鴨綠江上游江鱈的性狀變異和亞種地位
2018-05-15 02:11:52楊樹勛楊雨壯秦大公殷麗潔
生物學通報 2018年7期
關鍵詞:差異
楊樹勛 楊雨壯 鄭 偉 秦大公 殷麗潔
(1 延邊州水產站 吉林延吉 133002 2 北京大學生命科學學院 北京 100871)
0 引言
江鱈Lota lota(Linnaeus)發生學上屬北極淡水復合體的種類,是淡水魚類中最典型的冷水性魚類,酷寒期于冰下行生殖洄游并于水溫近0℃(0.1~0.2℃)時產卵。 該魚是高緯度北方水域珍貴的經濟魚類,尤以其肥大的肝臟和雄魚精巢的味美而著稱。廣泛分布于45°N 以北的歐、亞和北美的內陸水域及海灣處,最南可達美國東側40°N 的俄亥俄河上游。 在我國僅分布于新疆屬北極海水系的額爾齊斯河及東北的黑龍江水系,與黑龍江水系有著地理隔離的鴨綠江上游41~42°N 的水域亦有分布,這亦是歐亞大陸江鱈分布區的最南端角[1]。
江鱈屬鱈形目(Gadiformes)鱈亞目(Gadoidae)鱈科(Gadidae)江鱈屬(Lota),是鱈科魚類中唯一的淡水種類。依照謝維多維道夫(1948)[2]的分類,分布在歐洲、亞洲廣大北方地區水系流域,包括黑海、里海、貝加爾湖、黑龍江、烏蘇里江、鴨綠江上游的江鱈是“江鱈指名亞種”Lota lota lota(Linne);分布在東西伯利亞、 