不同作物-香蕉輪作對香蕉生產及土壤肥力質量的影響
2018-05-08 07:32:00賴朝圓陶成圓王一鳴郭繼陽王蓓蓓阮云澤
江蘇農業學報 2018年2期
賴朝圓, 楊 越, 陶成圓, 王一鳴, 郭繼陽, 王蓓蓓, 阮云澤, 趙 艷
(1.海南大學熱帶農林學院,海南 海口 570228; 2.南京農業大學資源與環境科學學院,江蘇 南京 210095)
香蕉(Musanana)是重要的熱帶水果, 同時也是僅次于水稻(Oryzasativa)、小麥(Triticumaestivuml)、玉米(Zeamays)的第4大糧食作物[1]。目前香蕉產業己成為中國熱帶亞熱帶區域的支柱產業之一, 對南方經濟的發展和農民收入水平的提高起著舉足輕重的作用[2]。
在當前香蕉生產中, 高度集約化導致的長期連作現象普遍存在。長期連作后,土壤酸化、有害線蟲增加、土傳枯萎病病菌積累、土壤養分不平衡、土壤微生物失衡等現象愈加嚴重, 且土壤質量嚴重退化, 直接影響了香蕉產量和品質[1,3-8]。目前主要的防控措施仍存在各種問題,例如化學農藥易殘留、農業措施和生物防控效果極不穩定,抗病品種的產量和品質下降等[9-14]。而輪作能夠有效促進有機質、養分等土壤資源的循環利用, 改善土壤的理化特性, 維持土壤微生物的多樣性及活性, 抑制單一有害微生物的爆發式生長, 進而促進土地資源的可持續性利用, 是減輕或避免連作障礙發生的最佳防范措施[15-16]。前期報道, 香蕉與韭菜(Alliumtuberosum)、水稻、菠蘿(Ananascomosus)、茄子(Solanummelongena)等作物輪作, 均能有效地改善蕉園土壤的連作障礙[17-20], 然而由于香蕉種植區的氣候、地形、地貌等差異較大, 因此, 有必要開展更多輪作體系的研究。
辣椒(Capsicumannuum)、南瓜(Cucurbitamoschata)、冬瓜(Benincasahispida)和甘蔗(Saccharumofficinarum)作為海南省主要作物, 具有較高的經濟價值。本研究運用主坐標分析(Principal coordinates analysis, PCoA)和多元回歸樹分析(Multivariate regression trees, MRT)方法綜合評價與上述4種作物的輪作對香蕉生長及土壤肥力的影響, 以期篩選出適合海南香蕉生產的輪作模式, 為連作障礙蕉園土壤修復和建立科學合理的輪作制度提供參考。
1 材料與方法
1.1 試驗材料與試驗設計
1.1.1 試驗材料 2015年6月, 在走訪調查海南島北部香蕉主產區蕉園香蕉枯萎病發病情況的基礎上,選取海南省澄邁縣橋頭鎮豐西村香蕉田塊作為試驗田塊。該地屬熱帶季風氣候, 年平均氣溫23.8 ℃, 年均降雨量1 786.1 mm, 年平均日照時數2 059 h, 熱雨同季, 全年無霜。試驗香蕉品種為寶島218, 由海南萬鐘實業有限公司提供。羊糞有機肥購自海南宜康生態農業開發有限公司,有機質≥45%, 總養分(N+P2O5+K2O)≥5.0%, 水分≤30%;生物有機肥購自江陰市聯業生物科技有限公司, 總養分(N+P2O5+K2O)≥6%,1 g生物有機肥有效活菌數≥2×107個;硝酸磷型復合肥(16∶6∶20)和硫酸鉀型復合肥(15∶15∶15)購自海南諾澤農農業發展有限公司。
1.1.2 試驗設計 試驗設連作與輪作2種種植模式, 共5個處理, 每個處理設3個小區重復, 采用隨機區組設計, 每個小區種植50棵香蕉苗。連作試驗選擇連作7年以上的香蕉地塊為對照(B);輪作試驗選擇香蕉連作4年以上的地塊種植1年不同作物后再植兩季香蕉, 輪作模式設為香蕉-辣椒-香蕉-香蕉(B-PE)、香蕉-甘蔗-香蕉-香蕉(B-S)、香蕉-冬瓜-香蕉-香蕉(B-W)、香蕉-南瓜-香蕉-香蕉(B-PU)。2016年7月為第1季香蕉收獲期,2017年6月為第2季香蕉收獲期。羊糞有機肥(5 997.0 kg/hm2)和生物有機肥(1 499.2 kg/hm2)以底肥的形式施入, 花芽分化期施硝酸磷型復合肥599.7 kg/hm2, 硫酸鉀型復合肥2 998.5 kg/hm2。各處理田間管理采用香蕉常規栽培管理方法。
