高溫脅迫對(duì)棗苗生理特性的影響
2018-05-08 06:14:01樊丁宇
新疆農(nóng)業(yè)科學(xué) 2018年3期
靳 娟,楊 磊,樊丁宇,郝 慶
(新疆農(nóng)業(yè)科學(xué)院園藝作物研究所/農(nóng)業(yè)部新疆地區(qū)果樹(shù)科學(xué)觀測(cè)實(shí)驗(yàn)站,烏魯木齊 830091)
0 引 言
【研究意義】棗(ZiziphusjujubaMill.)為鼠李科(Rhamnaceae)棗屬(ZizyphusMill)植物,是中國(guó)最早栽培的果樹(shù)之一,也是我國(guó)的第一大干果樹(shù)種[1-2]。新疆是棗的主產(chǎn)區(qū)之一,2015年底新疆紅棗種植面積約為4.95×105hm2,總產(chǎn)量約為3.05×106t,棗產(chǎn)業(yè)為帶動(dòng)新疆經(jīng)濟(jì)的發(fā)展起著重要作用[3]。【前人研究進(jìn)展】40℃高溫造成‘翠冠’梨和‘園黃’梨葉綠體、線粒體等細(xì)胞器數(shù)量、形態(tài)和結(jié)構(gòu)發(fā)生顯著變化[4],甚至改變葡萄[5-6]、獼猴桃[7]等果樹(shù)葉片的氣孔特征,降低灰棗[8]、砂梨[9]等果樹(shù)的光合速率。高溫還會(huì)造成梨[10]、棗[11]、葡萄[12]葉片細(xì)胞膜透性增大,膜脂過(guò)氧化加劇。此外,高溫脅迫下活性氧的積累促使獼猴桃[13]、蘋(píng)果[14]、葡萄[15]等果樹(shù)的SOD、POD和CAT等抗氧化酶活性增強(qiáng),以減輕高溫脅迫的傷害。【本研究切入點(diǎn)】近年來(lái)新疆頻繁出現(xiàn)超過(guò)40℃的短期極端高溫天氣[16],對(duì)棗樹(shù)的危害嚴(yán)重,表現(xiàn)為大面積“焦花”、落花落果、果面灼傷、產(chǎn)量和品質(zhì)下降等現(xiàn)象。研究高溫脅迫對(duì)棗葉片生理生化指標(biāo)的影響。【擬解決的關(guān)鍵問(wèn)題】以酸棗和灰棗穴盤(pán)苗為材料,測(cè)定棗葉片在高溫脅迫下的細(xì)胞膜透性、丙二醛(MDA)含量、滲透調(diào)節(jié)物質(zhì)、抗氧化酶活性以及光合色素含量的變化,研究棗實(shí)生苗對(duì)高溫脅迫的適應(yīng)機(jī)制,為制定防御高溫脅迫的策略提供理論參考。
1 材料與方法
1.1 材 料
挑選健康飽滿、大小一致的酸棗和灰棗種子,室溫清水浸泡24 h促進(jìn)種子發(fā)芽。種子催芽后播種于基質(zhì)中培養(yǎng)。待幼苗十四葉一心時(shí),于2017年5月30日挑選長(zhǎng)勢(shì)基本一致的植株置于人工智能氣候培養(yǎng)箱(上海一恒科學(xué)儀器有限公司制造),培養(yǎng)箱條件設(shè)置:晝/夜為14 h/10 h,溫度為30℃/20℃,相對(duì)濕度為80%左右,光合有效輻射為100 μmol/(m2·s)。
