秀珍菇單孢子菌株原生質體制備條件的優化
2018-05-07 08:32:57孫曉瑞陳博文張小林孟俊龍
生物技術通報 2018年4期
孫曉瑞 陳博文 張小林 孟俊龍,3
(1. 山西農業大學食品科學與工程學院,太谷 030801;2. 山西農業大學園藝學院,太谷 030801;3. 黃土高原食用菌提質增效協同創新中心,太谷 030801)
秀珍菇(Pleurotus geesteraniSinger)是一種可食用的大型真菌,又被稱為印度鮑魚菇、環柄香菇、姬平菇。隸屬于真菌門,擔子菌綱,傘菌目,側耳科,側耳屬,產于印度南部查摩省,是熱帶和亞熱帶地區蕈菌[1]。1974年,由菌物學家Jandiaik馴化成功,經分離并人工栽培,秀珍菇被證實是一株高產且性狀優良的食用真菌。為更好利用這一珍稀菌物資源,眾多研究者已開展了對其優良菌株的選育工作。與常規育種相比,原生質體技術可以實現遠緣雜交、擴大遺傳物質的重組范圍以及基因轉化和定向誘變,獲得有突出優良性狀的新型菌株,產生更豐富的遺傳變異[2]。原生質體因去掉了細胞壁的障礙而對外界刺激的敏感度顯著提高,是誘變的良好材料[3]。原生質體制備[4]是原生質體技術育種的基礎和重要前提,如何提高原生質體產量及同時保證其較高再生率是此項技術的關鍵研究點。
秀珍菇中含有的秀珍菇多糖具有抗氧化[5]、防衰老[6]、抗腫瘤等功效,目前國內研究多集中于秀珍菇多糖提取[7]和純化[8]方法的優化,以及秀珍菇多糖的結構[9]和生物活性[10]。利用原生質體育種[11]進行突變、雜交、基因轉導等操作,成為獲取秀珍菇多糖高產菌株的一種有效方法。有關秀珍菇原生質體制備的研究較少,且秀珍菇單孢子菌株原生質體制備條件的優化與再生的研究未見報道。本研究從影響酶促反應的因素(菌齡、穩滲液種類及濃度、酶濃度、酶解溫度、酶解時間)出發,系統地對影響原生質體制備的因素進行了研究,以期獲得原生質體制備和再生的最優條件,為利用原生質體融合及基因組改組等選育更為優良的菌株提供參考。
1 材料與方法
1.1 材料
1.1.1 供試菌株 秀珍菇188由山西農業大學食用菌中心提供。
1.1.2 試劑 溶壁酶(廣東省微生物研究所),Tris-HCl(Boster),其他試劑均為國產分析純。
1.1.3 培養基 秀珍菇188單孢菌株生長培養基PDA:馬鈴薯200 g,葡萄糖20 g,瓊脂20 g,用水定容至1 L,高壓滅菌。秀珍菇188單孢菌株培養基PDB:馬鈴薯200 g,葡萄糖20 g,用水定容至1 L,高壓滅菌。原生質體液體再生培養基:PDB添加0.6 mol/L甘露醇和10 mmol/L Tris-HCl。原生質體固體再生培養基:PDA添加0.6 mol/L甘露醇和10 mmol/L Tris-HCl。