阿拉斯加、 加拿大的江鱈是“細尾江鱈亞種”Lota lota leptura(Hubbs et Schrltz);分布在北美洲的江鱈是“北美斑江鱈亞種”Lota lota maculosa(Lesueuy)[3]。
由于黑龍江水系的江鱈恰位于“江鱈指名亞種”和“細尾江鱈亞種”分布的交錯地帶,而且位置更加向南擴展,其體態構造的性狀變異問題,自然地為魚類學家所注目[4]。 1958年《生物學通報》所刊“在松花湖內采到的江鱈”一文,是我國對江鱈的首篇文字資料報道,施白南和高岫先生,對江鱈的經濟價值和生物學特性做了一般性介紹之后,曾提出對松花湖和鴨綠江江鱈的進一步研究有可能發現一個江鱈新亞種[4]。 此后時跨半個世紀未見這方面的報道。直至2002年筆者發表了對與松花湖同屬松花江水系而地理位置更靠近鴨綠江上游的牡丹江上游(含鏡泊湖)水域江鱈的性狀變異方面的研究結果。 文中用了72 尾江鱈標本37 項體態性狀的測算統計數據,與屬北極海水系的貝加爾湖支流色楞格河所產江鱈的同樣性狀進行了對比。 差異顯著性檢驗(t 檢驗)顯示,在全部37 項測算性狀中,有17 項差異極顯著(P<0.001),7 項差異顯著(P<0.05);但用差異系數公式計算,則基本均未達到亞種劃分的標準。 2 項剛剛達標的性狀經作圖分析均難予撐立,結論為盡管兩地(水域)江鱈體態性狀間存在著不同地理種群間顯著差異,其亞種地位還是同屬江鱈指名亞種Lota lota lota(Linné)[3]。