1.2 樣品采集
1.2.1 土壤樣品采集 2017年6月香蕉收獲期采集土壤樣品。各小區內隨機選取5株至少間隔5 m以上的健康香蕉植株采集土壤樣品, 在香蕉植株滴水線以內用土鉆隨機鉆取4個土樣, 收集0~20 cm的表層土壤, 用四分法保留1 kg左右的土壤。土樣的一半在實驗室內自然晾干用于土壤養分分析, 剩余的一半土樣4 ℃下保存以備土壤微生物試驗用。
1.2.2 香蕉果實采集 2017年6月香蕉收獲期采收香蕉果實。各小區隨機選取3株成熟健康香蕉的果實稱質量, 計算單株產量。并從中隨機選取1株香蕉果實, 切下倒數第3把香蕉用于香蕉品質測定。
1.3 測定方法
1.3.1 香蕉枯萎病發病率及防控率的測定 2017年6月香蕉收獲期統計每個小區的香蕉枯萎病發病率。將出現黃葉、葉片萎蔫、假莖呈現褐色并開裂等典型枯萎病癥狀的香蕉植株和母株早已病死而重新發出吸芽苗的香蕉植株, 均統計為發病植株。計算枯萎病發病率和防控率,發病率=(發病香蕉株數/定植香蕉總株數)×100%, 防控率=[(對照發病率-處理發病率)/對照發病率]×100%[19-20]。
1.3.2 植株長勢的測定 在香蕉收獲期每個小區隨機選取3棵健康香蕉植株記錄莖圍和株高。莖圍為距離植株基部1 m處的莖圍。株高為香蕉植株基部到植株倒數第2片與倒數第3片葉片交匯處的直線距離。
1.3.3 果實品質的測定 用乙烯利400倍液催熟采集的香蕉果實作為樣品。可溶性蛋白質含量采用考馬斯亮藍G-250染色法測定,糖度采用手持折光儀WZ-103測定,酸度用滴定法測定,維生素C含量采用抗壞血酸法測定[21]。
1.3.4 土壤養分測定 土壤有機質、堿解氮、速效鉀和有效磷含量以及土壤pH的測定均參照鮑士旦的方法[22]。
1.3.5 土壤可培養微生物數量的測定 土壤可培養放線菌、細菌、真菌及尖孢鐮刀菌的數量均采用平板稀釋涂布法測定。稱取10 g新鮮土壤樣品至90 ml無菌水中, 30 ℃、180 r/min振蕩30 min, 將土壤懸液稀釋至合適的梯度。吸取100 μl稀釋液于不同培養基平板上均勻涂布后培養。放線菌采用改良的高氏一號培養基30 ℃培養4 d,細菌采用牛肉膏蛋白胨培養基30 ℃培養1 d,真菌采用馬丁氏培養基28 ℃培養2 d,尖孢鐮刀菌采用改良的Komada培養基K2[23]28 ℃培養2 d。培養后計數平板上形成的菌落數, 并轉換成每1 g干土形成的菌落數(Colony formingunit, CFU)。土壤線蟲(Caenorhabditiselegans)數量的測定:采用淺盤法[24]分離線蟲。稱取100 g土樣, 室溫下培養48 h后, 收集線蟲懸液, 在體視顯微鏡下進行計數。
1.4 數據統計分析
采用Microsoft Excel 2003軟件對數據進行處理和繪圖。采用SPSS22.0統計分析軟件, 通過單因素方差分析(ANOVA)進行數據比較,Duncan’s新復極差法檢驗處理間差異顯著性, 利用Pearson和Spearman相關性分析比較各指標相關關系。采用R語言vegan包進行主坐標分析(PCoA)、多元回歸樹分析(MRT)。