1.1.4 穩滲液制備 甘露醇穩滲液:稱取10.9 g甘露醇溶解于100 mL蒸餾水中,制備成0.6 mol/L的甘露醇穩滲液,滅菌備用。稱取7.2 g MgSO4溶解于100 mL蒸餾水中,制備成0.6 mol/L的MgSO4穩滲液滅菌備用。稱取20.54 g蔗糖溶解于100 mL蒸餾水中,制備成0.6 mol/L的蔗糖穩滲液滅菌備用。稱取3.51 g NaCl溶解100 mL蒸餾水中,制備成0.6 mol/L的NaCl穩滲液滅菌備用。稱取4.5 g KCl溶解于100 mL蒸餾水中,制備成0.6 mol/L的KCl穩滲液滅菌備用。1.1.5 酶解液制備 按試驗設計的濃度要求精確稱取溶壁酶溶解于滅菌的穩滲液中,制備成酶解液,過濾除菌備用。
1.2 方法
1.2.1 秀珍菇188單孢菌株獲取 秀珍菇188栽培種培養基配方:棉籽殼81%、麩皮15%、過磷酸鈣1%、石膏粉1%、石灰1%、葡萄糖1%。秀珍菇188栽培種在22℃培養箱中發菌,待菌絲滿瓶后置于25℃,65%濕度條件的人工氣候培養箱中出菇。采收菇體健壯的秀珍菇子實體,剪去菌柄,將子實體菌褶朝下放置于鋪有無菌硫酸紙的無菌平板內。12 h以后在硫酸紙上可見白色單孢子印,用無菌刀片刮取單孢子于無菌試管中,稀釋涂布于PDA平板上,25℃培養3-4 d后,于顯微鏡下顯微操作挑出單個孢子菌絲體,接種于PDA試管中培養。單孢菌絲約一周左右長滿滿管(15 mm×150 mm),菌絲呈放射狀生長,菌絲體整齊均勻潔白。挑選出優良單孢菌絲菌株供原生質體制備與再生使用。
1.2.2 原生質體的制備與再生 將秀珍菇188單孢菌株接種于加有玻璃紙的PDA培養基上,25℃培養。收集菌絲于已知重量的離心管中,稱重,用無菌滲透壓穩定劑沖洗2-3次,4 000 r/min離心15 min,除去上液,得到純凈的菌絲,備用。將收集的菌絲置于5 mL離心管中,按菌絲:酶解液=1 g∶5 mL的比例加入酶解液,置于水浴鍋中試驗設計溫度下水浴酶解,隔10 min振蕩一次,按照試驗設計時間酶解結束后,取0.1 mL原生質體稀釋適當倍數后,血球計數板計數,重復3次。將酶解液用5層擦鏡紙過濾,按試驗設計的酶解時間加入8倍體積的穩滲液終止酶解后制備成原生質懸浮液,原生質體懸浮液4 000 r/min離心10 min后去除上清液,沉淀用滲透壓穩滲液稀釋再離心,重復3次后用滲透壓穩定劑重懸得到較純的原生質體懸液。