磷鎘交互作用對白骨壤幼苗體內鎘的亞細胞分布和生理特性的影響
2018-05-02 03:12:46戴閩玥嚴重玲
農業環境科學學報 2018年4期
關鍵詞:植物
王 嵐,戴閩玥,嚴重玲*
(1.銅仁學院農林工程與規劃學院,貴州 銅仁 554300;2.廈門大學環境與生態學院,福建 廈門 361102)
鎘(Cd)是毒性很強、移動性大的重金屬之一,廣泛存在于土壤和水體中,容易在生物體內富集,對生物危害很大[1-2]。紅樹林是生長在熱帶、亞熱帶海岸潮間帶的木本植物群落,在維持生態平衡和環境保護方面起著重要作用[3]。隨著工業化的快速發展,大量污水排放,近海水域及紅樹林等面臨著嚴重的重金屬Cd污染問題[4]。有研究人員報道過紅樹林濕地沉積物中Cd 含量范圍為 0.06~0.98 μg·g-1[5],Wu 等[6]研究表明漳江口紅樹林可利用磷為(28.41±1.07)mg·kg-1,可利用氮(97.85±4.67)mg·kg-1。磷(P)是植物生長發育不可缺少的大量元素之一,參與植物體內多種化合物的形成以及代謝調控,施加P肥能降低Cd的有效性[7],緩解Cd對植物的毒害作用[8]。
大量研究表明,細胞壁的固定和液泡區室化在植物應對重金屬的毒性方面起著相當重要的作用[3,9]。Cd在植物體中主要積累在細胞壁和細胞可溶部分,說明細胞壁是植物對Cd解毒的首要屏障,Cd2+進入細胞后主要富集在液泡中,使得其他細胞器免受毒害[10-13]。植物在抗Cd的脅迫中,抗氧化酶的作用不容忽視。Cd導致活性氧(ROS)生成而對植物產生傷害,植物通過抗氧化酶類(如SOD、POD等)逐步形成保護機制來減輕或消除ROS對植物的傷害[14-15]。
白骨壤[Avicennia marina(Forsk.)Vierh]是主要的紅樹植物之一,也是紅樹林植物群落的主要樹種之一,對重金屬污染的耐受性較強。目前,有關重金屬Cd對白骨壤的研究主要集中在Cd對白骨壤脅迫下幼苗的生理生態特性、鉀鈉鎂吸收和分配、Si對Cd脅迫下白骨壤幼苗低分子質量有機酸代謝和Cd脅迫對白骨壤幼苗生理影響等方面[16-17],有關 P、Cd交互作用下白骨壤幼苗不同細胞器對重金屬區域化固定和生理特性的研究尚未見報道。因此,本文選取白骨壤為材料,采用溶液培養,進行Cd和P交互試驗處理,研究不同濃度P對Cd脅迫下白骨壤幼苗體內Cd亞細胞分布、抗氧化酶、葉綠素和蛋白質含量的影響,以說明P存在情況下Cd在白骨壤幼苗體內的分布及部分生理變化,從而緩解Cd的毒性,為紅樹林濕地環境安全與削減重金屬污染提供基礎數據。
1 材料與方法
1.1 試驗材料及處理
白骨壤成熟胚軸采集于福建漳江口紅樹林國家級自然保護區(東經 117°24′,北緯 23°53′),沙培 120 d,培養期間統一管理。待白骨壤幼苗子葉完全枯黃掉落后,先后采用1/4、1/2、全Hoagland營養液依次進行培養直至第3對葉子成熟,調節營養液鹽濃度為1%,pH控制在6.5左右(采用NaOH或HCl溶液調節)。選取長勢一致、規格相同的白骨壤幼苗移栽至口徑15 cm、底內徑10 cm、深20 cm的塑料小桶中,用Hoagland營養液進行適應性培養7 d,然后進行不同濃度的Cd、P交互處理。試驗設定9個處理,即Cd(0、0.5、5 mg·L-1),P(0、30、90 mg·L-1),每個處理重復 3盆,每盆3株,濃度搭配如表1。實驗期間,每3 d更換含Cd和P的營養液,處理30 d后收苗。收苗時白骨壤幼苗根系用Mini-Q超純水沖洗,用濾紙吸干植物體表面水分,根和葉分別用于不同亞細胞組分Cd含量、抗氧化酶活性和蛋白質含量以及對白骨壤葉片中葉綠素的測定。