本文則是對位處歐、 亞大陸江鱈分布區最南端角,又以長白山為分水嶺與松花江上游,有著地理隔離的鴨綠江上游水域的江鱈,進行了同樣步驟的系列測量和計算對比,借以察看鴨綠江上游江鱈可否劃分出新亞種。
1 材料和方法
本文共測量1+~5+齡,體長15.6~48.8 cm 江鱈標本31 尾(♀♀19,♂♂12),均于2010年1月采自42°N 以南云峰水庫以北鴨綠江上游河段。 測量和計算的可量比例性狀和可數性狀共37 項,測量數據經統計處理后,與“江鱈指名亞種”傳統分布區貝加爾湖支流色楞格河江鱈的同樣性狀項目的統計數據(Сорокин B.H.1976)[5],用Mayr 差異系數公式C.D=|M1-M2|/(s1+s2)[6]進行計算,察看其性狀差異有否達到C.D≥1.28 的常用亞種劃分標準。對已達標的性狀項目,又與屬黑龍江水系的松花江支流牡丹江上游江鱈同樣性狀項目間計算的C.D 值進行察看對比,并將3 個水域同樣項目分別做統計數據總體柱形圖,再具體分析評價后,方得出結論性意見。
2 結果
表1~表3 匯列了鴨綠江上游31 尾江鱈標本體態性狀測量計算的統計數值及Сорокин Β.Η.(1976)報道的從貝加爾湖溯入色楞格河群體江鱈的同樣項目測量統計數據[5],以及用Mayr 差異系數公式[6]計算的2 個水域江鱈各對應性狀的差異程度的量化數值。