于2017年6月6日選擇生長(zhǎng)一致的植株進(jìn)行高溫處理,高溫脅迫期間濕度保持在80%左右,處理溫度為45℃/35℃,光暗周期為14 h/ 10 h。材料在人工培養(yǎng)箱內(nèi)隨機(jī)排列,高溫脅迫期間每天20:00澆1次水,保證水分充足,并按照順序變換位置以保證生長(zhǎng)條件一致。于高溫處理0、1、3和5 d采集葉片,取樣時(shí)間定為16:00,3次重復(fù),葉片經(jīng)液氮速凍后保存于-80℃冰箱中。
1.2 方 法
1.2.1 相對(duì)電導(dǎo)率和MDA含量測(cè)定
葉片相對(duì)電導(dǎo)率參考李合生[17]的方法,使用DDS-307A型電導(dǎo)率儀測(cè)定。MDA含量參照Herth和Paker[18]的方法測(cè)定。
1.2.2 滲透調(diào)節(jié)物質(zhì)含量測(cè)定
可溶性糖、可溶性蛋白和脯氨酸含量參照鄒琦[19]的方法測(cè)定。
1.2.3 抗氧化酶活性測(cè)定
SOD活性測(cè)定采用氮藍(lán)四唑光化還原法[20]。POD活性測(cè)定采用愈創(chuàng)木酚氧化法[21]。CAT活性測(cè)定采用紫外分光光度法[22]。
1.2.4 光合色素含量測(cè)定
光合色素含量參照Arnon[23]的方法測(cè)定。
1.3 數(shù)據(jù)處理
采用Microsoft Excel 2016軟件進(jìn)行數(shù)據(jù)處理,Origin Pro 8.0軟件作圖,利用SPSS 17.0軟件進(jìn)行方差分析。表中數(shù)據(jù)以“平均值±標(biāo)準(zhǔn)差”表示。
2 結(jié)果與分析
2.1高溫脅迫對(duì)棗苗相對(duì)電導(dǎo)率和MDA含量的影響
研究表明,與對(duì)照(0 d)相比,45℃高溫處理第1 d和第3 d酸棗幼苗葉片相對(duì)電導(dǎo)率沒(méi)有顯著差異,處理第5 d相對(duì)電導(dǎo)率顯著提高18.1%。灰棗實(shí)生苗葉片相對(duì)電導(dǎo)率在處理第3 d和5 d較對(duì)照分別顯著提高25.4%和37.9%。隨著高溫處理時(shí)間的延長(zhǎng),酸棗葉片MDA含量變化不顯著,而灰棗實(shí)生苗葉片MDA含量在處理第1 d、3 d和5 d較對(duì)照分別顯著提高70.4%、121.8%和174.3%,在高溫脅迫下,酸棗葉片細(xì)胞膜穩(wěn)定性較灰棗實(shí)生苗強(qiáng),細(xì)胞膜損壞程度小,葉片受高溫傷害的程度輕。圖1
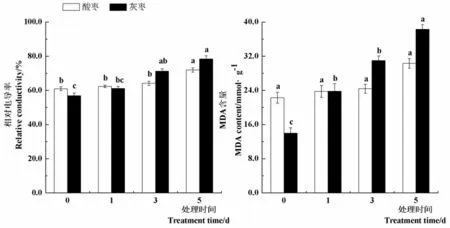
注:柱形圖上不同小寫(xiě)字母表示同一品種不同處理間差異顯著(P< 0.05),下同
Note: Different letters in the column indicate significant difference among different treatment of the same variety (P< 0.05) , the same as below
圖1 高溫脅迫下棗葉片相對(duì)電導(dǎo)率和丙二醛含量變化
Fig.1 Effect of high temperature stress on relative conductivity and MDA content of jujube leaves
2.2 高溫脅迫對(duì)棗苗滲透調(diào)節(jié)物質(zhì)含量的影響