表1 Cd、P交互處理Table 1 The treatments of Cd and P
1.2 試驗方法
1.2.1 不同亞細胞組分的提取及測定
利用差速離心法提取不同亞細胞組分[18]。分別稱取 0.50 g 根和葉鮮樣,加入 10 mL 50 mmol·L-1Tris-HCl緩沖溶液(pH 7.4)、250 mmol·L-1蔗糖和 1.0 mmol·L-1二硫赤蘚糖醇(DTE)的混合液,研磨成勻漿后轉至離心管,500×g離心10 min,沉淀為細胞壁組分,上清液繼續在10 000×g下離心10 min,沉淀為含葉綠體和細胞核組分,上清液繼續在18 000×g下離心10 min,沉淀為線粒體組分,上清液為可溶性組分。全部操作在4℃下進行。所有組分用10 mL混酸(硝酸∶H2O2=2∶1,V∶V)145 ℃消化 4 h,超純水定容至 40 mL,用0.22 μm濾膜過濾,電感耦合等離子質譜儀ICP-MS(PerkinElmer,UK)測定各亞細胞 Cd 含量。
1.2.2 抗氧化酶活性的測定
參照李合生等[19]的方法測定超氧化物岐化酶(SOD)(氮藍四唑法)和過氧化物酶(POD)活性(愈創木酚法)。
1.2.3 葉綠素含量的測定
參照張雯[9]的方法測定葉綠素含量。去除白骨壤葉片主脈,稱取0.10 g鮮樣,將樣品剪碎后置于帶塞玻璃試管,加入 5 mL 丙酮和乙醇混合液(V∶V=1∶1),避光放置 24 h直至組織完全變白,3000 r·min-1離心10 min,分別在663 nm和645 nm處測定OD值,測定葉綠素a、b含量,并計算總葉綠素和葉綠素a/b。
1.2.4 蛋白質含量的測定
蛋白質含量的測定參照李合生等[19]的考馬斯亮藍G-250法。
1.3 數據分析
采用Excel 2013制作圖表,SPSS 17.0軟件進行方差分析,對同一Cd處理水平下不同P處理組進行單因素方差分析和LSD顯著性檢驗,顯著性水平為0.05。
2 結果與討論
2.1 P、Cd交互處理對白骨壤幼苗體中Cd的亞細胞分布影響
劉芳等[20]研究了P和Cd交互作用對煙草生長及吸收積累P和Cd的影響,研究表明隨著P肥用量增加,高Cd環境下煙草積累Cd量呈上升趨勢,高Cd高P水平時植物增加了對Cd總量的吸收。從表2中可知,Cd 濃度從 0.5 mg·L-1增大到 5 mg·L-1時,白骨壤幼苗體內亞細胞Cd的含量均不同程度地增加,其中細胞壁和可溶性組分中Cd的含量增加較多,可能是由于細胞壁上的纖維素、半纖維素、蛋白質和多糖與Cd結合,從而限制了Cd的轉移[21],研究結果與張雯等[9-10]一致。不同處理條件下,由于Cd是由根部直接進入白骨壤幼苗體內,所以根部Cd的含量明顯多于葉的含量,與張雯、Qiu等[9,21]研究結果相同。