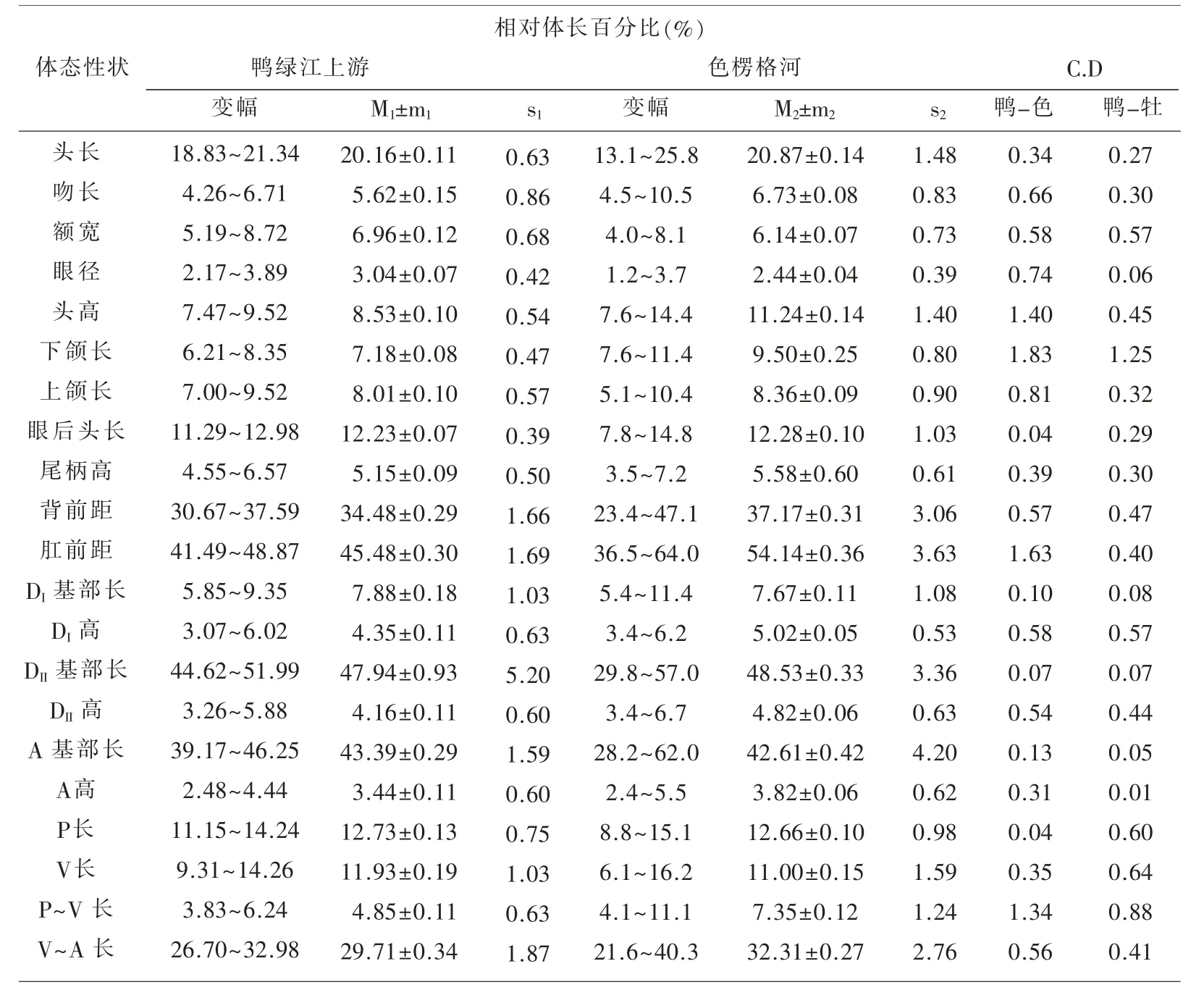
表1 鴨綠江上游與色楞格河江鱈體態性狀相對體長百分比的比較
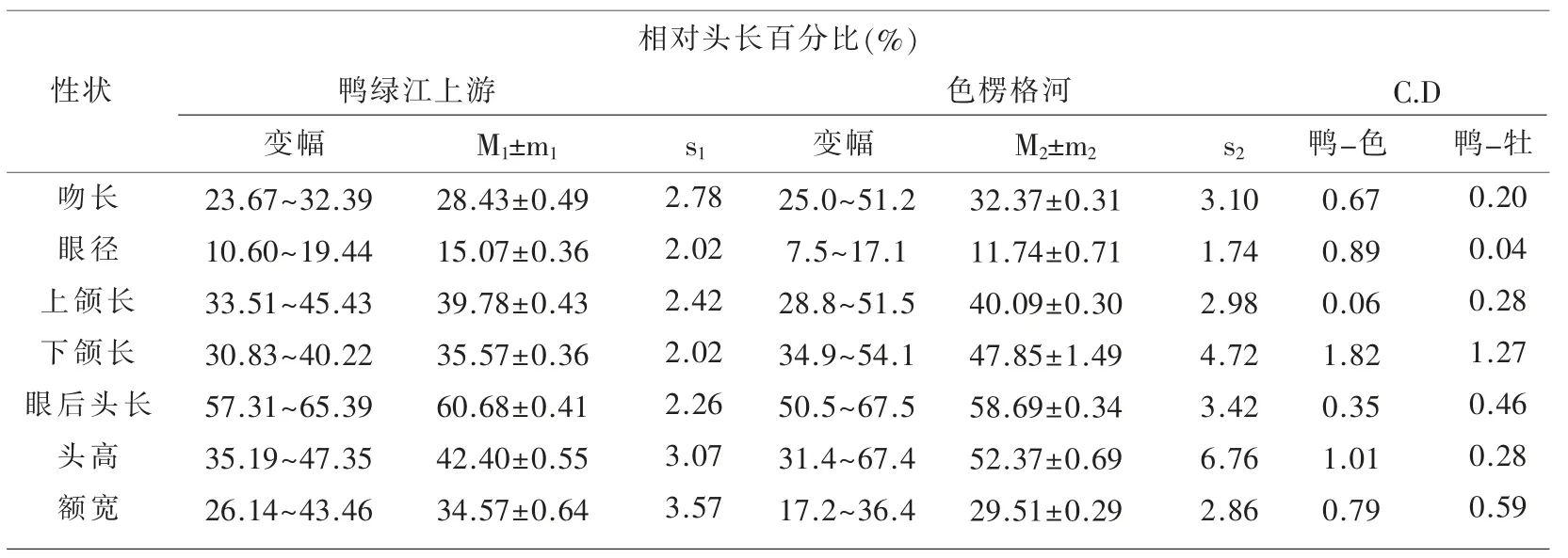
表2 鴨綠江上游與色楞格河江鱈體態性狀相對頭長百分比的比較
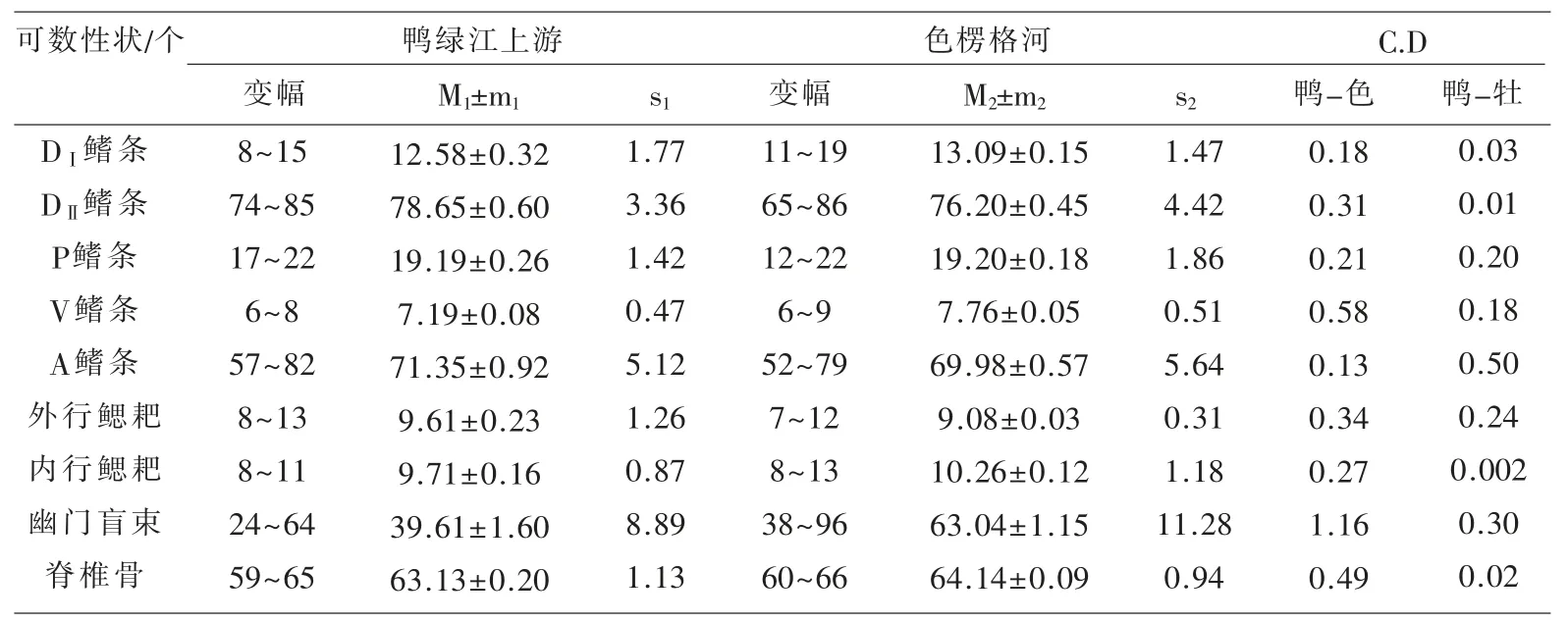
表3 鴨綠江上游與色楞格河江鱈體態可數性狀比較
用Mayr C.D≥1.28 亞種劃分標準衡量,2 個水域的江鱈間確有5 個性狀達標,即頭高相對體長的百分比(1.40)、下頜長相對體長的百分比(1.83);肛門前緣至吻端距離相對體長的百分比(1.63)、胸鰭至腹鰭距離相對體長的百分比(1.34);下頜長相對頭長的百分比(1.82)。 雖然出現了上述5 個亞種差異達標性狀,但不能就此輕易斷定出現了新亞種。 Mayr 在提出這一方法時就強調指出:“……方法并沒有可能考慮到許多在亞種確認上的生物學和生物地理學因素……還必須借助其他資料予以估量”[6]。