研究表明,與對(duì)照相比,酸棗和灰棗實(shí)生苗葉片可溶性糖含量均在高溫處理第1 d顯著下降,較對(duì)照分別降低13.3%和87.6%,之后顯著上升。高溫處理下酸棗和灰棗實(shí)生苗葉片中可溶性蛋白含量變化不大,均在高溫處理第5 d達(dá)到最大值,分別為4.477和4.212 mg/g。高溫處理第3 d和5 d酸棗和灰棗實(shí)生苗葉片中脯氨酸含量較對(duì)照均顯著升高,其中酸棗分別提高82.5%和230.3%,灰棗實(shí)生苗分別提高59.5%和211.6%,由此可以說(shuō)明,酸棗和灰棗實(shí)生苗主要是通過(guò)提高葉片中可溶性糖和脯氨酸含量來(lái)抵御高溫脅迫的。圖2
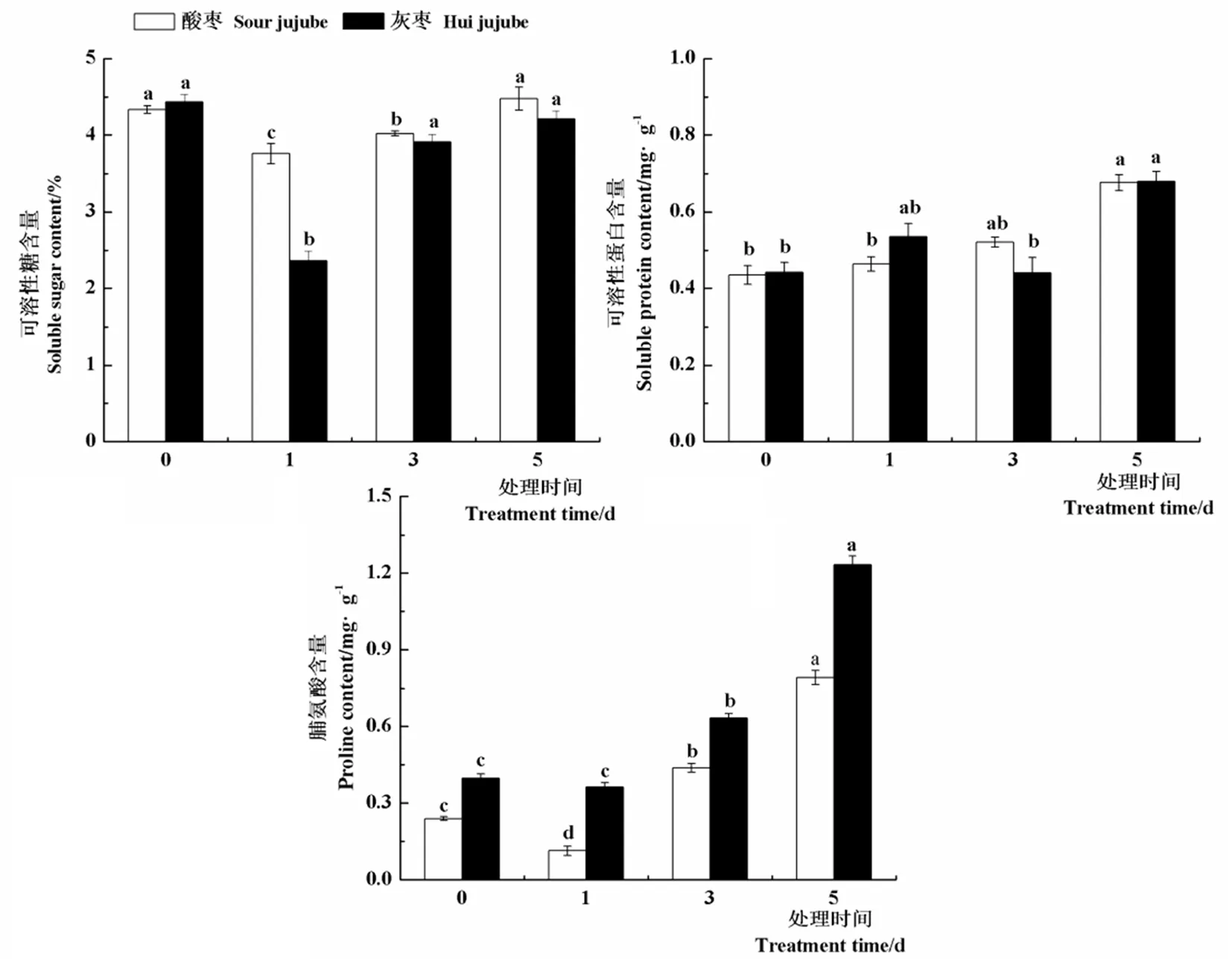
圖2 高溫脅迫下棗葉片可溶性糖、可溶性蛋白及脯氨酸含量變化
Fig.2 Effect of high temperature stress on soluble sugar, soluble protein and proline content of jujube leaves
2.3 高溫脅迫對(duì)棗苗3種抗氧化酶活性的影響
研究表明,與對(duì)照相比,隨著高溫處理時(shí)間的延長(zhǎng),酸棗葉片SOD活性顯著升高,處理第1 d、3 d和5 d葉片SOD活性較對(duì)照分別提高28.9%、61.7%和65.3%;灰棗實(shí)生苗SOD活性表現(xiàn)出先下降后上升的趨勢(shì)。高溫處理下酸棗和灰棗實(shí)生苗POD活性變化不大,均在高溫處理第5 d達(dá)到最大值,分別為592.92和575.97 U/(g·min)。與對(duì)照相比,隨著高溫處理時(shí)間的延長(zhǎng),酸棗和灰棗實(shí)生苗CAT活性均顯著升高,高溫處理第1 d、3 d和5 d酸棗葉片CAT活性分別比對(duì)照提高18.2%、67.3%和92.7%,灰棗實(shí)生苗葉片CAT活性分別比對(duì)照提高66.6%、92.5%和200.0%。由此可以看出,高溫脅迫下,酸棗和灰棗實(shí)生苗葉片通過(guò)保持較高的SOD和CAT活性來(lái)減輕高溫脅迫的傷害。圖3
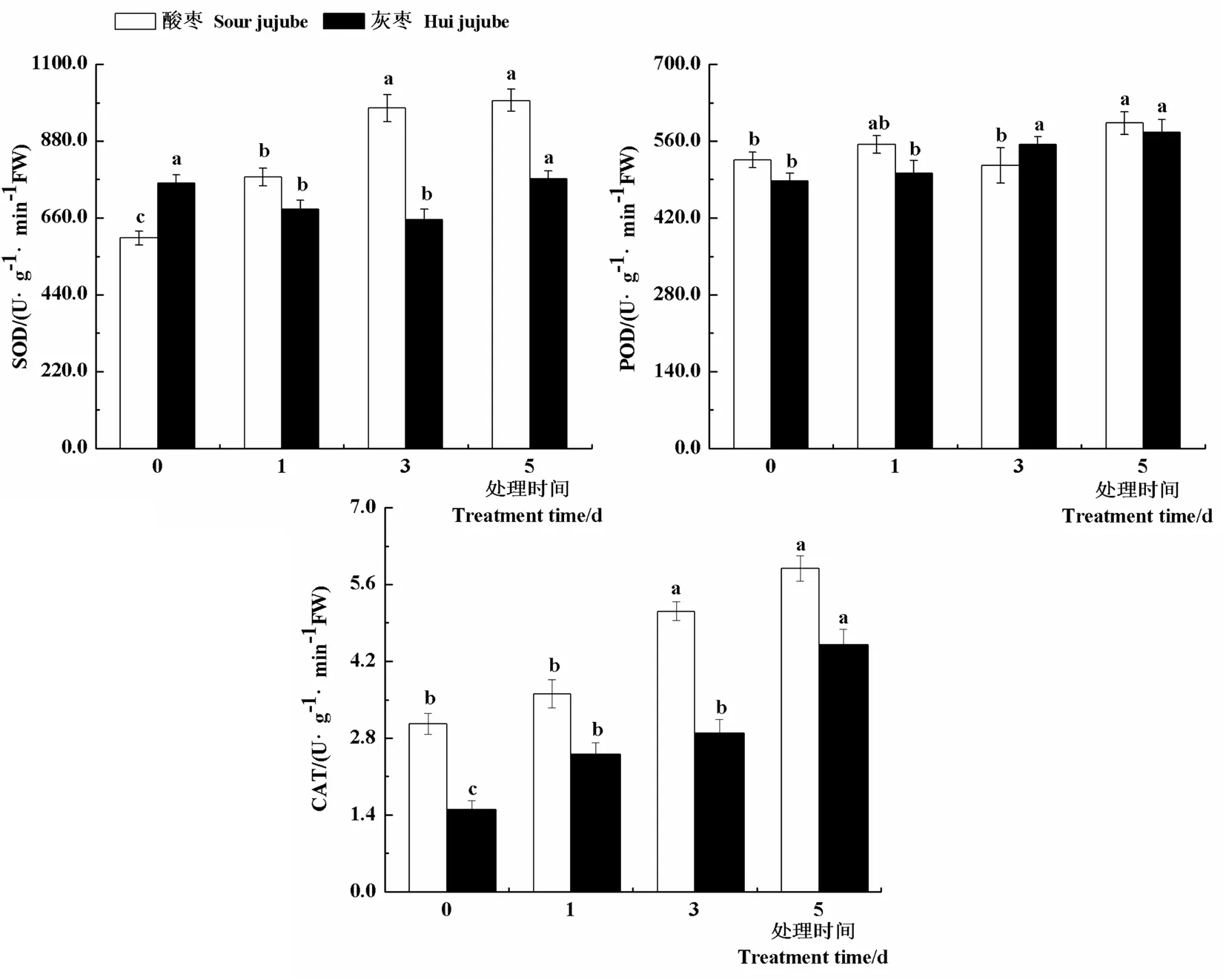
圖3 高溫脅迫下棗葉片抗氧化酶活性變化
Fig.3 Effect of high temperature stress on antioxidant enzyme activity of jujube leaves
2.4 高溫脅迫對(duì)棗苗光合色素含量的影響
在逆境條件下,植物的光合功能降低常常與光合色素的變化有關(guān)。研究表明,高溫處理1 d后,酸棗和灰棗實(shí)生苗葉片中葉綠素a、葉綠素b、葉綠素總量和類胡蘿卜素含量較對(duì)照沒(méi)有顯著差異,之后顯著下降,高溫處理5 d后酸棗和灰棗實(shí)生苗葉片中葉綠素a含量分別比對(duì)照降低13.8%和25.2%,葉綠素b含量分別比對(duì)照降低30.7%和34.6%,葉綠素總量分別比對(duì)照降低21.1%和24.1%,類胡籮卜素含量分別比對(duì)照降低18.0%和33.9%。隨著高溫脅迫時(shí)間的延長(zhǎng),酸棗和灰棗實(shí)生苗葉片光合色素含量顯著下降,光合作用受到抑制。圖4