本試驗中,各處理中白骨壤幼苗根部Cd的含量明顯多于葉的,可能是由于P的添加增加了HPO-4和H2PO-4等負離子,負離子與Cd2+結合,從而減少Cd2+從根遷移到地上部分,進一步減少Cd在葉中的含量[21-22],從而緩解了Cd對白骨壤的毒害。在相同Cd水平處理下,隨著P濃度的升高,白骨壤葉和根的細胞壁中Cd含量呈上升趨勢,可能是添加的P促進了根系發育,增加了根吸收Cd的含量。當Cd濃度為0.5 mg·L-1時,白骨壤葉和根中可溶性組分的Cd含量先降后升,葉中可溶性組分Cd的含量多于在細胞壁中的,施加適量濃度的 P(30 mg·L-1)能夠促進 Cd 向液泡(可溶性組分)轉移。在高濃度 Cd(5 mg·L-1)處理下,Cd 在可溶性組分中的含量變化不一,具體原因有待進一步研究。總體來說,隨著P濃度的增加,葉和根中可溶性組分Cd的百分比升高,說明液泡也是植物細胞貯存重金屬的重要場所[9]。所以,細胞壁是白骨壤對Cd解毒的首要屏障,Cd2+進入細胞后主要富集在液泡中使得其他細胞器免受毒害[3,10-13],減輕了 Cd 對白骨壤的毒害,進一步說明了細胞壁的固定及液泡區室化是白骨壤對Cd的解毒機制之一。

表2 P對不同Cd處理下白骨壤幼苗體內Cd亞細胞分布的影響Table 2 Subcellular distribution of Cd in A.marina seedlings under different P and Cd concentration
2.2 P、Cd交互處理對白骨壤幼苗體中抗氧化酶活性的影響
Cd脅迫能干擾許多植物的正常生長和代謝,產生ROS,使植物細胞受到傷害[23]。植物清除ROS有很多方式,其中抗氧化酶參與解毒過程是一個非常重要的方式[24]。SOD能將超氧陰離子歧化為H2O2和O2,POD利用H2O2來催化過氧化物的氧化和分解,這是植物應對逆境脅迫的響應機制之一[25]。
從圖1可以看出,單Cd處理時,隨著Cd濃度的增加,白骨壤葉片和根的SOD活性呈上升趨勢,與Kavita等[26]研究結果一致。隨著施Cd及P濃度的增加,各處理葉片和根的SOD活性均有一定的升高。添加90 mg·L-1的P后,白骨壤幼苗葉中的SOD活性增加,與對照相比差異顯著,與Qiu等[27]研究結果一致,具體原因可能是施P促進了白骨壤的生長,為了應對Cd對植株的毒害,葉和根的SOD活性升高,有效清除了由Cd引起的氧化脅迫產生的活性氧。
由圖1還可見,單Cd處理時,隨著Cd濃度的增加,白骨壤葉片和根的POD活性變化趨勢與SOD活性一致。施加相同P水平處理下,隨著施Cd濃度的增加,各組葉片和根的POD活性有一定的升高,與對照相比,各處理間差異不顯著。在高 P(90 mg·L-1)高Cd(5 mg·L-1)處理下,白骨壤葉和根的 POD 活性最高,可能原因是Cd脅迫下促進POD升高,P的添加進一步增加了POD的含量,提高了抗氧化能力,有效清除毒害產生的活性氧。因此,施加一定濃度的P可緩解Cd對白骨壤幼苗的脅迫和傷害。
2.3 P、Cd交互處理對白骨壤幼苗中葉綠素含量的影響
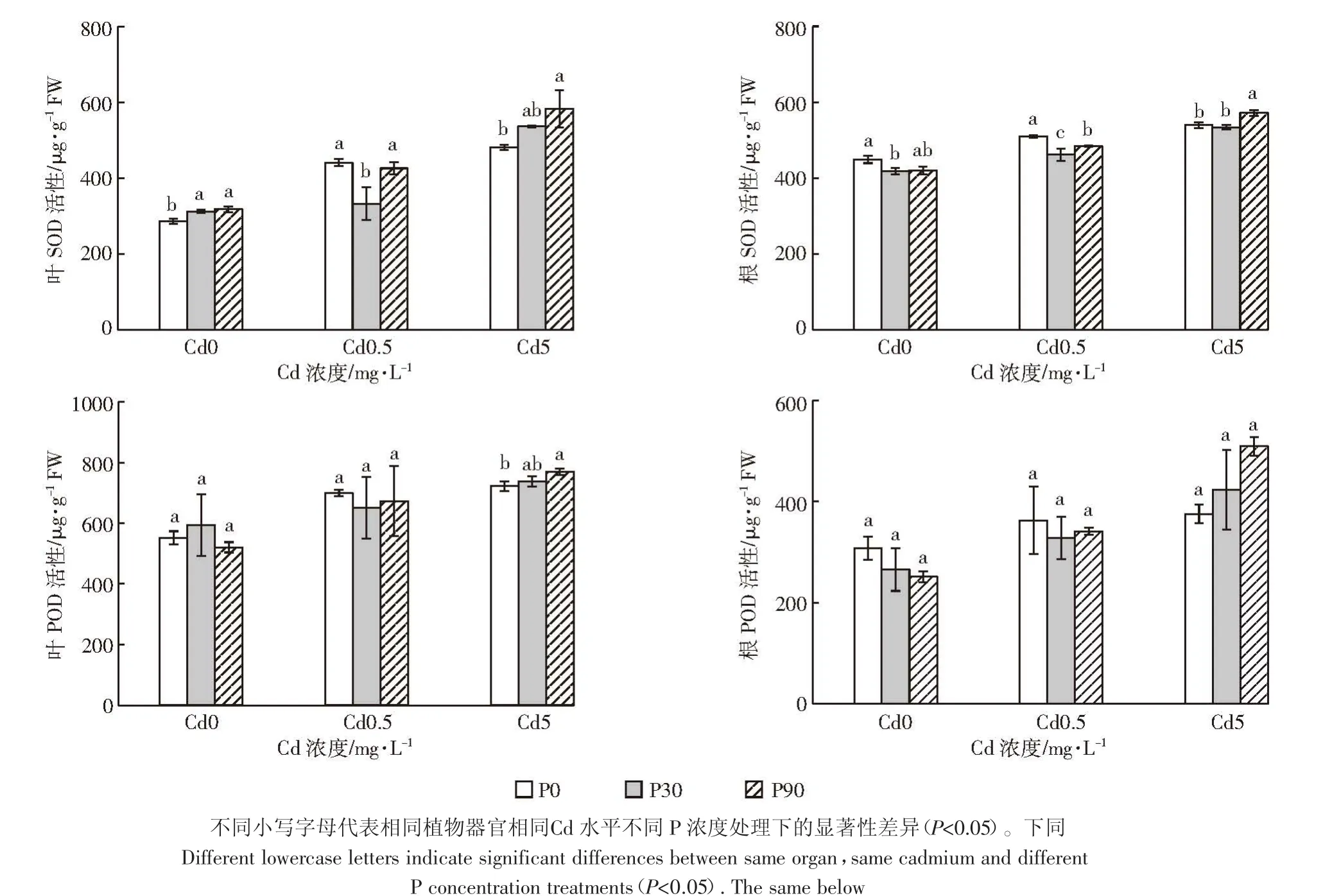
圖1 P、Cd交互處理下白骨壤幼苗抗氧化酶活性Figure 1 Antioxidant enzyme activity of A.marina seedlings under P and Cd treatments

