可能是造成小白菜體內硝酸鹽積累的重要原因。
NRT在植物體內介導根系對NO3-的吸收以及在不同組織、器官間的轉運。高等植物共有2 個家族的轉運蛋白:NRT1和NRT2,分別與LATS和HATS有關[37]。從實時熒光定量PCR的結果來看,NRT1在‘香港特選奶白菜’和‘揭農四號春白菜’中均顯著表達,而未檢測出NRT2表達量。這與孔敏等[38]的結果不一致。這可能是由于NRT基因在植物組織中各器官中表達有很大的差異,且NRT2在根部表達量要比葉、花等器官中的表達量要高[39]。本實驗中,‘揭農四號春白菜’中的NRT1相對表達量顯著高于‘香港特選奶白菜’,這與‘揭農四號春白菜’和‘香港特選奶白菜’葉片硝酸鹽積累量差異表現一致。此結果與趙首萍等[15]的結果大致相同。說明NRT1的表達在一定程度上可以解釋品種間葉片硝酸鹽積累的差異原因。在本實驗中,由于在硝銨比為100∶0時兩個小白菜葉片中的硝酸鹽含量最高,且葉片是主要食用部分,因此僅對硝銨比為100∶0處理時的‘香港特選奶白菜’和‘揭農四號春白菜’葉片進行了實時熒光定量PCR測定,僅檢測出NRT1的表達量。因此,要從根本上去探究硝酸鹽積累差異的機理,還需要對硝銨比為75∶25、50∶50的小白菜根、葉柄中的NRT1和NRT2的表達進行研究,并且研究NRT1和NRT2家族各成員在白菜各組織器官中發揮的作用對于探究小白菜中硝酸鹽積累的機理有重要意義。
NRT1表達部位與其功能密切相關,通常無機營養的吸收同化需要各種膜結合的離子轉運蛋白,一部分直接參與土壤中離子的吸收,而另一部分則參與離子的轉運和儲存[39]。CHL1基因的蛙卵實驗表明該基因蛋白位于細胞膜上,屬于質子的硝酸鹽依賴共轉運系統[40]。本研究亞細胞定位結果顯示小白菜NRTl定位于細胞膜上,是一個質膜蛋白。說明NRT1發生功能的部位是細胞膜,這與離子轉運蛋白大部分是膜蛋白的特征一致。但是龐永奇等[41]認為目前所進行的亞細胞定位分析大多是穩定過量表達或者瞬時表達產生的結果,在一定程度上可以反映蛋白質的定位情況,但由于表達位置的不同、表達量的變化以及可能存在某些調控因子,可能不能夠真實呈現基因的定位情況,而采用免疫原位雜交的方法或利用自身啟動子驅動基因融合標記的表達可能會得到更準確的結果。因此,對于NRT1在細胞膜上如何發生轉運功能,還需要進一步研究。
4 結 論
供試2 個品種小白菜體內硝酸鹽含量在不同硝銨比下均表現為葉柄>葉片>根。相同硝銨比處理下,‘香港特選奶白菜’葉、葉柄和根中的硝酸鹽含量均明顯低于‘揭農四號春白菜’。
不同硝銨比處理中,除硝銨比50∶50處理下的NiR、GDH活力外,其他氮代謝相關酶活力在兩個小白菜品種之間均達到顯著性差異。‘香港特選奶白菜’和‘揭農四號春白菜’葉片GDH活力在75∶25時均達到最大值。隨著硝銨比的增加,小白菜葉片NR、GOGAT活力呈降低趨勢,NiR活性呈上升趨勢,GDH活性呈先增加后降低趨勢。
低親和轉運蛋白基因NRT1在篩選出的兩個小白菜品種中均顯著表達,且硝酸鹽高富集品種‘揭農四號春白菜’的表達量顯著高于低富集硝酸鹽品種‘香港特選奶白菜’。NRT1是定位于細胞膜上的低親和NRT。
參考文獻:
[1] 趙婉伊, 姚云柯, 徐衛紅, 等. 外源硒對莖瘤芥硒形態及硒吸收的影響[J]. 食品科學, 2017, 38(1): 105-109. DOI:10.7506/spkx1002-6630-201701017.
[2] 趙靜, 王娜, 馮敘橋, 等. 蔬菜中硝酸鹽和亞硝酸鹽檢測方法的研究進展[J]. 食品科學, 2014, 35(8): 42-49. DOI:10.7506/spkx1002-6630-201408008.
[3] 陳云增, 陳志凡, 馬建華, 等. 沙潁河流域典型癌病高發區土壤硝態氮對地下水和蔬菜硝酸鹽積累的影響[J]. 環境科學學報, 2016,36(3): 990-998. DOI:10.13671/j.hjkxxb.2015.0497.