多元回歸樹分析(MRT)在植物群落研究中是一種檢驗多物種和多環境因子之間關系的有效方法, 它是按照一定的規則通過重復分解自變量來減少樣地中群落的非相似性, 以此解釋、描述和預測多物種數據和多個環境特征之間的關系。本試驗借鑒該方法, 研究不同輪作處理后土壤的差異, 自變量為不同耕作方式(連作B為Farming=1, 輪作B-PE、B-S、B-W和B-PU為Farming=2)及不同前茬作物(B為Plant=1, B-PE為Plant=2,B-S為Plant=3,B-W為Plant=4,B-PU為Plant=5), 因變量為土壤的pH值、速效鉀含量、速效磷含量、堿解氮含量、有機質含量、細菌總數、放線菌總數、真菌總數、尖孢鐮刀菌數量和線蟲總數這10個指標。
2 結果與分析
2.1 輪作對再植香蕉生產的影響
從表1可知, 輪作可顯著促進再植香蕉植株生長, 提高香蕉果實產量。與連作對照B相比, 輪作處理(B-PE、B-S、B-W、B-PU)的香蕉株高、莖圍、產量、單把質量分別提高了 13.14%~41.80%、14.82%~38.42%、9.69%~45.61%、52.60%~104.55%。其中B-PE、B-S、B-W輪作處理的株高、莖圍、產量和單把質量均顯著比對照B提高;B-PU處理的株高、莖圍、產量與對照B間沒有顯著差異, 單把質量顯著高于對照B。由此可知B-PE、B-S、B-W輪作處理的促生效果較好, 可以顯著促進香蕉植株生長, 提高香蕉果實產量。
表1不同輪作處理對香蕉植株生長的影響
Table1Effectsofdifferentrotationtreatmentsonthegrowthofbananaplants

處理 株高(cm)莖圍(cm)產量(kg)單把質量(kg)B167.40±8.29b47.03±3.78c20.63±0.67d1.54±0.32cB?PE237.37±17.23a65.10±5.56a30.04±0.75a3.15±0.06aB?S228.93±19.34a62.90±4.73ab26.02±1.99b2.85±0.19aB?W229.10±11.66a60.30±5.37ab25.41±1.47bc2.88±0.32aB?PU189.40±14.61b54.00±7.89bc22.63±2.72cd2.35±0.13b
B: 香蕉連作對照; B-PE: 辣椒-香蕉輪作; B-S: 甘蔗-香蕉輪作; B-W: 冬瓜-香蕉輪作; B-PU: 南瓜-香蕉輪作。同列數據后不同小寫字母表示在0.05 水平上有顯著差異。
輪作處理改變了香蕉果實的品質(表2)。與連作對照B相比, 輪作處理香蕉果實的可溶性蛋白質含量提高了 3.01%~21.08%,pH值增加了 0.25~0.45, 而酸度降低了 20.56%~40.56%。其中處理B-S、B-W的可溶性蛋白質含量顯著高于對照B。輪作處理香蕉果實的pH值均顯著高于連作對照B,果實的酸度和VC含量顯著低于連作對照B。各處理間香蕉果實的可溶性固形物含量沒有明顯差異。該結果表明B-S、B-W輪作處理能在可溶性蛋白質含量這個指標上提高香蕉果實品質。
表2不同輪作處理對香蕉果實品質的影響
Table2Effectsofdifferentrotationtreatmentsonthequalityofbananafruit

處理 可溶性蛋白質含量(mg/g)可溶性固形物含量(%)pH值酸度(%)VC含量(mg/g)B1.66±0.04c20.87±0.23a4.88±0.04b7.20±0.12a0.0420±0.0002aB?PE1.71±0.03bc20.70±0.26a5.13±0.15a5.72±0.13b0.0260±0.0004dB?S2.01±0.02a20.47±0.12a5.33±0.14a5.43±0.20c0.0234±0.0002eB?W1.73±0.03b20.07±0.81a5.28±0.12a4.28±0.06d0.0299±0.0003bB?PU1.66±0.03c20.73±0.64a5.17±0.13a5.40±0.03c0.0267±0.0004c