表3 P、Cd交互處理下白骨壤幼苗葉綠素含量及葉綠素a/b變化Table 3 Chlorophyll contents and chlorophyll a/b in A.marina seedlings under P and Cd treatments
從表3中可以看出,隨著Cd濃度的增加,葉綠素含量總體呈下降趨勢,Chla/b上下波動,變化不一。在無Cd處理下,白骨壤幼苗的總葉綠素含量隨著P濃度的增大而增多,P 濃度為 90 mg·L-1時,Chla、Chlb和總葉綠素最大。在施加Cd的處理下,Chla、Chlb和總葉綠素的含量呈下降趨勢,但各處理間差異不顯著。實驗結果說明Cd濃度增加,抑制或破壞了白骨壤幼苗葉綠素的合成,可能是由于Cd抑制葉綠素酸酯還原酶和影響氨基-γ-酮戊二酸的合成,直接破壞了葉綠體結構及功能[28]。當Cd濃度為0.5 mg·L-1和5 mg·L-1時,總葉綠素含量隨著P濃度的增大而有所增多:一方面,P的添加可能在一定程度上降低了Cd的有效性[8],緩解了Cd對葉綠素的破壞,實驗結果與劉芳等[29]研究一致;另一方面可能由于添加的P濃度增加,有效P也隨之增加,葉綠素生物合成過程中δ-氨基-γ-酮戊酸(ALA)合成速率和卟膽原(PBG)酶活性升高,葉綠素生物合成增多[30],這也說明了P能夠緩解Cd對白骨壤幼苗光合系統的破壞。
2.4 P、Cd交互處理對白骨壤幼苗蛋白質含量的影響
植物處于不良環境中,逆境蛋白增加,以增加滲透調節物質而增強植物細胞對逆境的抵抗力[19]。從圖2中可知,當Cd濃度為0.5 mg·L-1時,隨著P濃度增大,白骨壤幼苗葉片中的蛋白質含量呈下降趨勢,各處理間差異不顯著。在根中蛋白質含量先降后升,但各處理間差異也不顯著。當Cd濃度為5 mg·L-1時,隨著添加的P濃度上升,白骨壤葉片中的蛋白質含量也呈上升趨勢,根中的蛋白質含量先降后升,各處理間差異顯著。當P濃度為90 mg·L-1時,葉片和根中的蛋白質含量顯著增加,分析原因可能是P為植物生長發育所需的大量營養元素,P的添加在一定程度上改善了植株的營養狀況,從而促進了根系的發育和植株地上部的生長[31],提高了植物體內蛋白質的含量,進而減輕Cd對植株的毒害,增強白骨壤幼苗對Cd的耐受性。
3 結論
(1)細胞壁的固定及液泡區室化是白骨壤幼苗Cd污染解毒的重要機制之一。
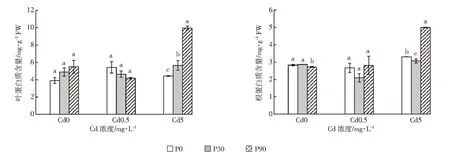
圖2 P、Cd交互處理下白骨壤幼苗蛋白質含量Figure 2 Protein contents of A.marina seedlings under P and Cd treatments
(2)外源P能促使Cd向液泡轉移,同時提升白骨壤植株SOD、POD活性,增加了葉綠素和蛋白質含量。
(3)外源P能夠緩解Cd對紅樹植物白骨壤的影響,增強白骨壤對Cd的耐受能力。
參考文獻:
