缺氧環境下自噬對內皮祖細胞遷移及凋亡的影響
2018-05-10 03:37:49余森楊杰張繼航黃嵐
解放軍醫學雜志 2018年4期
余森,楊杰,張繼航,黃嵐
內皮祖細胞(endothelial progenitor cells,EPCs)是血管內皮的前體細胞,主要參與了受損內皮的修復。EPCs從骨髓、脾臟等部位動員、遷移、歸巢至血管損傷部位,在多種激素或細胞因子刺激下分化為成熟內皮細胞(endothelial cells,ECs)從而修復損傷血管,其作用在急性心肌梗死、缺血再灌注損傷、糖尿病視網膜病變、肢體缺血等血管病變中均有報道[1]。然而傳統以EPCs移植為基礎的細胞治療,在移植后普遍面臨缺血缺氧等惡劣環境[2],這導致移植后細胞存活率較低,制約了EPCs移植向臨床的轉化。因而明確EPCs在缺血缺氧等應激狀態下的存活機制,對提高移植治療效果、加強病理狀態下的血管修復有積極的指導作用。自噬(autophagy)作為真核細胞降解胞內蛋白、細胞器的重要方式,對干細胞的發育、分化、存活和歸巢都有重要作用[3-4]。本課題組前期研究發現自噬對氧化應激下EPCs的增殖存活發揮了保護作用[5],但缺氧環境下自噬對EPCs遷移、凋亡等過程有何作用還有待研究。因此,本實驗旨在觀察缺氧環境下自噬對EPCs遷移及凋亡的影響,初步探討自噬在其中發揮的作用,為缺氧環境下EPCs移植修復血管提供新的理論依據。
1 材料與方法
1.1 主要試劑及儀器 DMEM培養基和胎牛血清(FBS)購自美國Gibco公司,EGM-2MV培養基購自瑞士Lonza公司,微管相關蛋白1輕鏈3(LC3)抗體、自噬相關蛋白-7(Atg7)抗體、SQSTM1(p62)抗體購自美國Cell Signal Technology公司,β-肌動蛋白(β-actin)抗體購自美國Santa Cruz公司,淋巴細胞分離液購自美國Sigma公司,Annexin V-PE和7-AAD染液購自中國凱基生物技術公司。Transwell小室(Corning公司,美國);倒置顯微鏡(Olympus公司,日本);細胞培養箱(Kendro公司,美國);激光共聚焦顯微鏡(Leica公司,德國);流式細胞儀(Beckman Coulter公司,美國)。