[4] 孫鐘雷, 許藝, 李宇, 等. 離子電極法快速測定榨菜中硝酸鹽含量[J].食品科學, 2016, 37(14): 184-188. DOI:10.7506/spkx1002-6630-201614033.
[5] 都韶婷, 章永松, 林咸永, 等. 蔬菜積累的硝酸鹽及其對人體健康的影響[J]. 中國農業科學, 2007, 40(9): 2007-2014. DOI:10.3321/j.issn:0578-1752.2007.09.021.
[6] MARCO T A, NEIL P B, GIOVANNA S C, et al. Combining social learning with agro-ecological research practice for more effective management of nitrate pollution[J]. Environmental Science and Policy,2007, 10(6): 551-563. DOI:10.1016/j.envsci.2007.02.006.
[7] 都韶婷, 金崇偉, 章永松. 蔬菜硝酸鹽積累現狀及其調控措施研究進展[J]. 中國農業科學, 2010, 43(17): 3580-3589. DOI:10.3864/j.issn.0578-1752.2010.17.012.
[8] 劉永剛, 陳利軍, 武志杰. 蔬菜中硝酸鹽的積累機制及其調控措施[J]. 土壤通報, 2006, 37(3): 612-616. DOI:10.3321/j.issn:0564-3945.2006.03.045.
[9] CHEN Guilin, GAO Xiurui, ZHANG Xianbin. Effect of partial replacement of nitrate by amino acid and urea on nitrate content of non-heading Chinese cabbage and lettuce in hydroponic condition[J].Agricultural Sciences in China, 2002, 1(4): 444-449.
[10] 邢素芝, 汪建飛, 李孝良, 等. 氮肥形態及配比對菠菜生長和安全品質的影響[J]. 植物營養與肥料學報, 2015, 21(2): 527-534.DOI:10.11674/zwyf.2015.0229.
[11] 汪李平, 向長萍, 王運華. 白菜不同基因型硝酸鹽含量差異的研究[J]. 園藝學報, 2004, 31(1): 43-46; 16. DOI:10.3321/j.issn:0513-353X.2004.01.010.
[12] 杜紅艷, 于平彬, 季延海, 等. 不同氮素濃度對水培韭菜生長發育與品質的影響[J]. 中國蔬菜, 2015, 1(5): 45-48. DOI:10.3969/j.issn.1000-6346.2015.05.013.
[13] 都韶婷, 李玲玲, 章永松, 等. 不同基因型小白菜硝酸鹽積累差異及篩選研究[J]. 植物營養與肥料學報, 2008, 14(5): 969-975.DOI:10.3321/j.issn:1008-505X.2008.05.024.
[14] 陳振德, 陳建美, 何金明, 等. 大白菜不同品種硝酸鹽含量的分析[J].中國蔬菜, 2002, 1(2): 40. DOI:10.3969/j.issn.1000-6346.2002.02.019.
[15] 趙首萍, 張瑞麟, 徐明飛, 等. 銨硝比例對不同基因型小白菜硝酸鹽積累影響機理研究[J]. 土壤, 2011, 43(1): 32-38. DOI:10.13758/j.cnki.tr.2011.01.024.
[16] 田真, 李敬蕊, 王祥, 等. 生菜硝酸還原酶基因的克隆及高氮水平下外源γ-氨基丁酸對其表達和葉片硝酸鹽含量的影響[J]. 西北植物學報, 2015, 35(6): 1098-1105. DOI:10.7606/j.issn.1000-4025.2015.06.1098.
[17] 朱美紅, 蔡妙珍, 邢承華, 等. 蔬菜硝酸鹽累積的生理和分子生物學機理[J]. 土壤通報, 2008, 39(3): 694-697. DOI:10.3321/j.issn:0564-3945.2008.03.045.
[18] 王利群, 王文兵, 吳守一, 等. 蔬菜硝酸鹽含量與硝酸還原酶活性的研究[J]. 食品科學, 2003, 24(12): 37-40. DOI:10.3321/j.issn:1002-6630.2003.12.003.
[19] 遲蓀琳, 楊蕓, 徐衛紅, 等. 小白菜硝酸鹽含量與NH4+/NO3-及氮代謝關鍵酶的相關性研究[J]. 食品科學, 2015, 36(23): 70-77.DOI:10.7506/spkx1002-6630-201523014.
[20] 劉娜, 閆志山, 范有君, 等. 不同氮素水平對甜菜氮代謝酶和可溶性蛋白含量的影響[J]. 中國農學通報, 2015(30): 149-154.
[21] 宗學鳳, 王三根. 植物生理研究技術[M]. 重慶: 西南師范大學出版社, 2011: 112-116.
[22] OZAWA K, KAWAHIGASHI H. Positional cloning of the nitrite reductase gene associated with good growth and regeneration ability of calli and establishment of a new selection system for Agrobacterium mediated transformation in rice (Oryza sativa L.)[J]. Plant Science,2006, 170(2): 384-393. DOI:10.1016/j.plantsci.2005.09.015.