將上述得到的原生質體懸液取等體積2份,一份用滲透壓穩定劑稀釋至1×105CFU/mL,取1 mL加入到9 mL含有滲透壓穩定劑的液體PDA培養基中,25℃,100 r/min搖床中恢復再生培養24 h,每個PDA平板取0.1 mL液體菌液均勻涂布,10 mL共涂布100個平行(每個平板上獲得的菌落數的均值記為A),另一份加入與上述稀釋原生質體所用的滲透壓穩定劑體積相同的無菌水作為對照,同樣條件下液體恢復再生培養24 h后,每個PDA平板取0.1 mL液體菌液均勻涂布,10 mL共涂布100個平行(每個平板上獲得的菌落數的均值記為B)。原生質體再生率=[(A-B)/C]×100%,選取未染菌的10個最優平板計數。C:每個平板上涂布的原生質體數量(0.1×0.1 mL×105CFU/mL=103CFU)。平板置于25℃恒溫培養箱中培養。
1.2.3 顯微觀察 制片,于顯微鏡100×油鏡下觀察秀珍菇188菌株與其單孢菌株的顯微形態。取少量原生質體懸液于凹玻片中,顯微鏡100×油鏡和40×物鏡下觀察原生質體釋放過程并拍照。
1.2.4 秀珍菇單孢子菌絲體最佳原生質體制備和再生單因素試驗
1.2.4.1 菌絲體菌齡對原生質體制備及再生影響 菌絲體菌齡設定為:4 d、5 d、6 d、7 d、8 d、9 d、10 d,在酶解液濃度為1.6%,酶解溫度29℃,酶解時間為160 min的條件下,以0.6 mol/L甘露醇和10 mmol/L Tris-HCl為穩滲液酶解制備原生質體懸浮液并鏡檢計數,以原生質體釋放量大和原生質體再生率高為優選指標,篩選優化的菌絲體菌齡。
1.2.4.2 酶解液濃度對原生質體制備及再生影響 酶解液濃度設定為:0.4%、0.8%、1.2%、1.6%、2.0%、2.4%和3.0%,以菌齡為8 d的秀珍菇188單孢子菌絲體為酶解對象,在29℃下,酶解時間為160 min的條件下,以0.6 mol/L甘露醇和10 mmol/L Tris-HCl為穩滲液酶解制備原生質體懸浮液并鏡檢計數,以原生質體釋放量大和原生質體再生率高為優選指標,篩選優化的酶解液濃度。
1.2.4.3 酶解溫度對原生質體制備及再生影響 酶解溫度設定為:23、25、27、29、31、33和35℃,以菌齡為8 d的秀珍菇188單孢子菌絲體為酶解對象,在酶解液濃度為1.6%,酶解時間為160 min的條件下,以0.6 mol/L甘露醇和10 mmol/L Tris-HCl為穩滲液酶解制備原生質體懸浮液并鏡檢計數,以原生質體釋放量大和原生質體再生率高為優選指標,篩選優化的酶解溫度。