1.2 EPCs的分離鑒定及培養 成年雄性SD大鼠由新橋醫院實驗動物中心提供。取大鼠脛、腓骨,PBS沖洗后收集骨髓腔沖洗液,密度梯度離心后將分離出的細胞接種于EGM-2MV的培養基中進一步培養。為證實EPCs的表型,我們采用熒光雙染法鑒定EPCs,在培養的細胞中依次加入DiI-acLDL和FITC-UEA-I,通過流式細胞術雙染陽性被確定為EPCs。
1.3 缺氧處理EPCs及分組 分離培養的EPCs每隔24h換液1次,待細胞密度為80%時,常規進行消化傳代。為檢測缺氧對EPCs遷移、凋亡及自噬水平的影響,我們將細胞分為4組:常氧培養組(即缺氧0h組),缺氧1h組,缺氧3h組和缺氧6h組;為檢測缺氧環境下自噬對EPCs的作用,我們將細胞分為4組:常氧培養組(即缺氧0h組),缺氧3h組,缺氧3h+沉默Atg7(ShAtg7)組和ShAtg7組。常氧培養條件為:37℃、5%CO2、21%O2、74%N2常氧培養箱中常規培養,缺氧培養條件為:37℃、5%CO2、1%O2、94%N2低氧培養箱中低氧培養。
1.4 流式細胞術檢測凋亡率 按前述實驗分組處理EPCs后,收集各組細胞上清液于對應離心管,PBS洗滌1次后用胰酶消化貼壁細胞1min,加入完全培養基終止消化,收集培養液與對應組的上清液細胞混合,1000×g離心3min,PBS洗滌2次后制備細胞懸液,加Annexin V-PE(AV-PE)和7-AAD染色,流式細胞儀檢測各組EPCs凋亡率。
1.5 Western blotting檢測 收集各組細胞,預冷PBS洗滌3次,加入細胞裂解液,離心后提取總蛋白,BCA蛋白定量試劑盒測定樣本蛋白質濃度。按每泳道50μg上樣量,進行10% SDS-PAGE電泳后,轉至PVDF膜;5%脫脂奶粉室溫封閉1h后,加入一抗在4℃條件下孵育過夜(一抗采用LC3、p62和β-actin,1:1000),TBST洗滌3次,加入二抗標記有辣根過氧化物酶標記的羊抗兔IgG抗體(1:5000),37℃室溫孵育1h,TBST洗滌3次,ECL顯影,利用Image J軟件進行半定量分析。
1.6 自噬相關蛋白Atg7沉默 慢病毒載體(LV) Atg7 shRNA由漢恒生物技術(上海,中國)構建。在骨髓來源的單核細胞(BMNCs)播種2d后,按照感染復數(multiplicity of infection,MOI)為100加入病毒。轉染液培養基在48h后換為新鮮培養基繼續培養。Western blotting檢測分為空白對照組(control)、空載組(vector control,VC)及ShAtg7組,以期驗證EPCs中Atg7的敲減效率。