[1]王曉娟,王文斌,楊 龍,等.重金屬鎘(Cd)在植物體內的轉運途徑及其調控機制[J].生態學報,2015,35(23):7921-7929.WANG Xiao-juan,WANG Wen-bin,YANG Long,et al.Transport pathways of cadmium(Cd)and its regulatory mechanisms in plant[J].E-cologica Sinica,2015,35(23):7921-7929.
[2]程 皓,陳桂珠,葉志鴻.紅樹林重金屬污染生態學研究進展[J].生態學報,2009,29(7):3893-3900.CHENG Hao,CHEN Gui-zhu,YE Zhi-hong.Research progress of heavy metal pollution in mangrove wetlands[J].Acta Ecologica Sinica,2009,29(7):3893-3900.
[3]Weng B S,Xie X Y,Weiss D J,et al.Kandelia obovata(S.,L.) Yong tolerance mechanisms to cadmium:Subcellular distribution,chemical forms and thiol pools[J].Marine Pollution Bulletin,2012,64(11):2453-2460.
[4]黃志亮.Cd低積累蔬菜品種篩選及其Cd積累與生理生化特性研究[D].湖北:華中農業大學,2012.HUANG Zhi-liang.Screening of low Cd-accumulation vegetable cultivars and research on its properties of Cd-accumulation and physiology[D].Hubei:Huazhong Agricultural University,2012.
[5]李柳強.中國紅樹林濕地重金屬污染研究[D].廈門:廈門大學,2008.LI Liu-qiang.Heavy metals in the mangrove wetland of China[D].Xiamen:Xiamen University,2008.
[6]Wu G R,Hong H L,Yan C L.Arsenic accumulation and translocation in mangrove(Aegiceras corniculatum L.)grown in arsenic contaminated soils[J].Public Health,2015,12(7):7244-7253.
[7]劉昭兵,紀雄輝,彭 華,等.磷肥對土壤中Cd的植物有效性影響及其機理[J].應用生態學報,2012,23(6):1585-1590.LIU Zhao-bing,JI Xiong-hui,PENG Hua,et al.Effects of phosphorous fertilizers on phytoavailability of cadmium in its contaminated soil and related mechanisms[J].Chinese Journal of Applied Ecology,2012,23(6):1585-1590.