各處理見表1注。
2.2 輪作對再植香蕉土傳枯萎病發病率及可培養尖孢鐮刀菌數量的影響
輪作處理降低了再植香蕉土傳枯萎病的發病率, 提高了防控率。連作對照B的香蕉枯萎病發病率為16.35%, 而輪作處理B-PE、B-S、B-W、B-PU的發病率分別僅為4.03%、4.37%、5.46%、9.24%(圖1)。所有輪作處理的香蕉枯萎病發病率均顯著低于連作對照, 其中B-PE、B-S、B-W處理的發病率顯著低于B-PU處理。相比于對照B,處理B-PE、B-S、B-W、B-PU的防控率分別達到75.41%、73.33%、66.62%和43.51%。其中B-PE、B-S、B-W處理的防控率顯著高于B-PU處理。說明輪作處理均對香蕉枯萎病具有較好的防控效果, 其中B-PE、B-S、B-W處理比B-PU處理效果更為顯著。
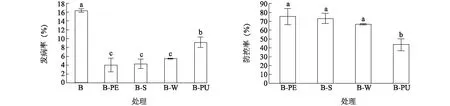
各處理見表1注。不同字母表示處理間差異達到0.05顯著水平。圖1 不同輪作處理對香蕉枯萎病發病率及防控率的影響Fig.1 Effects of different rotation treatments on the incidence and control rate of banana Fusarium wilt disease
輪作處理降低了再植香蕉收獲期土壤中可培養尖孢鐮刀菌數量(圖2)。各處理的土壤尖孢鐮刀菌數量表現出較大差異, 處理B-PE、B-S的尖孢鐮刀菌數量分別為 3.69×104CFU/g、4.18×104CFU/g, 與對照B相比, 顯著降低了 21.90%~31.11%。而對照B、處理B-W、B-PU的尖孢鐮刀菌數量高達 5.36×104CFU/g、5.74×104CFU/g、5.84×104CFU/g, 三者之間無顯著差異。該結果表明B-PE和B-S輪作處理能顯著降低土壤尖孢鐮刀菌的數量。
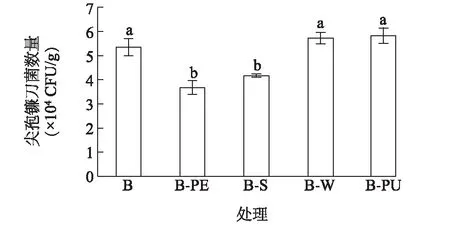
各處理見表1注。不同字母表示處理間差異達到0.05顯著水平。圖2 不同輪作處理對土壤尖孢鐮刀菌數量的影響Fig.2 Effects of different rotation treatments on the quantity of Fusarium oxysporum in soil
Pearson 相關性分析結果(圖3)顯示, 在輪作不同作物后再植香蕉收獲期的土壤中尖孢鐮刀菌數量對數值與枯萎病發病率之間呈顯著正相關關系(P=0.039,R2=0.288 7)。
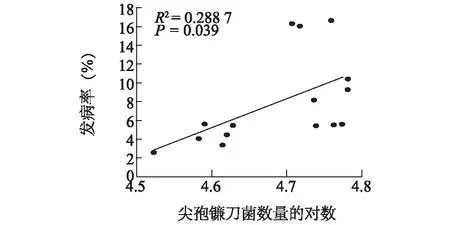
圖3 香蕉發病率與尖孢鐮刀菌數量之間的相關性Fig.3 Correlation between disease incidence and quantity of Fusarium oxysporum
將土壤養分和土壤微生物指標結合為土壤肥力質量指標, 用Pearson和Speaman相關系數計算土壤尖孢鐮刀菌數量與其他土壤肥力質量指標之間的相關性。結果(表3)顯示,在Pearson和Speaman分析結果中速效磷、細菌總數與尖孢鐮刀菌數量間均呈顯著負相關關系, 而pH值僅在Pearson分析結果中呈負相關關系。有機質含量、速效鉀含量、堿解氮含量、真菌總數、放線菌總數、線蟲總數與尖孢鐮刀菌數量之間沒有顯著相關關系。說明輪作處理制約土壤尖孢鐮刀菌數量的關鍵因子為pH值、速效磷含量、細菌總數。
表3尖孢鐮刀菌與土壤肥力質量指標的相關系數
Table3CorrelationcoefficientbetweenFusariumoxysporumandsoilfertilityqualityindex