1.7 Transwell侵襲小室實驗 取對數生長期的EPCs,調整細胞密度為3×104個/ml,在鋪好膠的上室中加入200μl細胞懸液(不含血清),下室加入500μl含VEGF的EGM-2培養液,每組設3個復孔,按前述實驗分組對EPCs進行孵育。孵育結束后從小室取出各組EPCs,4%多聚甲醛固定10min,PBS洗滌細胞3次,結晶紫染色10min,倒置顯微鏡下計數細胞數并拍照,隨機選擇視野進行統計分析。
1.8 統計學處理 采用SPSS 19.0軟件對數據進行分析。實驗數據以表示,多組間比較采用單因素方差分析(One-way ANOVA),進一步兩兩比較采用LSD-t檢驗,實驗重復≥3次。P<0.05為差異有統計學意義。
2 結 果
2.1 缺氧對EPCs遷移能力的影響 取缺氧0、1、3、6h分組處理EPCs后,利用經典的Transwell小室遷移實驗比較各組EPCs遷移能力的變化。結果顯示,隨著低氧暴露時間的延長,EPCs遷移能力逐漸降低。常氧培養組(即缺氧0h組),顯微鏡下平均每視野EPCs為50.6±5.2個,缺氧培養3h后平均每視野EPCs減少為36.60±3.36個(P<0.05),缺氧培養6h后平均每視野EPCs進一步減少為28.80±4.38個(P<0.05,圖1)。
2.2 缺氧對EPCs凋亡的影響 我們用Annexin V-PE(AV-PE)和7-AAD雙染色標記細胞,并采用流式細胞儀檢測各組EPCs凋亡水平。結果顯示,隨著低氧暴露時間的延長,EPCs凋亡率逐漸升高。常氧培養組(即缺氧0h組)凋亡率為3.87%±0.55%,缺氧3h后凋亡率(6.85%±0.91%)明顯升高(P<0.05),缺氧6h后凋亡率(9.60%±0.78%)較缺氧0h組進一步升高(P<0.05),與缺氧3h組相比凋亡率也有明顯升高(P<0.05,圖2)。
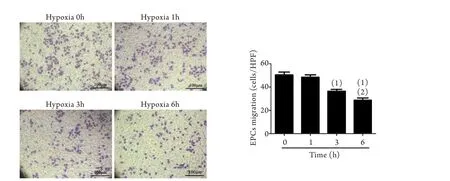
圖1 缺氧對EPCs遷移的影響(Transwell小室)Fig.1 Influence of hypoxia on EPCs migration (Transwell chambers)