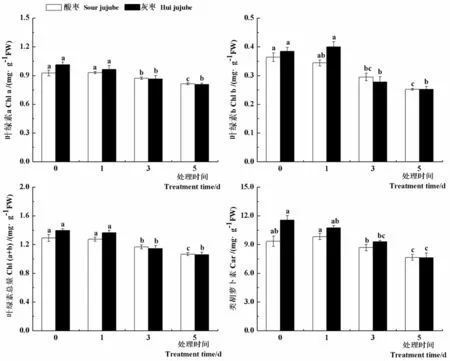
圖4 高溫脅迫下棗葉片光合色素變化
Fig.4 Effect of high temperature stress on the photosynthetic pigments of jujube leaves
3 討 論
在逆境脅迫下,植物體內(nèi)活性氧(ROS)會(huì)大量積累,活性氧代謝系統(tǒng)失去平衡,ROS攻擊細(xì)胞內(nèi)不飽和脂肪酸,膜脂過(guò)氧化加劇,導(dǎo)致膜系統(tǒng)損傷和細(xì)胞傷害,植物生理代謝受阻,嚴(yán)重時(shí)導(dǎo)致植株死亡[24-25]。高溫脅迫下植物體內(nèi)活性氧積累的直接表現(xiàn)為細(xì)胞膜透性增加[26]。高溫脅迫下梨[10]、棗[11]、葡萄[12]等果樹(shù)葉片細(xì)胞膜透性增大,植株受到傷害。研究結(jié)果顯示,高溫處理不同時(shí)間下,酸棗葉片相對(duì)電導(dǎo)率僅在處理第5 d 顯著增加,MDA含量變化不顯著,而灰棗實(shí)生苗葉片相對(duì)電導(dǎo)率和MDA含量隨著高溫處理時(shí)間的延長(zhǎng)逐漸升高,說(shuō)明在高溫脅迫下,酸棗幼苗葉片細(xì)胞膜穩(wěn)定性較灰棗實(shí)生苗強(qiáng),受高溫傷害的程度輕。
超氧化物歧化酶(SOD)、過(guò)氧化物酶(POD)、過(guò)氧化氫酶(CAT)、抗壞血酸過(guò)氧化物酶(APX)、谷胱甘肽還原酶(GR)等抗氧化酶活性與植物的耐熱性密切相關(guān)[27]。‘海沃德’獼猴桃葉片SOD、POD、CAT、APX活性在高溫脅迫初期均呈上升趨勢(shì),隨處理時(shí)間延長(zhǎng),活性下降[14]。40℃高溫脅迫下‘嘎啦’蘋(píng)果葉片APX、GR等酶活性顯著高于對(duì)照,在處理4 h達(dá)到峰值,之后下降[15]。王進(jìn)等[10]研究發(fā)現(xiàn)高溫脅迫下黃花梨和綠寶石梨葉片的SOD、CAT、POD活性先降后升再下降。試驗(yàn)中,與對(duì)照相比,隨著高溫處理時(shí)間的延長(zhǎng),酸棗苗葉片SOD和CAT活性顯著升高,灰棗實(shí)生苗SOD活性先下降后上升,CAT活性顯著升高,表明酸棗和灰棗實(shí)生苗均可通過(guò)提高體內(nèi)SOD和CAT活性來(lái)清除活性氧,減輕高溫傷害。
高溫脅迫下,植物細(xì)胞能主動(dòng)合成糖類、脯氨酸、可溶性蛋白、甜菜堿等物質(zhì),以增加細(xì)胞溶質(zhì)濃度,降低滲透勢(shì),緩和高溫脅迫[28]。Van等[29]研究發(fā)現(xiàn)44℃高溫脅迫下四季杧和臺(tái)農(nóng)一號(hào)杧2種杧果葉片可溶性糖、可溶性蛋白和脯氨酸含量均出現(xiàn)不同程度的上升,并且臺(tái)農(nóng)一號(hào)杧的游離脯氨酸含量和可溶性糖含量積累高于四季杧。42℃高溫脅迫下葡萄葉片脯氨酸含量明顯高于對(duì)照,可溶性糖含量隨脅迫時(shí)間的延長(zhǎng)呈現(xiàn)先降低后升高再降低的趨勢(shì)[30]。