1.2.4.4 酶解時間對原生質體制備及再生影響 酶解時間設定為:40、80、120、160、200、240和280 min,以菌齡為8 d的秀珍菇188單孢子菌絲體為酶解對象,在酶解液濃度為1.6%,酶解溫度29℃的條件下,以0.6 mol/L甘露醇和10 mmol/L Tris-HCl為穩滲液酶解制備原生質體懸浮液并鏡檢計數,以原生質體釋放量大和原生質體再生率高為優選指標,篩選優化的酶解時間。
1.2.4.5 穩滲液種類對原生質體制備及再生影響 穩滲液種類為:0.6 mol/L的甘露醇、MgSO4、蔗糖、NaCl、KCl穩滲液,以菌齡為8 d的秀珍菇188單孢子菌絲體為酶解對象,在酶解液濃度1.6%,酶解溫度29℃,酶解時間為160 min的條件下制備原生質體懸浮液并鏡檢計數,以原生質體釋放量大和原生質體再生率高為優選指標,篩選優化的穩滲液。
1.2.4.6 優選穩滲液濃度以及Tris-HCl的添加對原生質體制備及再生影響 穩滲液濃度為:0.2 mol/L、0.4 mol/L、0.6 mol/L、0.8 mol/L、1.0 mol/L、0.6 mol/L+T、0.8 mol/L+T,以菌齡為8 d的秀珍菇188單孢子菌絲體為酶解對象,在酶解液濃度1.6%,酶解溫度29℃,酶解時間為160 min的條件下制備原生質體懸浮液并鏡檢計數,以原生質體釋放量大和原生質體再生率高為優選指標,進行穩滲液濃度以及Tris-HCl添加的優化篩選。
1.2.5 秀珍菇單孢子菌絲體最佳原生質體制備和再生正交試驗設計 根據單因素試驗結果,表1設定不同因素和水平,進行L9(33)正交設計。在優化的菌齡和穩滲液條件下,制備原生質體懸浮液,進行L9(33)正交設計,以原生質體釋放量和原生質體再生率為指標,篩選優化的酶濃度、酶解溫度、酶解時間。

表1 秀珍菇單孢子菌絲體最佳原生質體制備和再生L9(33)正交試驗因素水平
2 結果
2.1 顯微觀察
由圖1可見,秀珍菇188單孢菌株菌絲體較秀珍菇188菌株菌絲體纖細,秀珍菇188菌株菌絲體有明顯的鎖狀聯合,秀珍菇188單孢菌株菌絲無鎖狀聯合。

圖1 秀珍菇188菌株與其單孢菌株顯微觀察
秀珍菇188菌株單孢菌株細胞壁被酶分解后,原生質體的釋放有多種形式,且釋放時間不同步。在40×物鏡和100×油鏡下可以清楚的看到原生質體逐漸從菌絲尖端(圖2-A和2-B)釋放出來,在100×油鏡下觀察到原生質體從菌絲體側面(圖2-C)釋放出來,原生質體呈球體狀,大小差異較大。在40×物鏡下觀察到,部分原生質體內含大液泡(圖2-D)。原生質體經4-6層無菌擦鏡紙過濾純化后,菌絲殘渣被完全過濾掉,40×鏡下可觀察到大量大小不一的原生質體(圖2-E)。