指標 均值標準差樣品數量(N)皮爾遜相關系數斯皮爾曼相關系數pH6.240.4715-0.528?-0.395有機質含量(%)2.350.35150.1780.13速效鉀含量(mg/kg)346.5088.5415-0.418-0.484速效磷含量(mg/kg)87.8479.3715-0.799??-0.745??堿解氮含量(mg/kg)129.1222.34150.1820.183細菌總數(CFU/g)5521333.331358964.4515-0.770??-0.763??放線菌總數(CFU/g)1397733.33499019.52150.3380.327真菌總數(CFU/g)146753.33108931.09150.092-0.366線蟲總數(條/g)2.4882.0515-0.1220.073
*為置信度(雙測)為 0.05 時, 相關性顯著;**為置信度(雙測)為 0.01 時, 相關性顯著。
2.3 輪作后再植香蕉園土壤肥力質量的綜合評價
于土壤肥力質量指標的主坐標分析(PCoA)結果(圖4)顯示,輪作后再植香蕉園土壤肥力質量不同處理間差異明顯。從第1主成分(貢獻率為37.61%)來看, 輪作處理B-W、B-PU散點高、分布明顯, 能顯著地與連作對照B區分開, 表明這2個輪作處理的土壤肥力質量均與連作對照B間存在明顯差異; 但從第2主成分(貢獻率為36.11%)來看, 輪作處理B-PE、B-S、B-W散點分布明顯, 能顯著與連作對照B區分開, 而B-PU處理與連作對照B仍存在部分重合。表明不同作物輪作后能改變土壤肥力質量, B-PE、B-S、B-W處理在10個土壤肥力質量指標上與連作對照B已經完全區分開, 而B-PU處理并未區分開來。
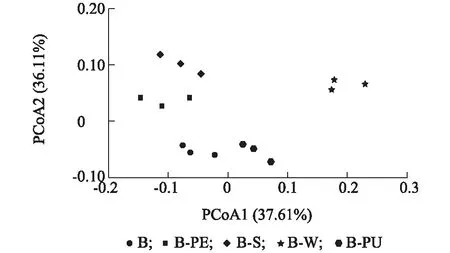
各處理見表1注。圖4 不同輪作處理后土壤肥力質量指標PCoA分析Fig.4 PCoA analysis of soil fertility quality indices after different rotation treatments
運用多元回歸樹分析(MRT)評價輪作后再植香蕉的土壤肥力質量差異。結果(圖5)顯示, 基于Bray-Curtis距離的多元回歸樹分析解釋了輪作后再植香蕉的土壤肥力質量變化的95.3%。不同處理的土壤肥力質量差異第一次分割是以前茬作物(Plant=4.5)為節點分裂出2個分支(B-PU和其他處理), 然后再根據不同耕作模式為節點(Farming=1.5)分裂成2個分支(連作模式和輪作模式), 最后再以不同前茬作物為2個節點(Plant=3.5和Plant=2.5)分裂成3個分支(B-W、B-PE、B-S)。表明與連作土壤相比, 辣椒、甘蔗及冬瓜輪作處理(B-PE、B-S、B-W)間的土壤肥力質量差異較小。同時, 不同耕作模式貢獻了總差異的39.1%, 表明不同耕作模式是造成土壤肥力質量差異的主要驅動因子。
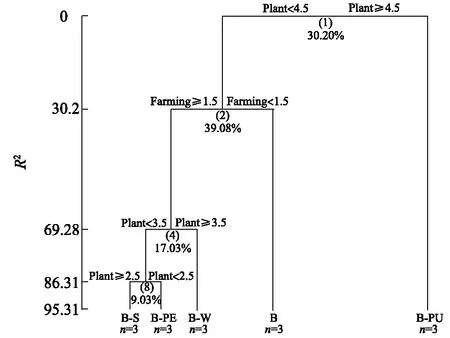
各處理見表1注。圖5 不同輪作處理后土壤肥力質量指標MRT分析Fig.5 MRT analysis of soil fertility quality indices after different rotation treatments
3 討 論
3.1 輪作處理可促進香蕉植株生長, 提高香蕉果實產量和品質