試驗(yàn)中,酸棗和灰棗實(shí)生苗葉片可溶性糖含量先下降后上升,可能是由于酸棗和灰棗實(shí)生苗突然受到脅迫時(shí)體內(nèi)產(chǎn)生的活性氧遠(yuǎn)遠(yuǎn)超過(guò)其正常歧化能力,造成細(xì)胞內(nèi)各種功能破壞,生理代謝紊亂,可溶性糖含量下降,之后可溶性糖含量逐漸上升,這說(shuō)明酸棗和灰棗實(shí)生苗可通過(guò)提高可溶性糖含量來(lái)保護(hù)其內(nèi)部組織,減輕高溫傷害。此外,高溫處理下酸棗和灰棗實(shí)生苗葉片中可溶性蛋白含量變化不大,酸棗和灰棗實(shí)生苗脯氨酸含量在高溫處理第3 d和5 d顯著升高,高溫脅迫可以誘導(dǎo)脯氨酸的積累,在棗實(shí)生苗抵御高溫脅迫過(guò)程中起到積極的作用。
光合作用是植物對(duì)溫度變化最為敏感的代謝反應(yīng)[31]。高溫會(huì)導(dǎo)致葉綠素分解,植物光合能力下降[32]。類胡蘿卜素既可參與植物的光合作用,還可吸收細(xì)胞內(nèi)剩余能量,淬滅活性氧,防止膜脂過(guò)氧化[33]。高溫處理降低了酸棗和灰棗實(shí)生苗葉片葉綠素a、葉綠素b、葉綠素總量和類胡蘿卜素含量,且酸棗苗葉片中光合色素含量下降幅度小于灰棗實(shí)生苗。
4 結(jié) 論
4.1 與對(duì)照相比,高溫脅迫下,酸棗葉片相對(duì)電導(dǎo)率僅在高溫脅迫第5 d顯著提高18.1%,葉片MDA含量變化不顯著,灰棗實(shí)生苗葉片相對(duì)電導(dǎo)率在高溫脅迫第3 d和5 d分別提高25.4%和37.9%,葉片MDA含量隨著脅迫時(shí)間的延長(zhǎng)較對(duì)照提高了70.4%~174.3%。
4.2 與對(duì)照相比,隨著高溫脅迫時(shí)間的延長(zhǎng),酸棗葉片脯氨酸含量提高82.5%~230.3%,可溶性糖含量先下降后上升,SOD活性提高28.9%~65.3%,CAT活性提高18.2%~92.7%;灰棗實(shí)生苗葉片脯氨酸含量提高59.5%~211.6%,可溶性蛋白、SOD活性先下降后上升,CAT活性提高66.6%~200.0%。高溫脅迫對(duì)酸棗和灰棗實(shí)生苗可溶性蛋白、POD活性影響不大。此外,高溫處理下酸棗CAT活性始終高于灰棗實(shí)生苗。
4.3 高溫脅迫5 d后,酸棗和灰棗實(shí)生苗葉片中葉綠素a、葉綠素b含量、葉綠素總量和類胡籮卜素含量顯著降低,其中酸棗分別降低13.8%、30.7%、21.1%和18.0%,灰棗實(shí)生苗分別降低25.2%、34.6%、24.1%和33.9%。
隨著高溫脅迫時(shí)間的延長(zhǎng),酸棗和灰棗實(shí)生苗葉片可溶性糖、脯氨酸、SOD和CAT活性均提高,光合色素含量均下降,并且酸棗葉片細(xì)胞膜穩(wěn)定性較灰棗實(shí)生苗強(qiáng),光合色素下降幅度較灰棗實(shí)生苗小,耐熱性較灰棗實(shí)生苗強(qiáng)。
參考文獻(xiàn)(References)
[1] 曲澤洲, 王永蕙, 彭士琪, 等. 中國(guó)果樹(shù)志·棗卷[M]. 北京:中國(guó)林業(yè)出版社, 1993:5-7.