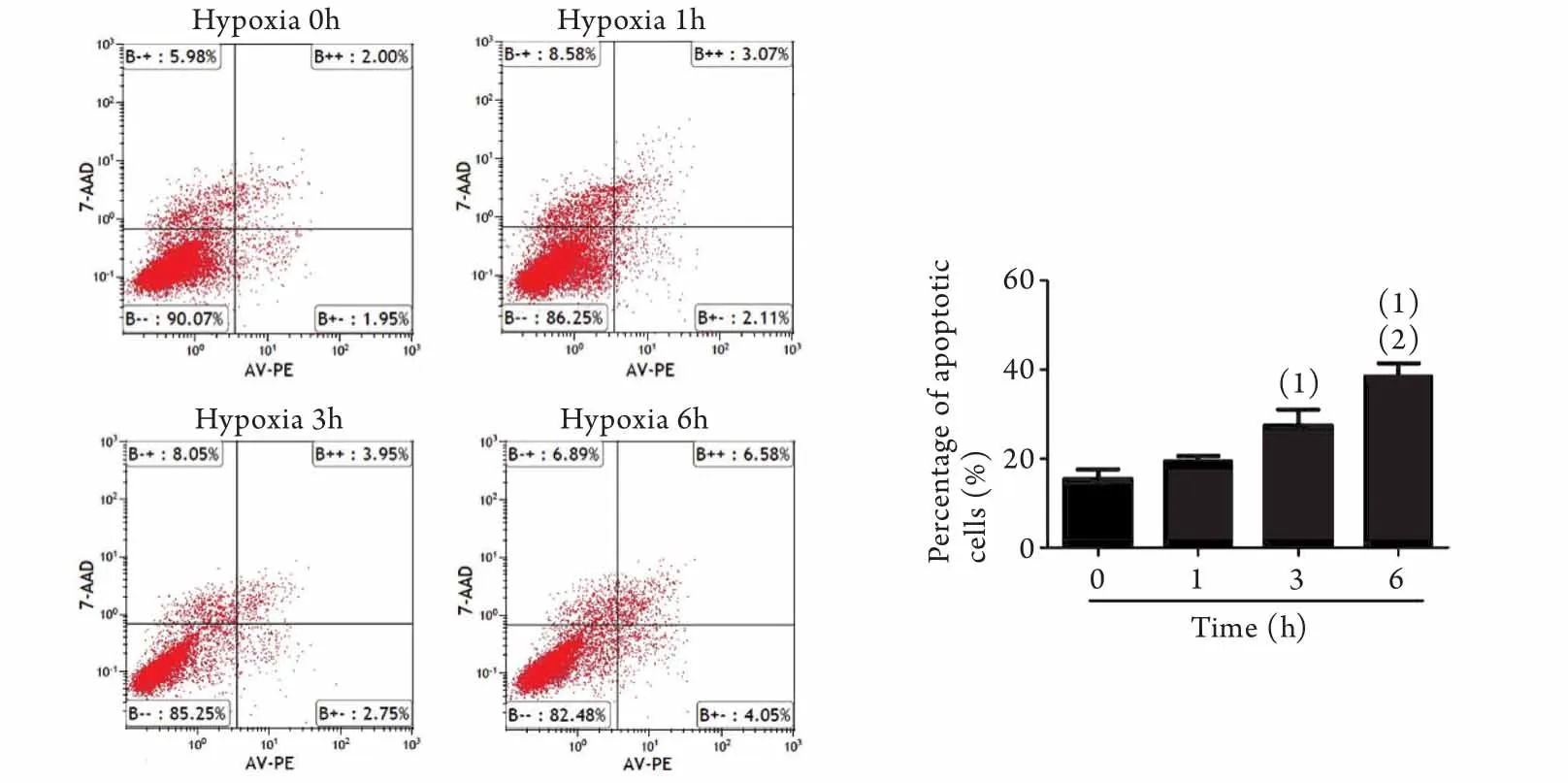
圖2 缺氧對EPCs凋亡的影響(流式細胞儀)Fig.2 Influence of hypoxia on EPCs apoptosis (Flow cytometry)
2.3 缺氧對EPCs自噬的影響 Western blotting檢測結果顯示,隨著缺氧時間的延長,EPCs自噬關鍵蛋白LC3-Ⅰ向LC3-Ⅱ的轉化增加,LC3-Ⅱ/Ⅰ比值增大,在缺氧3h后差異有統計學意義(P<0.05),自噬泛素化底物p62逐漸減少,在缺氧3h后差異有統計學意義(P<0.05,圖3)。根據遷移和凋亡結果,結合文獻[6]報道,后續研究我們采用1%氧濃度培養3h作為缺氧模擬條件。
2.4 缺氧激活的自噬在EPCs遷移和凋亡中作用我們采用慢病毒轉染方式穩定沉默自噬蛋白Atg7,Western blotting驗證Atg7被成功敲除,空載組(vector-control,VC)Atg7表達差異無統計學意義(P>0.05),LC3-Ⅱ/Ⅰ和p62表達變化證實沉默Atg7后自噬被成功抑制(圖4A、B)。我們再次利用流式細胞儀分析抑制自噬后細胞凋亡率的改變,結果顯示,缺氧3h組EPCs凋亡率(6.68%±0.95%)與缺氧0h組(3.45%±0.79%)比較明顯升高(P<0.05);缺氧3h+ShAtg7組EPCs凋亡率(8.41%±1.08%)與缺氧3h組相比,凋亡率進一步升高(P<0.05,圖4C、D)。
同時,我們利用Tanswell小室再次檢測抑制自噬后細胞遷移數目的改變,結果顯示,缺氧3h組EPCs遷移數目(34.00±3.53)明顯低于缺氧0h組(47.80±7.50,P<0.05),缺氧3h+ShAtg7組EPCs遷移數目(23.60±3.51)與缺氧3h組相比,EPCs遷移數目進一步減少(P<0.05,圖5)。
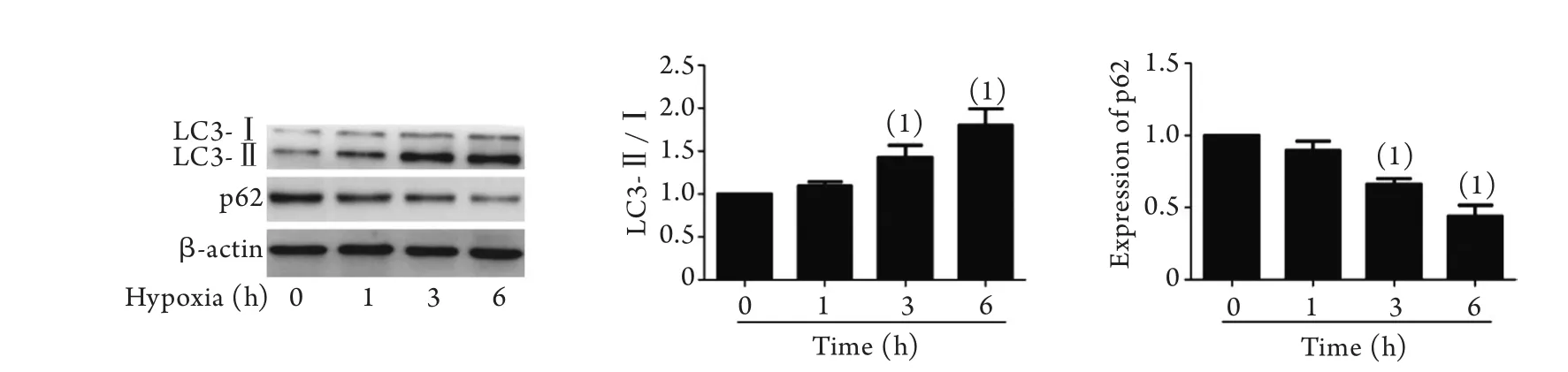
圖3 缺氧對EPCs LC3-Ⅱ/Ⅰ及p62表達的影響(Western blotting)Fig.3 Influence of hypoxia on the expressions of LC3-Ⅱ/Ⅰ and p62 (Western blotting)
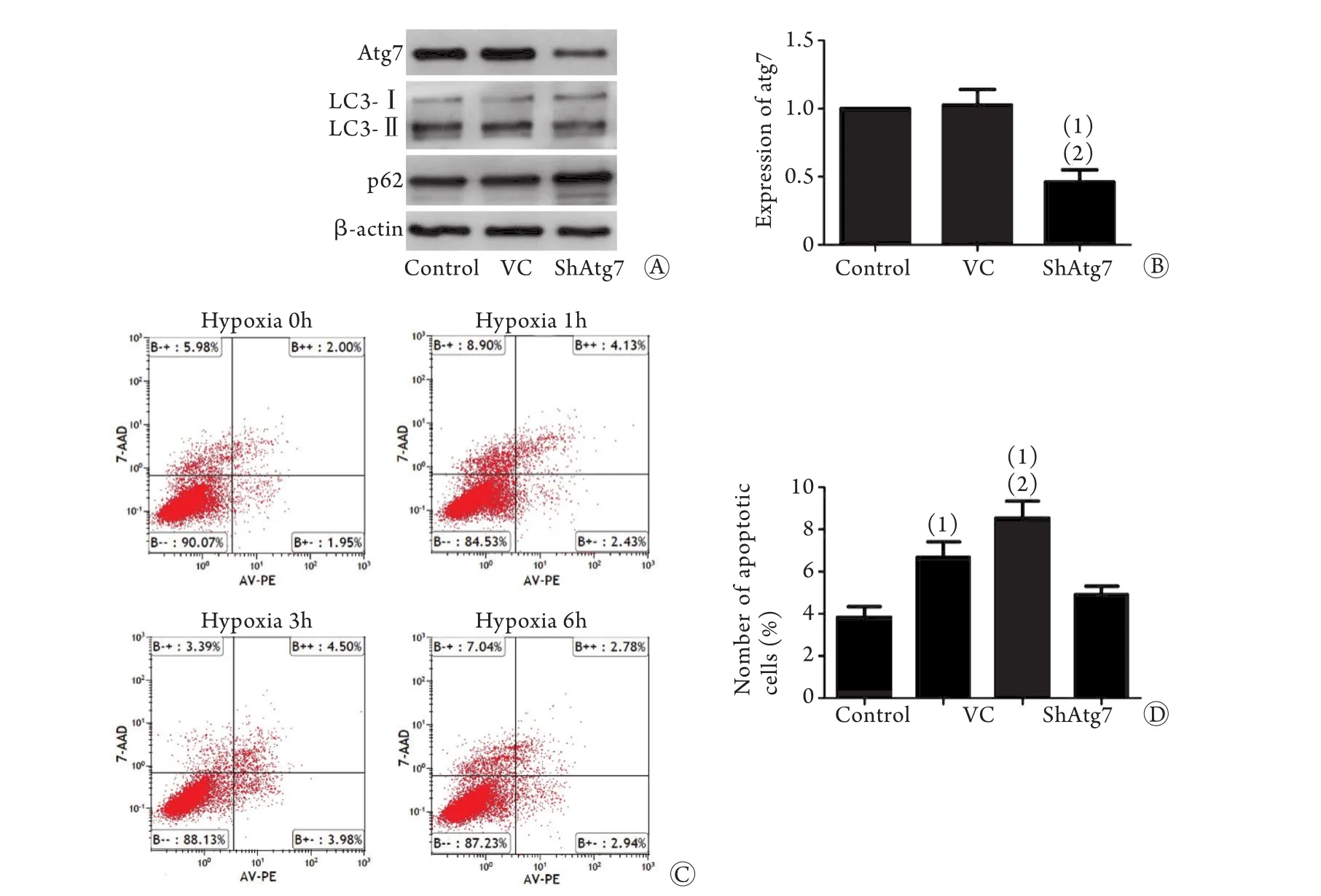