作為江鱈指名亞種Lota lota lota(Linné)正宗產地,位處106°E50°N 屬北極海水系的貝加爾湖支流色楞格河下游水域,與位處130°E42°N 屬太平洋黃海水系的鴨綠江上游水域,其間幾乎為整個黑龍江水系的分布區,作為黑龍江最大支流的松花江的發源地(長白山天池),與鴨綠江的發源地(長白山主峰南麓)相距僅20 km,兩者屬毗鄰水域。 整個黑龍江水系包括松花江上游各支流水域均有江鱈分布,所以要確定鴨綠江上游江鱈的亞種地位,必須顧及松花江上游江鱈的性狀變異狀況和亞種地位。
經用與本文同樣的方法,筆者于2002年的研究結果已表明,對可代表松花江上游水系的牡丹江上游(含鏡泊湖)江鱈的分類地位,與流入貝加爾湖的色楞格河江鱈同為江鱈指名亞種Lota lota lota(Linné)[3]。所以上述5 個達標性狀還必須與牡丹江上游江鱈的同樣性狀進行對比檢驗。 經計算兩者間差異系數C.D 值分別為:頭高/體長(%)0.45;下 頜 長/體 長(%)1.25;肛 前 距/體 長(%)0.40;P ~V 長/體 長(%)0.88;下 頜 長/頭 長(%)1.27。5 個性狀均未達到和超過C.D≥1.28 的亞種區別標準。
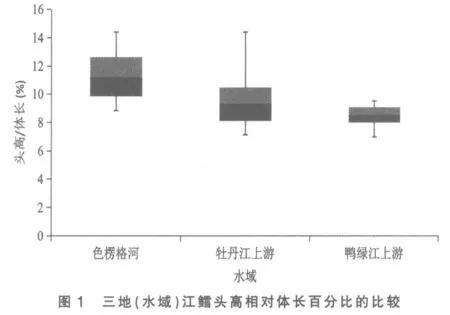
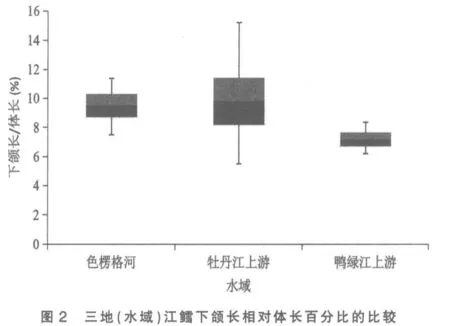
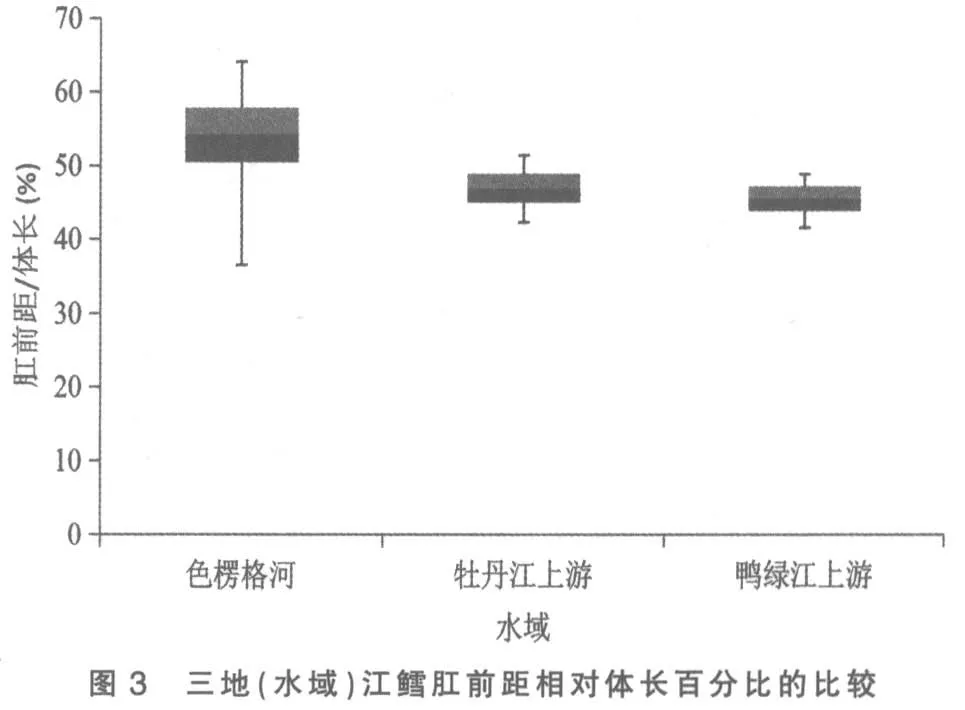