[23] 鄒奇. 植物生理學實驗指導[M]. 北京: 中國農業出版社, 2003:125-126.
[24] 葉利庭, 呂華軍, 宋文靜, 等. 不同氮效率水稻生育后期氮代謝酶活性的變化特征[J]. 土壤學報, 2011, 48(1): 132-140.
[25] 楊學東, 戴薇, 張昌偉, 等. 白菜病毒誘導基因沉默技術體系的建立[J].園藝學報, 2012, 39(11): 2168-2174.
[26] 艾紹英, 姚建武, 黃小紅, 等. 蔬菜硝酸鹽的還原轉化特性研究[J].植物營養與肥料學報, 2002, 8(1): 40-43. DOI:10.3321/j.issn:1008-505X.2002.01.007.
[27] ULLRICH W R. Transport of nitrate and ammonium through plant membranes[M]// MENGEL K, PILBEAM D. Nitrogen metabolism of plants. Oxford: Clarendon Press, 1992: 121-137.
[28] 何欣, 張攀偉, 丁傳雨, 等. 弱光下硝銨比對小白菜氮吸收和碳氮分配的影響[J]. 土壤學報, 2009, 46(3): 452-458. DOI:10.3321/j.issn:0564-3929.2009.03.011.
[29] 王朝輝, 田霄鴻, 李生秀. 葉類蔬菜的硝態氮累積及成因研究[J]. 生態學報, 2001, 21(7): 1136-1141. DOI:10.3321/j.issn:1000-0933.2001.07.015.
[30] CORREIA M J, FONSECA F, AZEDO-SILVA J, et al. Effects of water deficit on the activity of nitrate reductase and content of sugars,nitrate and free amino acids in the leaves and roots of sunflower and white lupin plants growing under two nutrient supply regimes[J].Physiologia Plantarum, 2005, 124(1): 61-70. DOI:10.1111/j.1399-3054.2005.00486.x.
[31] LILLO C. Signalling cascades integrating light-enhanced nitrate metabolism[J]. Biochemical Journal, 2008, 415(1): 11-19.DOI:10.1042/BJ20081115.
[32] CHEN Wei, LUO Jinkui, SHEN Qirong. Effect of NH4+-N/NO3--N ratios on growth and some physiological parameters of Chinese cabbage cultivars[J]. Pedosphere, 2005, 15(3): 310-318.
[33] GRANSTECK R C, HUFFAKER R C. Identification of the leaf vacuole as a major nitrate storage pool[J]. Plant Physiology, 1982,70(2): 410-413. DOI:10.1104/pp.70.2.410.
[34] KYAING M S, 顧立江, 程紅梅. 植物中硝酸還原酶和亞硝酸還原酶的作用[J]. 生物技術進展, 2011, 1(3): 159-164. DOI:10.3969/j.issn.2095-2341.2011.03.02.
[35] 李亦松. 氮肥對綠葉類蔬菜硝酸鹽累積的影響及機理的研究[D]. 烏魯木齊: 新疆農業大學, 2006: 1-40. DOI:10.7666/d.y873788.
[36] 胡承孝, 鄧波兒, 劉同仇. 施用氮肥對小白菜、番茄中硝酸鹽積累的影響[J]. 華中農業大學學報, 1992, 11(3): 239-243. DOI:10.13300/j.cnki.hnlkxb.1992.03.007.
[37] 賀瑞, 彭赟, 劉揚, 等. 低氮脅迫下香蕉硝酸鹽轉運蛋白Manrt2基因的克隆與表達分析[J]. 熱帶生物學報, 2016, 7(1): 76-81.DOI:10.15886/j.cnki.rdswxb.2016.01.013.
[38] 孔敏, 楊學東, 侯喜林, 等. 白菜NRT2基因的克隆及表達模式分析[J].園藝學報, 2011, 38(2): 2309-2316. DOI:10.16420/j.issn.0513-353x.2011.12.014.
[39] MARTINOIA E, WIEKEN A. Vacuoles as storage compartments for nitrate in barley leaves[J]. Nature, 1981, 289: 292-294. DOI:10.1038/289292a0.
[40] TSAY Y F, SCHROEDER J I, FELDMANN K A, et al. The herbicide sensitivity gene CHL1 of arabidopsis encodes a nitrate-inducible nitrate transporter[J]. Cell, 1993, 72(5): 705-713. DOI:10.1016/0092-8674(93)90399-B.
[41] 龐永奇, 王高奇, 王旭初, 等. 植物水孔蛋白的亞細胞分布與生理功能研究淺析[J]. 生物化學與生物物理進展, 2012, 39(10): 962-971.DOI:10.3724/SP.J.1206.2011.00617.