圖4 沉默Atg7表達后缺氧對EPCs凋亡的影響Fig.4 Influence of hypoxia on EPCs apoptosis after silencing Atg7 expression
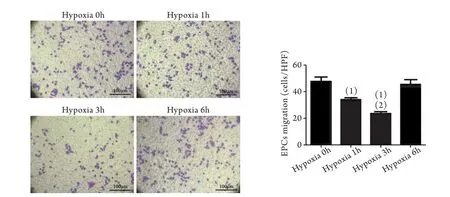
圖5 沉默Atg7表達后缺氧對EPCs遷移的影響(Transwell小室)Fig.5 Effects of hypoxia on EPCs migration after silencing Atg7 expression (Transwell chamber)
3 討 論
本研究結果表明,缺氧環境下大鼠骨髓來源的EPCs遷移能力下降,細胞凋亡率升高,同時,缺氧使EPCs內LC3-Ⅱ/Ⅰ表達增高,p62表達降低,證實缺氧是自噬的重要激活因子[7]。沉默Atg7抑制自噬后發現,與缺氧3h組比較,缺氧3h+ShAtg7組的EPCs遷移能力進一步降低,細胞凋亡率進一步升高,說明自噬在缺氧造成的損傷中發揮了保護作用。EPCs遷移和凋亡都受到自噬的影響,因此,缺氧在抑制EPCs遷移、增加細胞凋亡的同時,激活的自噬可降低缺氧對EPCs的損傷,恢復部分EPCs遷移并減少凋亡,增加EPCs的存活率。
在人體的大部分組織中,氧濃度低于3%被視為是一種低氧環境[8],本研究采用的是1%氧濃度的低氧培養箱處理細胞,由于長期缺氧可導致細胞大量死亡,使自噬作用難以體現,因此本研究采用短期缺氧處理1、3、6h后觀察EPCs遷移能力及凋亡數目變化。文獻報道處于胚胎期及骨髓內的EPCs常處在低氧狀態[9],當血管受損等發生時,EPCs動員至損傷處分化為內皮細胞,缺氧可以明顯增強缺氧誘導因子(HIF-1α)、趨化因子受體4(CXCR4)、VEGF等因子的表達,從而提高EPCs遷移、分化、增殖能力[6],因此缺氧預處理EPCs可增強細胞的抗凋亡能力。但是,缺氧預處理對EPCs生物學功能影響的最佳時間條件不甚明確,這可能與體內缺氧環境有多種神經體液調節共同作用有關。本實驗中,使用1%氧濃度缺氧處理3h后即對EPCs的遷移及凋亡產生明顯影響,說明體外環境下短期低氧處理即可激活EPCs凋亡通路并抑制遷移相關蛋白,使EPCs遷移能力減弱,凋亡增加。