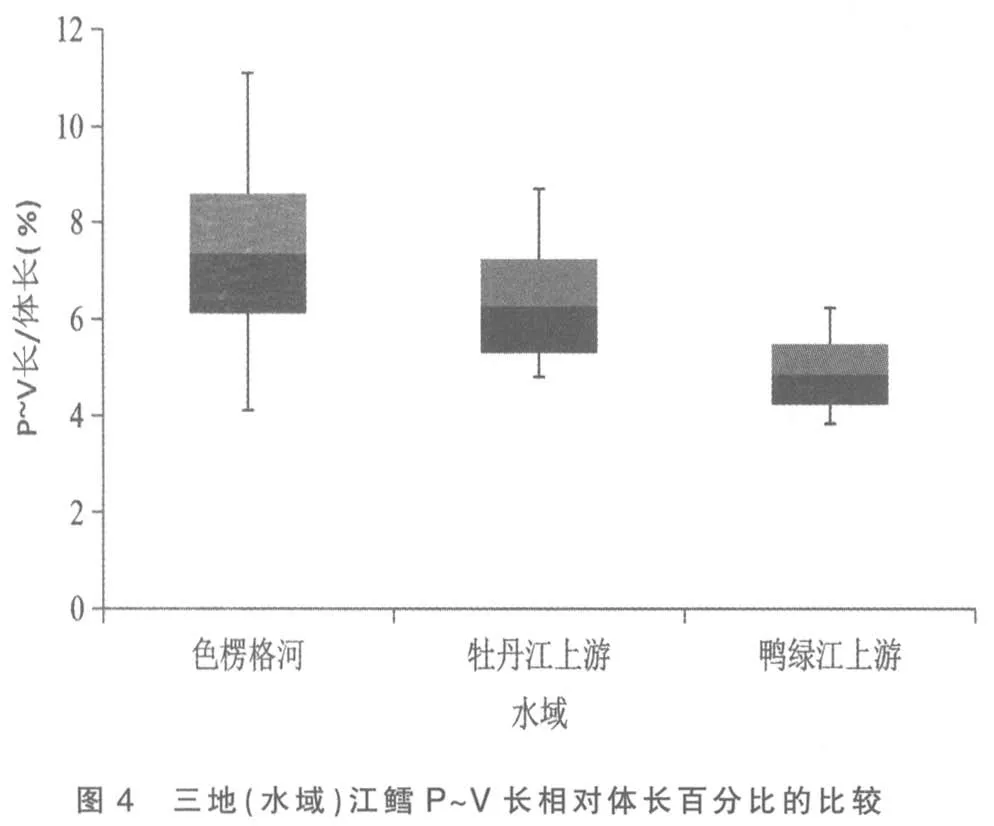
將色楞格河、 牡丹江上游和鴨綠江上游三地(水域)江鱈的這5 個性狀分別做其統計數據總體范圍及標準差對比柱形圖,以更清晰地揭示和顯現其性狀變異各類情況:圖3 為三地(水域)江鱈肛門至吻端距離與體長百分比的比較,可見鴨綠江上游江鱈所測樣本變量的中值(平均數)與色楞格河樣本中值相距較遠,但卻與牡丹江上游江鱈相近,依圖計算鴨綠江與色楞格河樣本間3 倍標準差亦不重疊(此亦Mayr 亞種差異的標志)[6],但與牡丹江上游樣本間2 倍標準差即重疊,顯然不存在亞種差異; 另外變幅較小的鴨綠江上游與牡丹江上游江鱈樣本變量總體均被變幅較大的色楞格河江鱈樣本變量總體所涵蓋,所以即使C.D 值達標,亦不能立新亞種。從圖1、圖4 看,三地(水域)江鱈的這2 個性狀樣本變量的中值及2 倍標準差均呈梯度分布,盡管鴨綠江上游與色楞格河樣本間C.D 值達標,但依圖計算,3 倍標準差彼此間均會重疊;沒達到不予重疊的亞種差異。
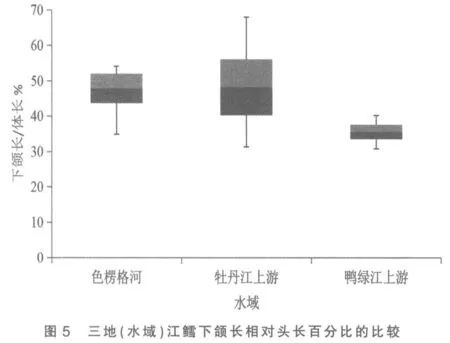
