MBP-1在人食管癌細胞株Ec109細胞增殖、凋亡和侵襲中的作用
2018-05-10 09:13:04陳家驊李元明葉力波伊斯拉木江木沙由夫哈里木
重慶醫學 2018年10期
關鍵詞:檢測
陳家驊,李元明,王 磊,葉力波,伊斯拉木江,木沙由夫,哈里木,齊 海
(新疆醫科大學第二附屬醫院心胸外科,烏魯木齊 830063)

研究表明,食管癌是多種因素、多基因、多步驟相互作用的結果[1]。癌癥-MYC基因(cancer-MYC,C-myc)作為myc家族重要成員,具有癌基因功能,在調控腫瘤細胞增殖、轉移、侵襲及凋亡等過程中發揮重要作用[2]。有研究指出,C-myc啟動子結合蛋白1(C-mycpromoterbindingprotein1,MBP-1)可特異性結合到C-myc基因P2啟動子上而從轉錄水平上抑制C-myc基因表達,從而特異性阻斷腫瘤發生及進展過程[3]。本研究分別上調和特異性沉默人食管癌Ec109細胞中MBP-1基因,觀察其對細胞生物學特性的影響。1 材料與方法1.1 材料 人食管癌Ec109細胞株購自通派生物公司,RPMI-1640培養基、胎牛血清、二甲基亞砜(DM-SO)、胰酶購自美國Gibco公司,Trizol總RNA提取試劑盒、脂質體LipofectionTM2000轉染試劑盒購自美國Invitrogen公司,逆轉錄試劑盒和PCR試劑盒購自大連TaKaRa公司,MBP-1引物、內參引物、干擾序列、模擬序列、陰性對照序列由上海生工生物公司設計合成,四甲基偶氮唑藍(MTT)液購自北京碧云天公司,細胞周期和凋亡檢測試劑盒購自南京凱基生物公司,兔抗人MBP-1、C-myc、細胞周期素(Cyclin)D1多克隆抗體均購自美國Abcam公司,兔抗人CyclinE單克隆抗體購自美國SantaCruz公司,化學發光液購自長沙艾佳生物公司,實時定量PCR儀購自美國ABI公司,Transwell小室、流式細胞儀均購自美國BD公司。1.2 方法1.2.1 細胞培養及分組 將Ec109細胞株置于含胎牛血清的RPMI-1640培養基中,37℃、5%CO2恒溫培養箱中培養。胰酶消化后傳代。取對數生長期細胞,利用脂質體LipofectionTM2000轉染試劑盒轉染,將細胞分為4組。(1)MBP-1模擬物組:轉染MBP-1模擬序列為正義鏈5'-TGGATGTGGAATGTGT-GCGA-3',反義鏈5'-TTTGTACACCCTAAGC-CTCC-3';(2)siRNA-MBP-1組:轉染siRNA-MBP-1序列為正義鏈5'-GAAGTATGACCTGGACTTC-3',反義鏈5'-GGAGACTGAAGATACCTTC-3';(3)陰性對照組:轉染空白質粒;(4)空白對照組:不做任何處理。各組轉染后培養48h,完成后續實驗。1.2.2 實時定量PCR檢測各組細胞中MBP-1表達 取各組轉染培養48h細胞,加入細胞裂解液,提取總RNA,用紫外分光光度計檢測純度,以A260/A280≥1.80為合格。將總RNA逆轉錄為cDNA,以cDNA為模板進行PCR。引物序列如下,MBP-1:上游5'-AAGTAAGCTGTGGGCAG-3',下游5'-AGTAT-TCTCATGGGTCAC-3';GAPDH:上游5'-AAATC-CCATCACCATCTTCC-3',下游5'-TCACACCCAT-GACGAACA-3'。反應條件:92℃1min,92℃45s,56℃30s,72℃30s,連續循環40次,每個樣品均設置3個平行反應復孔。用2-△△Ct法獲得各組細胞中MBP-1相對表達量[4]。1.2.3 MTT法檢測不同轉染組細胞增殖情況 取各組對數生長期細胞,接種于96孔板,調整濃度為1×104/孔,分別于培養24、48、72、96h時,加入MTT液,培養3h,取出培養板,去除培養液,加入二甲基亞砜(DMSO),室溫下振蕩搖勻10min。酶標儀檢測450nm處各孔吸光度(A)值,每孔檢測3次,取均值[5]。