自噬作為一種應激調控機制,在維持細胞正常功能中發揮著重要作用。既往研究表明適度激活的自噬可促進清除損傷的細胞器、細胞內代謝產物及突變基因,以延長細胞壽命[10]。近年來自噬與心血管疾病的聯系逐漸受到關注,尤其在缺氧缺血性血管疾病、缺血再灌注損傷[7,11]等疾病中受到廣泛研究,但自噬對于缺氧環境下EPCs的作用還有待進一步研究。目前關于缺氧激活細胞自噬的途徑主要集中在對HIF-1α的研究上[12],HIF-1α被認為是缺氧激活自噬的始動因子。Wang[13]的研究指出,由HIF-1α介導的自噬可增加細胞凋亡及氧化應激反應,并且這種損傷可被自噬抑制劑3-MA所阻斷。BNIP3/BNIP3L是缺氧誘導自噬發生的重要分子,缺氧時HIF-1α能使BNIP3/BNIP3L表達增加,競爭性地使Beclin-1從Bcl-2/Beclin-1復合體中解離并釋放,Beclin-1通過PI3K/AKT途徑調節下游多種自噬相關蛋白,從而激活缺氧誘導的自噬[11]。另外嚴重的缺氧可造成細胞內線粒體畸形和損傷,使線粒體內鈣超載,同時產生并釋放活性氧(ROS)、細胞色素C等物質,上調細胞凋亡蛋白酶(caspase),促進細胞凋亡或壞死[14],而自噬能清除受損線粒體及蛋白質,降低ROS細胞毒性,并維持細胞內能量穩定。上述為缺氧激活細胞自噬過程中可能存在的幾種途徑,在EPCs中仍有待進一步研究。
缺氧激活的自噬與細胞凋亡及遷移聯系緊密。研究發現缺氧環境下AKT和NF-κB通路對調控EPCs遷移和內皮分化發揮著重要作用[15],同時細胞自噬與凋亡存在共同通路,例如通過PI3K/AKT/mTOR途徑調節細胞凋亡與自噬,且PI3K/AKT/eNOS途徑對EPCs動員、遷移及分化都發揮著重要作用[16]。適度的低氧可激活自噬,清除細胞內的變形蛋白及衰老細胞器并提供細胞代謝所需的原材料,促進細胞在缺氧環境下的生存。然而,當缺氧暴露時間持續延長或程度加重時,自噬將被過度激活,細胞將出現自我消化,進一步導致細胞自噬性死亡或者凋亡[17-18]。因此,自噬作為一把雙刃劍,在保護細胞的同時也可能對細胞產生不良影響。在本研究中,我們并未采用傳統自噬誘導劑如mTOR抑制劑(雷帕霉素)、IP3R阻滯劑[19](Xestospongin C),因為此類自噬誘導劑常造成細胞自噬激活過度,引起細胞內細胞器及蛋白過度降解,導致細胞自噬性死亡或者凋亡的發生[20],從而掩蓋自噬對細胞的保護性作用。在后續的研究中,我們將進一步篩選及探討激活自噬的藥物,通過調控適度的自噬從而達到保護應激環境下EPCs生物學功能的作用。