圖2 秀珍菇188菌株單孢菌株原生質體釋放過程與方式的觀察
2.2 菌絲體菌齡對原生質體制備及再生影響
菌齡為4-5 d的單孢子菌絲體過于幼嫩很難獲得原生質體,而且獲得的原生質體容易降解;而生長8-10 d的單孢子菌絲體由于生長時間過長,過厚的菌壁使得溶壁酶難以裂解得到大量原生質體;取生長6-8 d的秀珍菇188單孢子菌絲體作為酶解對象可以成功裂解獲得量大質優的原生質體,菌齡為8 d時獲得的原生質體數量和再生率都最好(圖3)。
2.3 酶解液濃度對原生質體制備及再生影響

圖3 菌絲體菌齡對原生質體制備及再生影響
酶濃度從0.4%增大到1.2%時,原生質體產量迅速增加。酶濃度由1.2%增加到1.6%時,原生質體產量增加速度趨于緩慢(圖4),原因是酶濃度逐漸達到飽和。隨著溶壁酶濃度繼續提升,原生質體產量開始降低,原生質體的完整性逐漸被破壞,伴隨著原生質體的活力下降,原生質體再生率也下降。所以,最佳酶濃度應控制在1.6%。
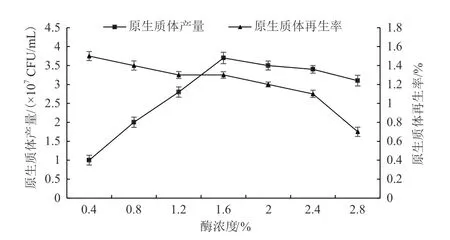
圖4 酶解液濃度對原生質體制備及再生影響
2.4 酶解溫度對原生質體制備及再生影響
隨著酶解溫度的提升,溶壁酶活力提高,原生質體產量也逐步增加,在酶解溫度為29℃時,秀珍菇188單孢子菌絲體原生質體產量達到最大且原生質體再生率也較高(圖5)。
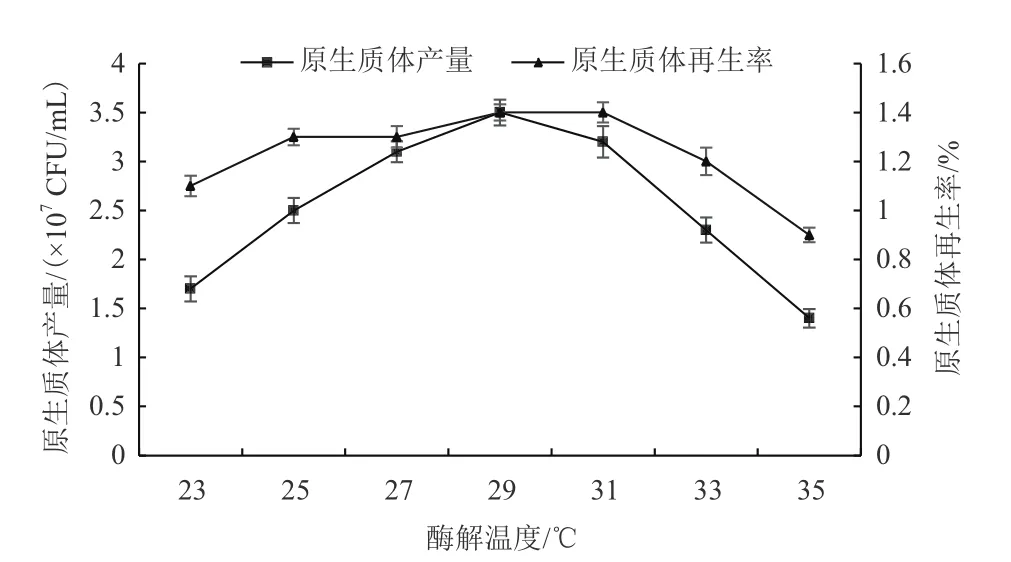
圖5 酶解溫度對原生質體制備及再生影響
2.5 酶解時間對原生質體制備及再生影響
酶解80-120 min后,由于未被酶解的菌絲體較多,釋放出的原生質體較少。酶解240-280 min后得到的原生質體有酶解過度破裂現象,原生質體再生率降低;酶解300 min后,原生質體開始被嚴重破壞,接著酶解,原生質體全部被酶解為碎片殘渣;酶解時間為120-200 min時,可獲得量大質優的秀珍菇188單孢菌株原生質體(圖6)。
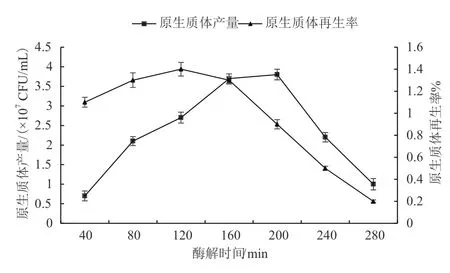
圖6 酶解時間對原生質體制備及再生影響
在原生質體制備過程中,菌絲在酶解作用下,細胞壁不斷被分解形成單個的原生質體。在一定范圍內無論酶解時間長短,原生質體的獲得量均在原生質體制備量最大值左右波動變化,繼續酶解最終會導致細胞壁被完全酶解破壞,過長的酶解時間使得原生質體的再生率下降,原生質體由于無法合成新的細胞壁而不能再生。考慮到節約實驗操作時間以減少雜菌污染,優化選擇的酶解時間為160 min。
2.6 穩滲液種類對原生質體制備及再生影響