前人研究發現, 輪作不同作物(辣椒、茄子、番茄、韭菜) 后可促進香蕉生長, 從而提高香蕉產量和品質[1,18,25]。與之類似, 本研究中, 與香蕉連作對照相比, 辣椒、甘蔗、冬瓜輪作處理(B-PE、B-S、B-W) 對再植香蕉的株高、莖圍、產量和品質(可溶性蛋白質含量)均有顯著促進作用;而南瓜輪作處理(B-PU)與連作對照相比, 在株高、莖粗、產量和品質指標上也有一定程度的提高。說明辣椒、甘蔗、冬瓜輪作后可促進再植香蕉植株的生長, 也更有利于提高香蕉果實產量和品質。這可能是由于辣椒、甘蔗、冬瓜的輪作增加了土壤中微生物數量, 改變了土壤肥力水平。
3.2 輪作處理可提高土壤pH值、速效磷含量和細菌總數, 從而制約土壤尖孢鐮刀菌數量, 降低發病率
香蕉枯萎病是香蕉生產中重點防治的土傳病害, 也是香蕉連作障礙的主要構成因素, 因此抑制鐮刀菌的繁殖, 降低枯萎病的發病率是判斷克服連作障礙效果的重要指標。辛侃等[16]研究結果表明, 與連作對照相比, 水稻輪作后再植香蕉的病原菌數量下降了71.5%, 香蕉發病率降低了81.7%, 本試驗結果與其一致。與連作對照相比, 辣椒-香蕉和甘蔗-香蕉輪作處理(B-PE和B-S)均能顯著降低土壤中尖孢鐮刀菌數量(減少21.90%~ 31.11%)和香蕉枯萎病發病率(降低11.98%~12.32%), 且提高枯萎病防控率至73.33%~75.41%。而冬瓜和南瓜輪作后再植香蕉(B-W、B-PU) 并未降低土壤中尖孢鐮刀菌數量, 仍可顯著降低香蕉枯萎病的發病率, 這可能是由于本試驗結果中土壤尖孢鐮刀菌總數包含有除香蕉枯萎病專化型以外的尖孢鐮刀菌。同時, 冬瓜-香蕉輪作處理(B-W)對枯萎病的防控效果顯著高于南瓜-香蕉輪作處理(B-PU)。因此, 辣椒-香蕉和甘蔗-香蕉輪作防控香蕉土傳病害的效果更好, 冬瓜-香蕉輪作次之, 南瓜-香蕉輪作效果最差。
本研究結果顯示香蕉枯萎病發病率與尖孢鐮刀菌數量呈正相關關系(P=0.039,R2=0.288 7), 表明尖孢鐮刀菌數量與香蕉枯萎病的爆發密切相關。這與王蓓蓓[26]的研究結果一致。尖孢鐮刀菌數量與土壤肥力質量指標的相關性分析結果顯示, pH值、速效磷和總細菌數與尖孢鐮刀菌數量間均呈顯著負相關關系。有研究結果表明,土壤pH與亞麻(LinymusitatissimumL.)、草莓(Fragariaananassa)、香蕉尖孢鐮刀菌之間呈負相關關系[27-30]。Davey等[30]研究發現速效磷含量高的土壤中,由絲核菌引起的小麥根腐病發病率和茄屬絲核菌引起的莖腐病發病率均較低。洪珊等[17]的研究結果表明,輪作茄子后再植香蕉的土壤中總細菌數量顯著增加,尖孢鐮刀菌數量顯著降低。Wiggins等[31]研究發現,燕麥作為綠肥使土壤中細菌和放線菌的密度增加后,會增加土壤對疫病病原菌的抑制效果。因此,本試驗中所有輪作處理的發病率顯著比連作對照低的原因可能為: 輪作改善了土壤酸性環境, 提高了速效磷含量和土壤總細菌數量,抑制了尖孢鐮刀菌的生長, 從而促進了植物生長和增強宿主抵御土傳病原菌的能力。
3.3 輪作能緩解香蕉連作障礙, 改變土壤肥力質量
多元回歸樹分析(MRT)和主坐標分析(PCoA)是檢驗多物種和多環境因子之間關系的2個有效方法[32-33], 本研究借鑒其評價輪作后再植香蕉的土壤肥力質量差異。MRT分析結果顯示,不同輪作耕作模式的貢獻率最高, 占了總差異的39.1%, 說明不同輪作模式是造成本試驗土壤肥力質量差異的主要驅動因子。而與連作土壤相比, 辣椒、甘蔗及冬瓜輪作處理(B-PE、B-S、B-W)間的土壤肥力質量差異較小。同時, PCoA分析結果顯示, 從第1主成分(貢獻率為37.61%)上能將冬瓜、南瓜輪作處理(B-W、B-PU)與連作對照B區分開來, 而辣椒、甘蔗輪作處理(B-PE、B-S)與連作對照B存在重合部分, 這表明第一主成分可能是以前茬作物的親緣關系來聚類, 而B-W和B-PU處理中前茬作物同為葫蘆科, 所以與其他處理區分開。