Shh基因在NIPBL+/-胎鼠肢芽內的表達*
2018-05-29 08:27:06潘金勇嵇繼宇張惠榮石河子大學醫學院第一附屬醫院兒科新疆石河子832000
現代醫藥衛生 2018年10期
關鍵詞:小鼠
王 莉,潘金勇,嵇繼宇,張惠榮(石河子大學醫學院第一附屬醫院兒科,新疆石河子832000)
Cornelia de Lange綜合征(CdLS)是一種罕見的先天性疾病,患病率為1/100 000~1/10 000[1]。臨床表現主要為嚴重的生長發育遲緩、智力障礙、畸形等[2]。CdLS的患者約60%是NIPBL基因發生了病理性變化[3]。有研究表明,NIPBL基因水平的高低與CdLS的嚴重程度密切相關[4]。Shh基因是由ZRS增強子調控參與肢芽的發育[5],Shh基因在由ZRS增強子調控的胚胎極化活性區(ZPA)進行肢芽的表達[6]。然而,在小鼠胚胎發育階段的肢芽中,NIPBL基因對位于ZPA區域的Shh基因具有調控作用,會影響Shh基因的表達。但2個基因之間
的具體交互作用及規律鮮見文獻報道。本研究采用NIPBL-Loxp小鼠與Cre小鼠雜交,建造NIPBL+/-小鼠模型,探討NIPBL對Shh基因表達的影響,為CdLS的診斷和干預提供新的策略。
1 材料與方法
1.1 材料
1.1.1 動物 NIPBL-Loxp小鼠與Cre小鼠的背景品系小鼠是C57BL/6J,SPF級,購于浙江大學,全部小鼠飼養于石河子大學藥學院實驗動物中心。實驗過程遵照石河子大學《動物實驗倫理委員會管理條例》。挑選NIPBL+/-小鼠雄性體重35 g左右,雌性體重30 g左右,晚上20:00將雌雄小鼠按2∶1合籠,次日早晨8:00檢查雌鼠的陰栓,陰栓陽性者將次日12:00作為其胚胎E0.5天。
1.1.2 儀器與試劑 逆轉錄試劑盒Prime Script RT reagent Kit購于Thermo公司;總RNA提取劑Trizol Reagent購于Life公司;逆轉錄試劑盒RevertAid First Strand cDNA Synthesis Kit購于Thermo公司;熒光定量試劑盒SYBR Green PCR Kit購于Thermo公司;普通聚合酶鏈反應(PCR)儀器購于TaKaRa公司;熒光定量聚合酶鏈反應(qRT-PCR)儀器購于Life公司。
1.2 方法
1.2.1 動物模型的建立和實驗分組 采用NIPBL-Loxp小鼠與Cre小鼠建造NIPBL+/-小鼠模型,NIPBL+/-小鼠與NIPBL+/-小鼠按照雌、雄2∶1合籠進行雜交,分別在孕鼠的E10、E11、E12天取出孕鼠放在超凈臺內,以頸部脫臼法處死孕鼠,在嚴格無菌操作條件下,分別在孕鼠的E10、E11、E12天用逆轉錄聚合酶鏈反應(RT-PCR)逆轉錄試劑盒鑒定得到6只NIPBL+/-胎鼠的肢芽作為實驗組,另選取6只NIPBL+/+胎鼠的肢芽作為對照組,引物設計見表1;最后分離胎鼠雙側四肢,放入液氮15~20 min,再放入-80℃冰箱,以備后期實驗。

表1 NIPBL-Loxp小鼠與Cre小鼠引物信息
1.2.2 采用qRT-PCR檢測Shh基因的表達情況 Shh基因引物的設計見表2。
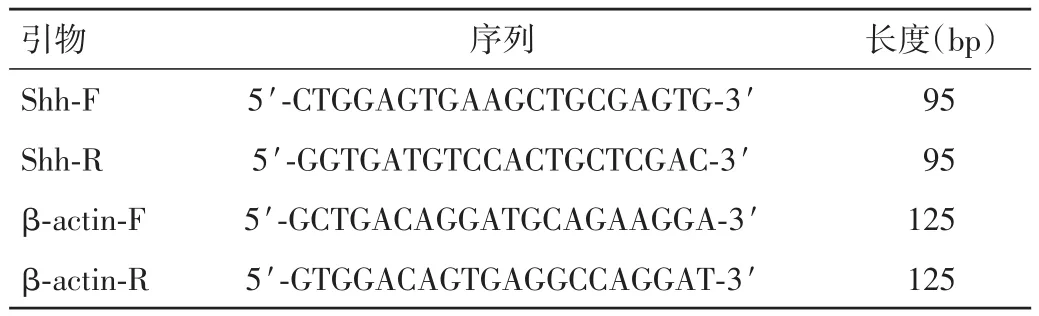