綜上所述,本研究證實,缺氧可減弱EPCs的遷移能力并增加細胞凋亡,同時可激活EPCs自噬。通過適度正向調控EPCs自噬,可增強細胞遷移、抑制凋亡,從而提高EPCs生存能力。該結果將有助于為缺血再灌注損傷、缺血缺氧性心血管病等疾病治療提供一種新的策略,也為后續的深入實驗研究奠定了基礎。
【參考文獻】
[1]Wang HJ, Zhang D, Tan YZ,et al. Autophagy in endothelial progenitor cells is cytoprotective in hypoxic conditions[J]. Am J Physiol Cell Physiol, 2013, 304(7): 617-626.
[2]Chong MS, Ng WK, Chan JK. Concise review: endothelial progenitor cells in regenerative medicine: applications and challenges[J]. Stem Cells Transl Med, 2016, 5(4): 530-538.
[3]Liu J, Hao H, Huang H,et al. Hypoxia regulates the therapeutic potential of mesenchymal stem cells through enhanced autophagy[J]. Int J Low Extrem Wounds, 2015, 14(1): 63-72.
[4]Li SY, Guo M, Wang Y,et al. Effect of chloroquine on hydrogen peroxide-induced apoptosis of smooth muscle cellsviaendoplasmic reticulum stress pathway[J]. Med J Chin PLA,2015, 40(12): 960-965. [李松巖, 郭敏, 王煙, 等. 氯喹對過氧化氫誘導平滑肌細胞內質網應激途徑凋亡的影響[J]. 解放軍醫學雜志, 2015, 40(12): 960-965.]