[8]安志裝,王校常,施衛明,等.重金屬與營養元素交互作用的植物生理效應[J].土壤與環境,2002,11(4):392-396.AN Zhi-zhuang,WANG Xiao-chang,SHI Wei-ming,et al.Plant physiological responses to the interactions between heavy metal and nutrients[J].Soil and Environmental Sciences,2002,11(4):392-396.
[9]張 雯.硫硒交互對水稻幼苗Cd累積和毒害的影響機制研究[D].上海:華東理工大學,2014.ZHANG Wen.Influence mechanism of sulfur and selenium interaction on cadmium accumulation and toxicity in rice seedling[D].Shanghai:East China University of Science and Technology,2014.
[10]張 雯,林匡飛,周 健,等.不同硫濃度下葉面施硒對水稻幼苗鎘的亞細胞分布及化學形態的影響[J].農業環境科學學報,2014,33(5):844-852.ZHANG Wen,LIN Kuang-fei,ZHOU Jian,et al.Effects of selenium foliar spray on subcellular distribution and chemical forms of cadmium in rice seedlings in different sulfur concentrations[J].Journal of Agro-Environment Science,2014,33(5):844-852.
[11]杜遠鵬,李洪敬,尹克林,等.霞多麗苗木中鎘的積累、亞細胞分布及化學存在形態[J].應用生態學報,2012,23(6):1607-1612.DU Yuan-peng,LI Hong-jing,YIN Ke-lin,et al.Cadmium accumulation,subcellular distribution,and chemical forms in Vitis vinifera cv.Chardonnay grapevine[J].Chinese Journal of Applied Ecology,2012,23(6):1607-1612.
[12]Wang X,Liu Y G,Zeng G M,et al.Subcellular distribution and chemical forms of cadmium in Bechmeria nivea(L.)Gaud[J].Environmental and Experimental Botany,2008,62(3):389-395.
[13]Fu X P,Dou C M,Chen Y X,et al.Subcellular distribution and chemical forms of cadmium in Phytolacca americana L.[J].Journal of Hazardous Materials,2011,186(1):103-107.
[14]Blokhina O,Virolainen E,Fagerstedt K V.Antioxidants,oxidative damage and oxygen deprivation stress:A review[J].Annals of Botany,2003,91(2):179-194.
[15]Shi G R,Cai Q S,Liu C F,et al.Silicon alleviates cadmium toxicity in peanut plants in relation to cadmium distribution and stimulation of antioxidative enzymes[J].Plant Growth Regul,2010,61(1):45-52.
[16]陳昌徐,陸志強,鄭文教.白骨壤幼苗對鎘脅迫的生長及生理生態響應[J].廈門大學學報(自然科學版),2015,5(4):485-492.CHEN Chang-xu,LU Zhi-qiang,ZHENG Wen-jiao.The growth and ecophysiological responses of mangrove Avicennia marina seedlings to cadmium[J].Journal of Xiamen University(Natural Science),2015,5(4):485-492.
[17]陸志強,陳昌徐,馬 麗,等.鎘脅迫對白骨壤幼苗鉀鈉鎂吸收與分配的影響[J].應用生態學報,2015,26(5):1313-1319.LU Zhi-qiang,CHEN Chang-xu,MA Li,et al.Absorption and distribution of K,Na and Mg in Avicennia marina seedlings under cadmium stress[J].Chinese Journal of Applied Ecology,2015,26(5):1313-1319.
[18]Wu F B,Dong J,Qian Q Q,et al.Subcellular distribution and chemical form of Cd and Cd-Zn interaction in different barley genotypes[J].Chemosphere,2005,60(10):1437-1446.
[19]李合生,孫 群,趙世杰.植物生理生化試驗原理和技術[M].北京:高等教育出版社,2000.LI He-sheng,SUN Qun,ZHAO Shi-jie.Principles and techniques of plant physiology and biochemistry[M].Beijing:Higher Education Press,2000.