圖2 與圖5 分別是3 處江鱈下頜長與體長、下頜長與頭長百分比的總體范圍圖。 這2 個表示下頜長度的性狀是鴨綠江上游與色楞格河江鱈差異系數達標(C.D≥1.28)的5 個性狀中,C.D 值最高的性狀(1.82,1.83),與牡丹江上游江鱈也近及達標(1.25,1.27)。2 個性狀在圖上的顯示是:牡丹江上游與色楞格河樣本變量的中值非常接近,兩者2 倍標準差柱形高度重疊,表明兩者同一亞種無疑。鴨綠江上游樣本變量的中值,則與上述兩地樣本中值拉開了較大距離,2 倍標準差柱形亦不重疊,做為亞種差異特征的3 倍標準差,與色楞格河樣本確亦不重疊,但與牡丹江上游樣本卻明顯重疊(依圖觀測),表明沒有達到亞種間的差異。
綜上所述,屬太平洋黃海水系的鴨綠江上游江鱈與屬北極海水系的色楞格河江鱈間體態性狀變異的差異系數確有5 項達到亞種差異標準,但與發源地相毗鄰的松花江支流牡丹江上游江鱈間這5 個性狀均沒達到亞種分化C.D≥1.28 的差異指標。 既然屬黑龍江水系的牡丹江上游江鱈與色楞格河江鱈同為一個亞種[3],顯然鴨綠江上游江鱈亦應同屬這一亞種,即江鱈指名亞種Lota lota lota(Linné)。 三地(水域)江鱈體態性狀間的差異,顯然是不同地理種群間的差異。 做為鴨綠江上游江鱈種群體態性狀變異的突出特點是下頜較長,差異系數表明其下頜長度明顯超過牡丹江上游江鱈,更超過色楞格河江鱈,可稱之為鴨綠江長下頜種群。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