研究(圖7)發現,在不加其他成分的PDB中,秀珍菇188單孢子菌絲體原生質體無法再生成功,PDB中添加甘露醇、硫酸鎂、氯化鈉作為滲透壓穩定劑時可以再生成功。穩滲液的添加以甘露醇效果最好,可以很好地保護秀珍菇188、單孢子菌絲體原生質體的完好性。
2.7 優選穩滲液濃度以及Tris-HCl的添加對原生質體制備及再生影響
優選穩滲液甘露醇的濃度和Tris-HCl的添加對秀珍菇188單孢子菌絲體原生質體形成及再生有很大影響,特別是0.6 mol/L的甘露醇和10 mmol/L的Tris-HCl混合穩滲液對于秀珍菇188單孢子菌絲體原生質的獲得及再生效果明顯(圖8)。
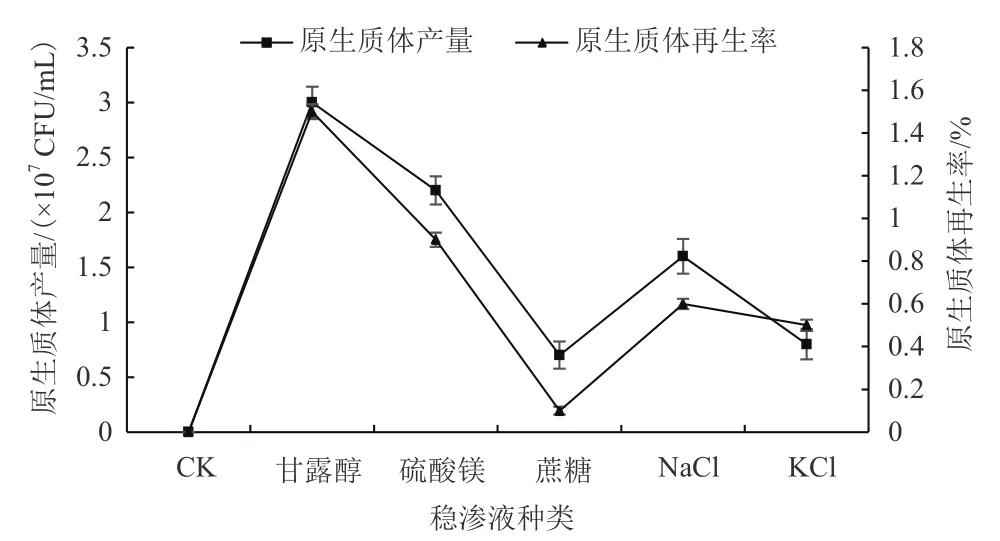
圖7 穩滲液種類對原生質體制備及再生影響
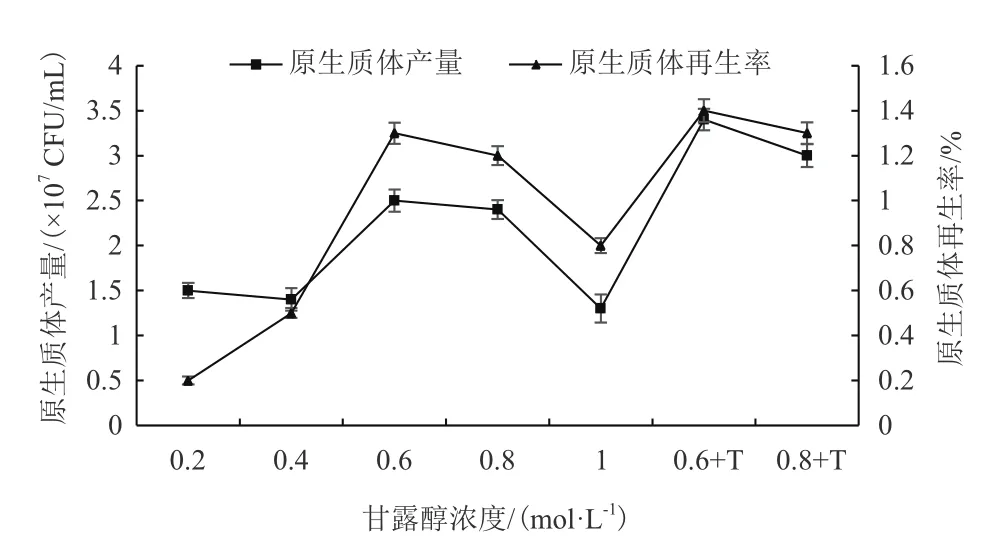
圖8 優選穩滲液濃度以及Tris-HCl的添加對原生質體制備及再生影響
2.8 正交試驗
原生質制備的酶濃度、酶解溫度、酶解時間正交試驗結果見表2。以原生質體產量為評價指標,對結果進行極差分析可知各因素對原生質體制備的影響從大到小的順序為:酶濃度(A)>酶解溫度(B)>酶解時間(C),最佳方案應為A2B1C3,此條件下原生質體產量達到最大值3.5×107CFU/mL。以原生質體再生率為評價指標,對結果進行極差分析可知各因素對原生質體制備的影響從大到小的順序為:酶濃度(A)=酶解時間(C)>酶解溫度(B),最佳方案應為A2B2C2,此條件下原生質體再生率達到最大值1.5%。
本試驗研究中要同時考慮到原生質體的制備量和原生質體的再生率,所以以原生質體產量和原生質體再生率的乘積數值為最終評價指標,對結果進行極差分析可知各因素對原生質體制備和再生的影響從大到小的順序為:酶濃度(A)>酶解時間(C)>酶解溫度(B),最終最佳方案為A2B2C2,此條件下原生質體產量和原生質體再生率的乘積數值達到最大值51。綜合以上評價指標分析,酶濃度對原生質體制備和再生影響最大。最終得到最佳秀珍菇單孢菌絲體原生質體制備和再生指標:酶濃度1.6%,酶解時間160 min,酶解溫度29℃。
2.9 驗證試驗
按照正交試驗優化結果選取最佳原生質體制備和再生條件:在菌齡為8 d、穩滲液為0.6 mol/L甘露醇和10 mmol/LTris-HCl的混合穩滲液,酶濃度1.6%,酶解時間160 min,酶解溫度29℃條件下進行驗證試驗。在此條件下秀珍菇188單孢子菌絲體原生質體釋放量為3.5×107CFU/mL,原生質體再生率為1.4%,原生質體釋放量與原生質體再生率乘積數值為4.9,與正交結果相近,證明本試驗結論可靠。