[20]劉 芳,介曉磊,孫巍峰,等.磷、鎘交互作用對煙草生長及吸收積累磷、鎘的影響[J].土壤通報,2007,38(1):116-120.LIU Fang,JIE Xiao-lei,SUN Wei-feng,et al.Influence of P and Cd interaction on their accumulation in tobacco and tobacco growth[J].Chinese Journal of Soil Science,2007,38(1):116-120.
[21]Qiu Q,Wang Y T,Yang Z Y,et al.Effects of phosphorus supplied in soil on subcellular distribution and chemical forms of cadmium in two Chinese flowering cabbage(Brassica parachinensis L.)cultivars differ-ing in cadmium accumulation[J].Food and Chemical Toxicology,2011,49(9):2260-2267.
[22]Bolan N S,Adriano D C,Naidu R.Role of phosphorus in(im)mobilization and bioavailability of heavy metals in the soil-plant system[J].Environ Contam Toxicol,2003,177(2):1-44.
[23]楊衛東,陳益泰.鎘脅迫對旱柳細胞膜透性和抗氧化酶活性的影響[J].西北植物學報,2008,28(11):2263-2269.YANG Wei-dong,CHEN Yi-tai.Membrane leakage and antioxidant enzyme activities in roots and leaves of Salix matsudana with cadmium stress[J].Acta Bot Boreal-Occident Sin,2008,28(11):2263-2269.
[24]Pereira G J G,Molina S M G,Lea P J,et al.Activity of antioxidant enzymes in response to cadmium in Crotalaria juncea[J].Plant and Soil,2002,239(1):123-132.
[25]Azevedo R A,Alas R M,Smith R J,et al.Response of antioxidant enzymes to transfer from elevated carbon dioxide to air and ozone fumigation,in the leaves and roots of wild-type and a catalase-deficient mutant of barley[J].Physiol Plant,1998,104(2):280-292.
[26]Kavita S,Ritambhara G K,Shalini V,et al.Effect of cadmium on lipid peroxidation,superoxide anion generation and activities of antioxidant enzymes in growing rice seedlings[J].Plant Science,2001,161(6):1135-1144.
[27]Qiu R L,Zhao X,Tang Y T,et al.Antioxidative response to Cd in a newly discovered cadmium hyper accumulator,Arabis paniculata F.[J].Chemosphere,2008,74(1):6-12.
[28]陶毅明,陳燕珍,梁士楚,等.鎘脅迫下紅樹植物木欖幼苗的生理生化特性[J].生態學雜志,2008,27(5):762-766.TAO Yi-ming,CHEN Yan-zhen,LIANG Shi-chu,et al.Physiological and biochemical properties of Bruguiera gymnorrhiza seedlings under cadmium stress[J].Chinese Journal of Ecology,2008,27(5):762-766.
[29]劉 芳,介曉磊,劉世亮,等.褐土中磷鎘交互作用對磷鎘有效性影響[J].生態環境,2007,16(5):1429-1432.LIU Fang,JIE Xiao-lei,LIU Shi-liang,et al.Effect of P and Cd interaction on bioavailability of P and Cd in cinnamon soil[J].Ecology and Environment,2007,16(5):1429-1432.
[30]吳 楚,王政權,孫海龍,等.氮磷供給對長白落葉松葉綠素合成、葉綠素熒光和光合速率的影響[J].林業科學,2005,41(4):31-36.WU Chu,WANG Zheng-quan,SUN Hai-long,et al.Effects of different concentrations of nitrogen and phosphorus on chlorophyll biosynthesis,chlorophyll a fluorescence,and photosynthetic rate in Larix olgensis seedlings[J].Scientia Silvae Sinicae,2005,41(4):31-36.
[31]劉世亮,劉忠珍,介曉磊.施磷肥對Cd污染土壤中油麥菜生長及吸收重金屬的影響[J].河南農業大學學報,2005,39(1):30-34.LIU Shi-liang,LIU Zhong-zhen,JIE Xiao-lei.Influence of phosphate fertilizer application on lettuce plant growth and absorbing of heavy metals in Cd polluted soil[J].Journal of Henan Agricultural University,2005,39(1):30-34.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