QU Ze-zhou, WANG Yong-hui, PENG Shi-qi, et al. (1993).China'sfruittrees·Jujubevolume[M]. Beijing: China Forestry Press: 5-7.(in Chinese)
[2] Liu, M. J., Zhao, Z. H., & Liu, M. J. (2009). Germplasm resources and production of jujube in china.ActaHorticulturae, 840(840): 25-32.
[3]新疆維吾爾自治區(qū)統(tǒng)計(jì)局.新疆統(tǒng)計(jì)年鑒[J].北京:中國(guó)統(tǒng)計(jì)出版社, 2016: 372-373.
Statistics Bureau of Xinjiang Uyghur Autonomous Region. (2016).Xinjiangstatisticalyearbook[J]. Beijing: China Statistics Press: 372-373. (in Chinese)
[4] Liu, D. F., Zhang, D., Liu, G. Q., Hussain, S., & Teng, Y. W. (2013). Influence of heat stress on leaf ultrastructure, photosynthetic performance, and ascorbate peroxidase gene expression of two pear cultivars (pyrus pyrifolia).JournalofZhejiangUniversity.science.b,14(12):1,070-1,083.
[5] Luo, H. B., Ma, L., Xi, H. F., Duan, W., Li, S. H., & Loescher, W., et al. (2011). Photosynthetic responses to heat treatments at different temperatures and following recovery in grapevine (vitis amurensis l.) leaves.PlosOne, 6(8): e23033.
[6]張睿佳, 李瑛, 虞秀明, 等. 高溫脅迫與外源油菜素內(nèi)酯對(duì)'巨峰'葡萄葉片光合生理和果實(shí)品質(zhì)的影響[J].果樹(shù)學(xué)報(bào), 2015, 32(4): 590-596.
ZHANG Rui-Jia, LI Ying, YU Xiu-ming, et al. (2015). Effects of heat stress and exogenous brassinolide on photosynthesis of leaves and berry quality of'Kyoho'grapevine [J].JournalofFruitScience, 32(4): 590-596. (in Chinese)
[7]王振興, 艾軍, 陳麗, 等. 軟棗獼猴桃葉片光系統(tǒng)Ⅱ活性對(duì)不同溫度的響應(yīng)[J].西北植物學(xué)報(bào), 2015, 35(2): 329-334.