3 討論
在育種試驗中,當需要獲取親本菌株的單核原生質體菌株時,通用方法為制備親本原生質體,然后由于在原生質體制備過程中,會有部分原生質體失去一個細胞核變成單核原生質體。但是在后期的試驗操作流程中,需用常規方法檢測是否有鎖狀聯合來區分單雙核原生質體菌株,試驗流程較為緩慢。針對那些可以產孢子的菌株,就可以先獲取其孢子,然后制備單核原生質體菌株,可以保證獲取到的原生質體菌株均為單核。而對于那些不產孢子的菌株,或者是野生條件下難以采集孢子的野生菌株[12],可以選用原生質體單核化技術獲取其單核菌株。
對于不同種屬的食用菌,細胞壁化學組成不盡相同,因此在制備原生質體時所需酶也會有差異[13]。許多研究者經過大量的試驗發現,溶壁酶對側耳屬的菌種酶解效果相對較好。因此本研究采用單一酶溶壁酶[14]來制備原生質體。相比較其他文獻中單一考慮原生質體的產量,本研究在單因素試驗和正交試驗中同時考慮到原生質體的產量和再生率[15],為與秀珍菇188單孢子原生質體相關試驗中原生質體的實際操作使用提供了數據支持,使得研究結果更符合試驗實際操作的要求。
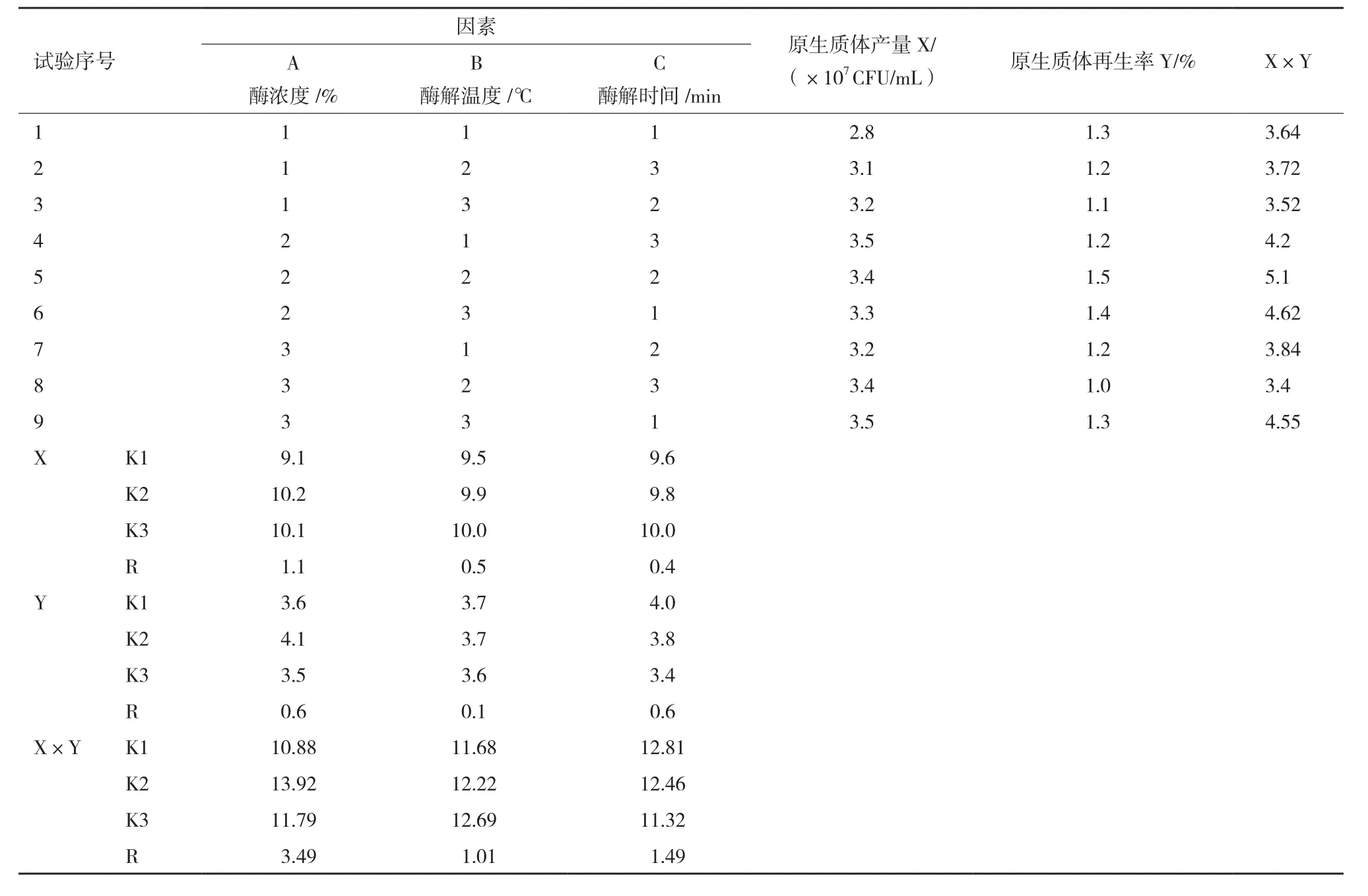
表2 秀珍菇單孢子菌絲體最佳原生質體制備和再生L9(33)正交試驗設計及結果
在原生質體純化上,采用5層擦鏡紙進行純化過濾。相比無菌脫脂棉過濾法[16],原生質體數量下降幅度較無菌脫脂棉過濾法小,操作更為簡單,也降低了染菌率。原生質體的得率是原生質體再生的前提條件,本研究選取加有玻璃紙[17]的PDA作為菌絲生長培養基,菌絲的收集更加方便快捷,縮短了菌絲收集流程,大幅度降低染菌幾率。綜合多個文獻資料,選擇菌絲體清洗的離心轉速為4 000 r/min,在此轉速條件下[18]收集到的原生質體量大且活性高。在原生質體制備和再生過程中,沒有對pH進行調節,保持pH自然,避免影響到原生質體外膜,進而影響原生質體再生率。
4 結論
本研究以原生質體產量,原生質體再生率以及兩者乘積數值為評價指標,采用單因素試驗和正交試驗對其原生質體制備條件進行優化,最終確定了原生質體制備的最佳條件:在菌齡為8 天、穩滲液為0.6 mol/L甘露醇和10 mmol/LTris-HCl的混合穩滲液,酶濃度1.6%,酶解時間160 min,酶解溫度29℃。在此條件下,原生質體產量達到3.5×107CFU/mL,原生質體再生率達到1.4%,兩者乘積數值為4.9。
[1] 袁菁藝. 利用甘草渣栽培秀珍菇和姬菇的研究[D]. 廣州:華南農業大學, 2016.