1.2.4 流式細胞儀檢測各轉染組細胞凋亡情況 取各組培養48h細胞,磷酸鹽緩沖液(PBS)洗滌3次,調整細胞密度為1×106/mL,取1mL,1000r/min離心6min,取細胞,用預冷PBS液重懸,再次離心,取細胞重懸于100μL緩沖液中,加入AnnexinV和碘化丙啶(PI)各5μL,搖勻,室溫下避光放置10min,加入緩沖液400μL,1h內上機檢測[6]。1.2.5 流式細胞儀檢測各轉染組細胞周期情況 取各組培養48h細胞,PBS洗滌3次,各樣品細胞總數為2×106,于1000r/min進行離心6min,預冷PBS洗滌,加入預冷的70%乙醇1mL,搖勻,4℃過夜固定。1200r/min離心5min,取細胞沉淀,加入預冷PBS,RNaseA40μL,避光溫育25min,加入PI200μL,避光染色15min,60min內上機檢測[7]。1.2.6 Transwell檢測細胞侵襲能力 取各組培養48h細胞,胰酶消化,取細胞1×105,置于Transwell小室上室,將含10%胎牛血清RPMI-1640培養基置于下室,于37℃、5%CO2恒溫培養箱中培養12h,取出小室,多聚甲醛固定,吉姆薩(Giemsa)染色,于顯微鏡下觀察穿孔細胞數,并拍照計數,重復檢測3次。1.2.7 Westernblot法檢測 各組細胞中MBP-1、C-myc、CyclinD1和CyclinE蛋白表達取各組培養48h細胞,加入細胞裂解液,提取總蛋白,用聚氰基丙烯酸正丁酯(BCA)蛋白濃度檢測試劑盒對檢測總蛋白純度,金屬浴100℃變性10min。取25μg蛋白,進行十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(SDS-PAGE)電泳,電轉至聚偏二氟乙烯(PVDF)膜上,用5%脫脂奶粉封閉120min,TBST沖洗3次,分別將一抗MBP-1、C-myc、CyclinD1、CyclinE抗體加入(稀釋比例1∶200、1∶800、1∶500和1∶800),4℃過夜孵育,加入二抗,室溫孵育1h,TBST沖洗3次,加入ECL發光液,顯影,利用Quantity-One圖像分析軟件分析,獲得各組細胞中蛋白相對表達量[8]。1.3 統計學處理 采用SPSS21.0統計分析軟件對數據進行分析,計量資料用x±s表示,多組間比較采用單因素方差分析,兩兩比較采用LSD-t檢驗,檢驗水準α=0.05,以P<0.05為差異有統計學意義。2 結 果2.1 各組細胞中MBP-1基因表達比較 siRNA-MBP-1組、MBP-1模擬物組、陰性對照組及空白對照
組細胞中MBP-1mRNA相對表達量分別為0.44±0.05、0.85±0.10、0.66±0.08和0.63±0.07,siRNA-MBP-1組細胞中MBP-1基因表達被抑制,而MBP-1模擬物組則表達增加(P<0.05),見圖1。2.2 各組細胞增殖情況比較 與空白對照組及陰性對照組相比,MBP-1模擬物組細胞48、72、96h時A值均降低,而siRNA-MBP-1組A值均升高(P<0.05),見圖2。2.3 各組細胞凋亡率比較 siRNA-MBP-1組、MBP-1模擬物組、陰性對照組、空白對照組細胞凋亡率分別為(14.7±3.3)、(39.5±6.2)、(25.6±5.1)、(24.5±4.9)%,相比于空白對照組及陰性對照組,siRNA-MBP-1組細胞凋亡率降低,MBP-1模擬物組則升高(P<0.05),見圖3。 a:P<0.05,與siRNA-MBP-1組比較圖1 各組細胞中MBP-1基因表達比較圖2 各組細胞增殖情況 A:MBP-1模擬物組;B:siRNA-MBP-1組;C:陰性對照組;D:空白對照組圖3 各組細胞凋亡情況2.4 各組細胞周期比較 與空白對照組及陰性對照組相比,MBP-1模擬物組細胞G0/G1期比例均降低,而S期和G2/M期細胞比例均增加;siRNA-MBP-1組細胞G0/G1期比例均升高,而S期和G2/M期細胞比例均降低(P<0.05),見表1。2.5 各組細胞侵襲能力比較 siRNA-MBP-1組、MBP-1模擬物組、陰性對照組和空白對照組侵襲細胞數分別為(144.3±10.1)、(64.0±8.1)、(86.5±7.6)、(88.2±8.4)個。