WANG Zhen-xing, AI Jun, CHEN Li, et al. (2015). Activity of photosystems Ⅱ in leaves ofActinidiaargutaunder different temperature treatments [J].ActaBot.Boreal.-Occident.Sin. , 35(2): 329-334.(in Chinese)
[8]徐斌, 白克力·塔西鐵木爾, 車?guó)P斌, 等.不同溫度環(huán)境下灰棗光合特征日變化的研究[J].新疆農(nóng)業(yè)科學(xué), 2015, 52(12): 2 222-2 229.
XU Bin, Baikeli Taximuer, CHE Feng-bin, et al. (2015).Study on the photosynthetic characteristics diurnal variation of huizao under different temperatures[J].XinjiangAgriculturalSciences, 52(12):2,222-2,229.(in Chinese)
[9]計(jì)瑋瑋, 邱翠花, 焦云, 等.高溫強(qiáng)光脅迫對(duì)砂梨葉片光合作用、D1蛋白和Deg1蛋白酶的影響[J].果樹(shù)學(xué)報(bào), 2012, 29(5):794-799.
JI Wei-wei, QIU Cui-hua, JIAO Yun, et al. (2012). Effects of high temperature and strong light on photosynthesis,D1 protein, and the Deg1 protease in pear (Pyrus pyrifolia) leaves [J].JournalofFruitScience, 29(5):794-799.(in Chinese)
[10]王進(jìn), 歐毅, 武崢, 等.高溫脅迫對(duì)早熟梨生理效應(yīng)和早期落葉的影響[J].西南農(nóng)業(yè)學(xué)報(bào), 2011, 24(2):546-551.
WANG Jin, OU Yi, WU Zheng, et al. (2011). Effects of high temperature stress on physiological indicators, early defoliation pf early maturing pear [J].SouthwestChinaJournalofAgriculturalSciences, 24(2):546-551.(in Chinese)
[11]田彩萍, 姚延梼.高溫脅迫對(duì)棗樹(shù)幼苗生長(zhǎng)發(fā)育的影響[J].天津農(nóng)業(yè)科學(xué), 2011, 17(3):15-17.
TIAN Cai-ping, YAO Yan-tao. (2011). Effect of heat stress on physiological index of jujube [J].TianjinAgriculturalSciences, 17(3):15-17. (in Chinese)
[12]孫軍利, 趙寶龍, 郁松林.外源水楊酸 (SA) 對(duì)高溫脅迫下葡萄幼苗耐熱性誘導(dǎo)研究[J].水土保持學(xué)報(bào), 2014, 28(3): 290-294.
SUN Jun-li, ZHAO Bao-long, YU Song-lin. (2014). Study of exogenous salicylic acid (SA) on the heat tolerance in grape seedlings under high temperature stress [J].JournalofSoilandWaterconservation, 28(3):290-294.(in Chinese)
[13]耶興元, 馬鋒旺, 王順才, 等.高溫脅迫對(duì)獼猴桃幼苗葉片某些生理效應(yīng)的影響[J].西北農(nóng)林科技大學(xué)學(xué)報(bào), 2004, 32(12):33-37.
YE Xing-yuan, MA Feng-wang, WANG Shun-cai, et al. (2004). Physiological effects of kiwifruit lamina under high temperature stress[J].JournalofNorthwestSci-TechUniv.ofAgri.andFor. , 32(12):33-37.(in Chinese)
[14] YH Ma, FW Ma, JK Zhang, MJ Li, YH Wang, & D Liang. (2008). Effects of high temperature on activities and gene expression of enzymes involved in ascorbate-glutathione cycle in apple leaves.PlantScience,175(6):761-766.
[15]孫軍利, 趙寶龍, 郁松林. 外源水楊酸對(duì)高溫脅迫下葡萄幾種抗氧化酶活性和抗氧化物含量的影響[J].植物生理學(xué)報(bào), 2014, 50(7):1 014-1 018.
SUN Jun-li, ZHAO Bao-long, YU Song-lin. (2014). Effect of exogenous salicylic acid on antioxidant enzymes activities and Antioxidants contents in grape seedlings under high temperature stress [J].PlantPhysiologyJournal, 50(7):1,014-1,018.(in Chinese)
[16]王鵬祥, 楊金虎. 中國(guó)西北近45a來(lái)極端高溫事件及其對(duì)區(qū)域性增暖的響應(yīng)[J].中國(guó)沙漠, 2007, 27(4):649-655.
WANG Peng-xiang, YANG Jin-hu. (2007). Extreme high temperature events and response to regional warming in recent 45 years in northwest China [J].JournalofDesertResearch, 27(4):649-655. (in Chinese)
[17]李合生.植物生理生化實(shí)驗(yàn)原理和技術(shù)[M]. 北京:高等教育出版社, 2006: 261-263.