而從第2主成分(貢獻率為36.11%)上能將辣椒、甘蔗和冬瓜輪作后再植香蕉(B-PE、B-S、B-W)顯著與連作對照B區分開。說明這3個輪作處理在10個土壤肥力質量指標上能顯著改變連作土壤的質量, 而南瓜輪作后再植香蕉(B-PU)并未有效地改變土壤肥力質量。與此類似, 高圣超等[34]在對土壤微生物多樣性的分析中發現, 3個施肥處理的細菌群落結構在PC1 軸上聚在一起, 而與對照是分開的, 表明施肥明顯改變了土壤細菌的群落構成。結合本試驗的MRT和PCoA結果分析可知, 輪作能緩解香蕉連作障礙, 促進土壤養分和土壤微環境平衡, 從而改變土壤肥力質量。試驗中輪作辣椒、甘蔗、冬瓜后再植香蕉的輪作模式, 均有效地優化了土壤肥力質量,但南瓜輪作后再植香蕉卻未改善土壤肥力質量,其原因有待進一步研究。
參考文獻:
[1] 黃永紅. 韭菜對香蕉枯萎病的防控效果及其作用機理的研究[D]. 長沙: 湖南農業大學, 2011.
[2] 王甲水,徐碧玉,李敬陽,等. 貴州香蕉發展現狀及分析[J]. 廣東農業科學, 2013, 40(16): 44-48.
[3] 聶燕芳,黃嘉瑤,周玲菀,等. 香蕉枯萎病菌熱帶4號小種基因組規模分泌蛋白的預測與分析[J] .江蘇農業學報,2017,33(2):288-294.
[4] 孫雪婷,龍光強,張廣輝,等. 基于三七連作障礙的土壤理化性狀及酶活性研究[J]. 生態環境學報, 2015, 24(3): 409-417.
[5] 覃柳燕, 郭成林 , 黃素梅, 等. 棘孢木霉菌株PZ6對香蕉促生效應及枯萎病室內防效的影響[J].南方農業學報 ,2017,48(2):277-283.
[6] 張雪艷,張亞萍,許 帆,等. 不同種植年限黃瓜溫室土壤線蟲群落結構及多樣性的比較[J]. 植物營養與肥料學報, 2017, 23(3): 696-703.
[7] 王 飛,周登博,廉法欽,等. 香蕉枯萎病拮抗放線菌8N-10 的分離鑒定及抑菌效果評價[J]. 江蘇農業科學,2016,44(4):179-183.
[8] 劉小龍,馬建江,管吉釗,等. 連作對棉田土壤枯、黃萎病菌數量及細菌群落的影響[J]. 棉花學報, 2015, 27(1): 62-70.
[9] XU L B, HUANG B Z, WU Y L, et al. The cost-benefit analysis for bananas diversity production in China Foc. zones[J]. American Journal of Plant Sciences, 2011, 2(4): 561-568.
[10] GETHA K, VIKINESWARY S, WONG W H, et al. Evaluation of Streptomyces sp. strain g10 for suppression ofFusariumwilt and rhizosphere colonization in pot-grown banana plantlets[J]. Journal of Industrial Microbiology and Biotechnology, 2005, 32(1): 24-32.
[11] SIVAMANI E, GNANAMANICKAM S S. Biological control ofFusariumoxysporumf. sp. cubense in banana by inoculation withPseudomonasfluorescens[J]. Plant and Soil, 1988, 107(1): 3-9.
[12] 呂 順,曾莉莎,王 芳,等. 一種香蕉-甘蔗輪作克服土壤連作障礙的生態栽培方法: CN104521663A[P]. 2015-04-22.