與空白對照組及陰性對照組相比,siRNA-MBP-1組侵襲細胞數增加,而MBP-1模擬物組則減少(P<0.05),見圖4。表1 不同轉染組細胞周期情況比較(x±s,%)組別G0/G1期S期G2/M期MBP-1模擬物組26.8±3.5ab42.3±2.6ab30.9±3.3absiRNA-MBP-1組64.2±2.1ab21.3±1.8ab14.5±1.9ab陰性對照組43.8±2.534.5±3.121.7±2.7空白對照組45.3±2.936.7±3.318.0±2.3F19.67225.38117.262P<0.01<0.01<0.01 a:P<0.05,與陰性對照組比較;b:P<0.05,與空白對照組比較 A:MBP-1模擬物組;B:siRNA-MBP-1組;C:陰性對照組;D:空白對照組圖4 各組細胞侵襲能力(Giemsa染色,×200)2.6 各組細胞中MBP-1、CyclinD1和CyclinE蛋白表達比較 與空白對照組及陰性對照組相比,MBP-1模擬物組細胞中MBP-1蛋白相對表達量升高,而siRNA-MBP-1組降低;MBP-1模擬物組細胞中C-myc、CyclinD1和CyclinE蛋白相對表達量降低,而siRNA-MBP-1組均升高,比較差異均有統計學意義(P<0.05),見表2、圖5。
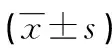
表2 各組細胞中MBP-1、CYclin D1和CYcline E蛋白表達比較
3 討 論
細胞周期紊亂、細胞侵襲能力增強是導致食管癌轉移的重要因素[9]。研究表明,癌基因C-myc過表達可加速細胞增殖,且可引起細胞DNA損傷及中心體數量異常而導致染色體破壞[10]。C-myc在多種惡性腫瘤細胞中呈過表達[11],在腫瘤惡性增殖、侵襲、轉移中發揮重要作用[12]。MBP-1作為特異性C-myc抑制基因,可在轉錄水平上抑制其表達,從而在腫瘤發生、進展過程中發揮類似于抑癌基因的功能[13]。
本研究結果顯示,MBP-1模擬物組細胞中MBP-1基因和蛋白表達量均上調,而siRNA-MBP-1組細胞中MBP-1基因和蛋白表達量均抑制,說明采用轉染模擬物和抑制物的方式可成功地將人食管癌Ec109細胞中MBP-1基因上調或抑制。MTT結果顯示,上調MBP-1基因可抑制細胞增殖,而抑制MBP-1基因則可促進細胞增殖,提示MBP-1基因與細胞增殖密切相關,是調控細胞增殖過程的重要基因。本研究結果顯示,MBP-1模擬物組細胞凋亡率顯著增加,且G0/G1期細胞比例降低,S期和G2/M期細胞比例升高,而siRNA-MBP-1組細胞凋亡率則降低,且G0/G1期細胞比例升高,S期和G2/M期細胞比例降低,說明MBP-1基因可通過影響細胞周期,而參與細胞凋亡的發生。同時,Western blot結果顯示,MBP-1模擬物組細胞中C-myc蛋白表達被抑制,而siRNA-MBP-1組則表達上調,說明MBP-1作為C-myc基因的抑制基因,可能通過抑制C-myc表達而達到減少細胞增殖、促進細胞凋亡的作用。本研究結果說明MBP-1基因與Ec109細胞侵襲能力密切相關。
研究顯示,Cyclin D1和Cyclin E作為重要的細胞周期蛋白激酶復合物,在調控細胞周期進程中發揮重要作用[14]。本研究顯示,MBP-1模擬物組細胞中C-myc、Cyclin D1和Cyclin E蛋白相對表達量均降低,而siRNA-MBP-1組均升高,說明MBP-1基因可通過促進或抑制細胞中Cyclin D1和Cyclin E蛋白表達,從而在調控細胞周期及細胞增殖、凋亡中發揮作用。
綜上所述,MBP-1與人食管癌細胞株Ec109細胞增殖、凋亡、侵襲能力密切相關,其機制可能與細胞周期異常有關。
[1]ALSOP B R,SHARMA P.Esophageal Cancer[J].Gastroenterol Clin North Am,2016,45(3):399-412.