LI He-sheng. (2006).Plantphysiologicalandbiochemicalprinciplesandexperimentaltechniques[M]. Beijing: Higher Education Press: 261-263. (in Chinese)
[18] Heath, R. L., & Packer, L. (1968). Photoperoxidation in isolated chloroplasts. i. kinetics and stoichiometry of fatty acid peroxidation.ArchivesofBiochemistry&Biophysics, 125(1): 189-198.
[19]鄒琦.植物生理學(xué)實(shí)驗(yàn)指導(dǎo)[M].北京:中國(guó)農(nóng)業(yè)出版社, 2004: 111-130.
ZOU Qi. (2004).Experimentalguidanceofplantphysiology[M]. Beijing: China Agriculture Press: 111-130. (in Chinese)
[20]王愛(ài)國(guó), 羅廣華, 邵從本, 等. 大豆種子超氧物歧化酶的研究[J]. 植物生理學(xué)報(bào), 1983, 9(1):77-84.
WANG Ai-guo, LUO Guang-hua, SHAO Cong-ben, et al. (1983). A study of superoxide dismutase of soybean seeds [J].JournalofPlantPhysiology, 9(1):77-84.(in Chinese)
[21] Omran, R. G. (1980). Peroxide levels and the activities of catalase, peroxidase, and indoleacetic acid oxidase during and after chilling cucumber seedlings.PlantPhysiology, 65(2): 407-408.
[22]郝建軍, 康宗利, 于洋. 植物生理實(shí)驗(yàn)技術(shù)[M]. 北京:化學(xué)工業(yè)出版社, 2006.
HAO Jian-jun, KANG Zong-li,YU Yang. (2006).Plantphysiologyexperimenttechnology[M]. Beijing: Chemical Industry Press.(in Chinese)
[23] Arnon, D. I. (1949). Copper enzymes in isolated chloroplasts. polyphenoloxidase in beta vulgaris.PlantPhysiology, 24(1): 1-15.
[24] Apel, K., & Hirt, H. (2004). Reactive oxygen species: metabolism, oxidative stress, and signal transduction.AnnualReviewofPlantBiology,55(x): 373-399.
[25] Miller, G., Shulaev, V., & Mittler, R. (2008). Reactive oxygen signaling and abiotic stress.PhysiologiaPlantarum, 133(3): 481-489.
[26] Yin, H., Chen, Q., & Yi, M. (2008). Effects of short-term heat stress on oxidative damage and responses of antioxidant system in lilium longiflorum.PlantGrowthRegulation, 54(1): 45-54.
[27] Almeselmani, M., Deshmukh, P. S., Sairam, R. K., Kushwaha, S. R., & Singh, T. P. (2006). Protective role of antioxidant enzymes under high temperature stress.PlantScience, 171(3): 382-388.
[28] Chen, T. H., & Murata, N. (2002). Enhancement of tolerance of abiotic stress by metabolic engineering of betaines and other compatible solutes.CurrentOpinioninPlantBiology, 5(3):250-257.
[29]VAN TOAN CAN,羅聰,何新華,等. 高溫脅迫對(duì)杧果幼苗生理生化指標(biāo)的影響[J]. 熱帶作物學(xué)報(bào),2016,37(1):53-58.
CAN VAN TOAN, LUO Cong, HE Xin-hua, et al. (2016). Effect of high temperature stress on physiology indices of mango seedlings [J].ChineseJournalofTropicalCrops, 37(1):53-58.(in Chinese)
[30]湯照云, 呂明, 張霞, 等. 高溫脅迫對(duì)葡萄葉片三項(xiàng)生理指標(biāo)的影響[J].石河子大學(xué)學(xué)報(bào), 2006, 24(2): 198-200.
TANG Zhao-yu, Lü Ming, ZHANG Xia, et al. (2006). High temperature physical coercion to several indicators of the impact of grape leaves[J].JournalofShiheziUniversity, 24(2): 198-200.(in Chinese)
[31] Virginia, W. (2011). How plants cope with temperature stress.BMCBiology, 9(1): 1-4.
[32]孫山,李鵬民,劉慶忠,等. 高灌越橘光合作用對(duì)若干環(huán)境因子的響應(yīng)[J].園藝學(xué)報(bào),2007,34 (1):67-70.
SUN Shan,LI Peng-min,LIU Qing-zhong,et al. (2007). Response of photosynthesis to some environmental factors of high-bush blueberry cultivars [J].ActaHorticulturaeSinica, 34(1):67-70. (in Chinese)
[33] Willekens, H., Van, C. W., Van, M. M., Inze, D., Langebartels, C., & Jr, S. H. (1994). Ozone, sulfur dioxide, and ultraviolet b have similar effects on mrna accumulation of antioxidant genes in nicotiana plumbaginifolia l.PlantPhysiology, 106(3):1,007-1,014.