[13] 楊瑞平. 西瓜連作障礙緩解技術及其機理研究[D]. 西安: 西北農林科技大學, 2016.
[14] 歐陽嫻,阮小蕾,吳 超,等. 香蕉輪作和連作土壤細菌主要類群[J]. 應用生態學報, 2011, 22(6): 1573-1578.
[15] 張俊芳,李 錚,李曉慧. 香蕉抗枯萎病研究現狀及展望[J]. 熱帶農業科學, 2015 , 35(12): 108-112.
[16] LARKIN R P, HALLORAN J M. Management effects of disease-suppressive rotation crops on potato yield and soilborne disease and their economic implications in potato production[J]. American Journal of Potato Research, 2014, 91(5): 429-439.
[17] 洪 珊,劇虹玲,阮云澤,等. 茄子與香蕉輪作配施生物有機肥對連作蕉園土壤微生物區系的影響[J]. 中國生態農業學報, 2017, 25(1): 78-85.
[18] 辛 侃,趙 娜,鄧小墾,等. 香蕉-水稻輪作聯合添加有機物料防控香蕉枯萎病研究[J]. 植物保護, 2014, 40(6): 36-41.
[19] WANG B B, LI R, RUAN Y Z, et al. Pineapple-banana rotation reduced the amount ofFusariumoxysporummore than maize-banana rotation mainly through modulating fungal communities[J]. Soil Biology & Biochemistry, 2015, 86: 77-86.
[20] 趙 娜. 三種茄科蔬菜輪作對高發枯萎病蕉園土壤微生物的調控效應[D]. 海口: 海南大學, 2014.
[21] 陳建勛,王曉峰. 植物生理學實驗指導[M]. 廣州: 華南理工大學出版社, 2002.
[22] 鮑士旦. 土壤農化分析[M]. 3版. 北京: 中國農業出版社, 2000.
[23] 胡 偉,趙蘭鳳,張 亮,等. 不同種植模式配施生物有機肥對香蕉枯萎病的防治效果研究[J]. 植物營養與肥料學報, 2012, 18(3): 742-748.
[24] GOODFRIEND W L, OLSEN M W, FRYE R J. Soil microfloral and microfaunal response toSalicorniabigeloviiplanting density and soil residue amendment[J]. Plant and Soil, 2000, 223: 23-32.
[25] 趙 索,周傳余,周 超,等. 蔬菜不同輪作方式對溫室甜瓜產量及根區土壤酶活性的影響[J]. 黑龍江農業科學, 2014(3): 78-80.
[26] 王蓓蓓. 輪作及生物有機肥防控香蕉土傳枯萎病的土壤微生物機制研究[D]. 南京: 南京農業大學, 2015.
[27] FANG X, YOU M P, BARBETTI M J. Reduced severity and impact ofFusariumwilt on strawberry by manipulation of soil pH, soil organic amendments and crop rotation[J]. Eur J Plant Pathol, 2012, 134(3): 619-629.
[28] 彭 雙,王一明,葉旭紅,等. 土壤環境因素對致病性尖孢鐮刀菌生長的影響[J]. 土壤, 2014, 46(5): 845-850.
[29] 沈宗專. 抑制香蕉土傳枯萎病土壤的微生物區系特征及調控[D]. 南京: 南京農業大學, 2015.
[30] 付 琳. 新墾香蕉園施用生物有機肥構建防控土傳枯萎病土壤微生物區系研究[D]. 南京:南京農業大學, 2016.
[31] WIGGINS B E, KINKEL L L. Green manures and crop sequences influence alfalfa root rot and pathogen inhibitory activity among soil-borne streptomycetes[J]. Plant and Soil, 2005, 268: 271-283.
[32] 劉 麗,徐明愷,汪思龍,等. 杉木人工林土壤質量演變過程中土壤微生物群落結構變化[J]. 生態學報, 2013, 33(15): 4692-4706.
[33] 張 榮,劉 彤. 古爾班通古特沙漠南部植物多樣性及群落分類[J]. 生態學報, 2012, 32(19): 6056-6066.
[34] 高圣超,關大偉,馬鳴超,等. 大豆連作條件下施肥對東北黑土細菌群落的影響[J]. 中國農業科學, 2017, 50(7): 1271-1281.