[2]TANAKA S,NAGATA N,MINE S,et al.Endoscopic appearance of AIDS-related gastrointestinal lymphoma with C-myc rearrangements:case report and literature review[J].World J Gastroenterol,2013,19(29):4827-4831.
[3]TROJANOWICZ B,WINKLER A,HAMMJE K,et al.Retinoic acid-mediated down-regulation of ENO1/MBP-1 gene products caused decreased invasiveness of the follicular thyroid carcinoma cell lines[J].J Mol Endocrinol,2009,42(3):249-260.
[4]宋佳倫,楊琦.轉錄因子扭曲基因表達異常在早期子宮頸癌發病中的意義及對預后的影響[J].中國婦幼健康研究,2016,27(9):1066-1068.
[5]林博川.兔骨折合并腦外傷模型血清對鼠骨折愈合影響的實驗研究[D].廣州:廣州醫學院,2011.
[6]許靜.miR-21對人宮頸鱗癌的體內外分子調控機制初探[D].廣州:暨南大學,2011.
[7]范才文,向秋,田晶,等.Stathmin沉默抑制鼻咽癌5-8F細胞的惡性表型[J].天津醫藥,2013,41(5):462-464.
[8]袁金,康佳麗,廖花,等.靶向融合肽TAT-OSBP-MKK6(E)對卵巢癌裸鼠移植瘤的抑制作用[J].腫瘤,2015,35(2):161-167.
[9]XU X,XIONG X,SUN Y.The role of ribosomal proteins in the regulation of cell proliferation,tumorigenesis,and genomic integrity[J].Sci China Life Sci,2016,59(7):656-672.
[10]SUN X X,SEARS R C,DAI M S.Deubiquitinating C-myc:USP36 steps up in the nucleolus[J].Cell Cycle,2015,14(24):3786-3793.
[11]CHAE H,RYU H,CHA K,et al.Neutrophil gelatinase-associated lipocalin as a biomarker of renal impairment in patients with multiple myeloma[J].Clin Lymphoma Myeloma Leuk,2015,15(1):35-40.
[12]時汀,張建淮.IWR-1對人肝癌細胞株Hep3B的增殖及Wnt/β-catenin信號通路的抑制作用[J].腫瘤防治研究,2016,43(3):207-210.
[13]ZEMPLENI J,LIU D,CAMARA D T,et al.Novel roles of holocarboxylase synthetase in gene regulation and intermediary metabolism[J].Nutr Rev,2014,72(6):369-376.
[14]WU M,WU Y,LAN T,et al.Type Ⅱ cGMPdependent protein kinase inhibits EGF-induced JAK/STAT signaling in gastric cancer cells[J].Mol Med Rep,2016,14(2):1849-1856.
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