[5]Yang J, Yu J, Li D,et al. Store-operated calcium entry-activated autophagy protects EPC proliferationviathe CAMKK2-MTOR pathway in ox-LDL exposure[J]. Autophagy, 2017, 13(1): 82-98.
[6]Zhou P, Tan Y, Wang H,et al. Hypoxic preconditioning-induced autophagy enhances survival of engrafted endothelial progenitor cells in ischaemic limb[J]. J Cell Mol Med, 2017, 21(10): 2452-2464.
[7]Dai S, Xu Q, Liu S,et al. Role of autophagy and its signaling pathways in ischemia/reperfusion injury[J]. Am J Transl Res,2017, 9(10): 4470-4480.
[8]Blasiak J, Petrovski G, Veréb Z,et al. Oxidative stress, hypoxia,and autophagy in the neovascular processes of age-related macular degeneration[J]. Biomed Res Int, 2014, 2014: 768026.
[9]Lee PSS. Endothelial progenitor cells in cardiovascular diseases[J]. World J Stem Cell, 2014, 6(3): 355-366.
[10]Li G, Lin X, Zhang H,et al. Ox-Lp(a) transiently induces HUVEC autophagyviaan ROS-dependent PAPR-1-LKB1-AMPK-mTOR pathway[J]. Atherosclerosis, 2015, 243(1): 223-235.
[11]Li M, Tan J, Miao Y,et al. The dual role of autophagy under hypoxia-involvement of interaction between autophagy and apoptosis[J]. Apoptosis, 2015, 20(6): 769-777.
[12]Fang Y, Tan J, Zhang Q. Signaling pathways and mechanisms of hypoxia-induced autophagy in the animal cells[J]. Cell Biol Int,2015, 39(8): 891-898.
[13]Wang L, Jin Z, Wang J,et al. Detrimental effect of Hypoxiainducible factor-1α-induced autophagy on multiterritory perforator flap survival in rats[J]. Sci Rep, 2017, 7(1): 11791.
[14]Patschan D, Schwarze K, Henze E,et al. Endothelial autophagy and Endothelial-to-Mesenchymal Transition (EndoMT) in eEPC treatment of ischemic AKI[J]. J Nephrol, 2016, 29(5): 637-644.
[15]Liu C, Tsai A, Li P,et al. Endothelial differentiation of bone marrow mesenchyme stem cells applicable to hypoxia and increased migration through Akt and NF-κB signals[J]. Stem Cell Res Ther, 2017, 8(1): 29.
[16]Heras-Sandoval D, Pérez-Rojas JM, Hernández-Damián J,et al. The role of PI3K/AKT/mTOR pathway in the modulation of autophagy and the clearance of protein aggregates in neurodegeneration[J]. Cell Signal, 2014, 26(12): 2694-2701.
[17]Mari?o G, Niso-Santano M, Baehrecke EH,et al. Selfconsumption: the interplay of autophagy and apoptosis[J]. Nat Rev Mol Cell Bio, 2014, 15(2): 81-94.
[18]Park S, Sun E, Lee Y,et al. Autophagy induction plays a protective role against hypoxic stress in human dental pulp cells[J]. J Cell Biochem, 2018, 119(2): 1992-2002.
[19]Singh A, Chagtoo M, Tiwari S,et al. Inhibition of inositol 1, 4,5-trisphosphate receptor induce breast cancer cell death through deregulated autophagy and cellular bioenergetics[J]. J Cell Biochem, 2017, 118(8): 2333-2346.
[20]Tian F, Dong L, Zhou Y,et al. Rapamycin-induced apoptosis in hgf-stimulated lens epithelial cells by AKT/mTOR, ERK and JAK2/STAT3 Pathways[J]. Int J Mol Sci, 2014, 15(8): 13833-13848.
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中老年保健(2021年12期)2021-08-24 03:30:40
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
科技傳播(2019年22期)2020-01-14 03:06:54
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
中國生殖健康(2019年11期)2019-01-07 01:28:02