[2] 常明昌. 食用菌栽培學[M]. 北京:中國農業出版社, 2003.
[3] 朱蘊蘭, 陳安徽, 王陶, 等. 冬蟲夏草原生質體誘變育種研究[J]. 食品科學 , 2010, 31(5):256-260.
[4] Li M, Yi P, Liu Q, et al. Biodegradation of benzoate by protoplast fusant via intergeneric protoplast fusion betweenPseudomonas putidaandBacillus subtili[J]. International Biodeterioration &Biodegradation, 2013, 85(7):577-582.
[5] 孫玉軍, 江昌俊, 祝嫦巍, 等. 秀珍菇多糖PMP-2a的理化性質及其體外抗氧化活性研究[J]. 現代食品科技, 2014, 30(12):79-84.
[6] 孫玉軍, 江昌俊, 任四海. 秀珍菇多糖對D-半乳糖致衰老小鼠的保護作用[J]. 食品科學, 2017, 38(5):251-256.
[7] 耿中華, 李超. 秀珍菇多糖的超聲波協同微波提取工藝及其抗氧化活性研究[J]. 食品工業, 2014, 35(9):110-114.
[8] 許媚. 秀珍菇子實體多糖的分離純化、結構鑒定和生物活性研究[D]. 杭州:浙江工業大學, 2013.
[9] 張安強, 許媚, 周賽靜, 等. 秀珍菇子實體多糖中3-O-甲基-半乳糖組分的譜學鑒定[J]. 藥物分析雜志, 2013, 33(01):90-93, 115.
[10] Silveira MLL, Smiderle FR, et al. Exopolysaccharide produced byPleurotus sajor-caju:Its chemical structure and anti-inflammatory activity[J]. Int J Biol Macromol, 2015, 75(1):90-96.
[11] 楊建遠, 李靜, 范亞葦, 等. 粗壯脈紋孢菌原生質體的制備、再生及轉化的條件[J]. 食品科學, 2015, 36(17):169-172.
[12] 孫勇, 曹小迎, 陳永強, 等. 樺褐孔菌原生質體制備與再生[J].食品科學, 2011, 32(16):161-164.
[13] 高躍, 袁萍, 茅仁剛, 等. 云芝菌絲體原生質體制備與再生條件的研究[J]. 食用菌學報, 2008(1):65-68.
[14] 李艷麗, 金周雨, 李玉. 刺芹側耳原生質體酶解條件的優化[J]. 食品科學 , 2010, 31(10):155-158.
[15] 彭軼楠, 王沛雅, 鞏曉芳, 等. 生防真菌寡雄腐霉原生質體的制備及再生[J]. 菌物學報, 2017, 36(6):679-690.
[16] 張鵬, 龔玲鳳, 朱堅, 等. 刺芹側耳與秀珍菇細胞融合及融合子的鑒定[J]. 食用菌學報, 2013, 20(3):1-5.
[17] 李江, 袁月祥, 閆志英, 等. 毛栓菌原生質體制備和再生及單核菌株產漆酶特性[J]. 菌物學報, 2012, 31(1):102-109.
[18] 周明明, 李曉雁, 任夢楠, 等. 韋蘭膠合成菌的原生質體制備與再生研究[J]. 生物技術通報, 2016(7):126-130.