表2 Shh基因和β-actin的引物信息
1.2.3 實驗組和對照組RNA的提取 按照RNA提取試劑盒說明書提取總RNA。
1.2.4 實驗組和對照組cDNA的合成 按照逆轉錄試劑盒說明書對上述提取的總RNA進行逆轉錄。
1.2.5 qRT-PCR檢測實驗組和對照組Shh基因的表達情況 以上述cDNA為模板,按照熒光定量試劑盒說明書檢測實驗組和對照組Shh基因的表達量。
1.3 統計學處理 采用SPSS17.0統計軟件進行數據處理。符合正態分布的計量資料以±s表示,對數據進行方差齊性檢驗,方差不齊計量資料的多組間比較采用非參數檢驗,組間比較采用t檢驗。P<0.05為差異有統計學意義。
2 結 果
2.1 NIPBL基因敲除小鼠模型的驗證 NIPBL基因敲除打靶載體構建策略圖,見圖1。
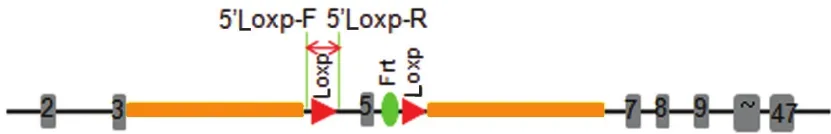
圖1 野生型及突變型等位基因示意圖
2.2 NIPBL-Loxp和Cre小鼠的引物鑒定結果 NIPBLLoxp和Cre小鼠建造的NIPBL+/-小鼠模型親代鼠,經RT-PCR技術鑒定出在孕鼠E10、E11、E12天的NIPBL+/-和NIPBL+/+胎鼠,經鑒定出來的引物凝膠電泳結果見圖 2~4。
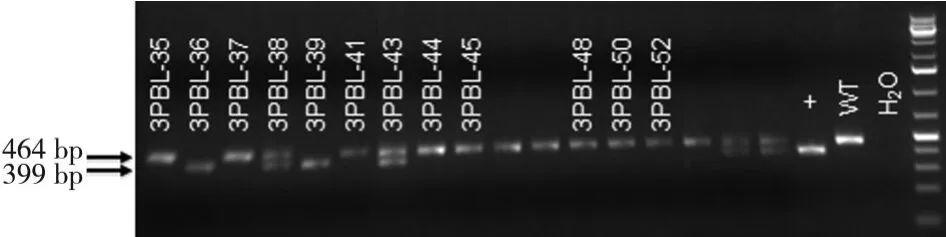
圖2 NIPBL-Loxp-F/NIPBL-Loxp-R凝膠電泳結果
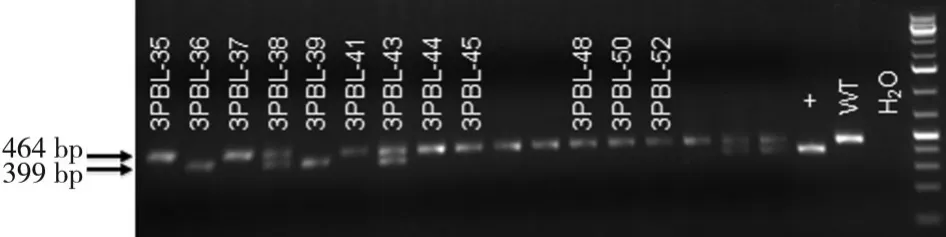
圖3 Cre-WT-F/Cre-WT-R凝膠電泳結果
圖4 Cre-WT-F/Cre-Mut-R凝膠電泳結果
2.3 qRT-PCR驗證結果 實驗組和對照組胎鼠肢芽內的Shh基因在E10、11、12天均有表達,實驗組和對照組胎鼠肢芽內Shh基因的ΔCt值由大到小依次為E10、E12、E11 天,表明在不同的孕期(E10、E11、E12天)時,實驗組和對照組胎鼠肢芽內Shh基因表達趨勢均為先升后降,即表示實驗組與對照組胎鼠肢芽的Shh基因表達量在E10天時有表達,E11天時達頂峰,E12天時表達量下降,但仍高于E10天表達量。實驗組在E10、E11、E12天Shh基因的表達量比較,差異有統計學意義(P<0.01)。與對照組比較,在相同的孕期(E10、E11、E12天)內,實驗組Shh基因的ΔCt值大于對照組,表明實驗組胎鼠肢芽內Shh基因的表達水平低于對照組,差異有統計學意義(P<0.01),見表3。
表3 實驗組與對照組Shh基因ΔCt值的比較(±s)
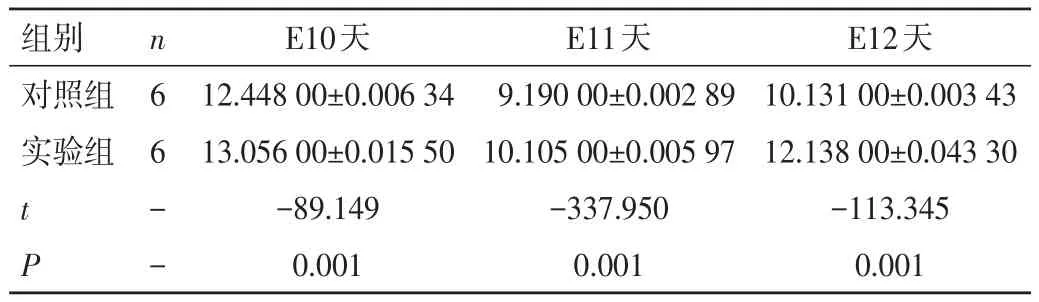
表3 實驗組與對照組Shh基因ΔCt值的比較(±s)
注:-表示無此項
組別對照組實驗組t P n6 6- -E10天12.448 00±0.006 34 13.056 00±0.015 50-89.149 0.001 E11天9.190 00±0.002 89 10.105 00±0.005 97-337.950 0.001 E12天10.131 00±0.003 43 12.138 00±0.043 30-113.345 0.001
3 討 論
CdLS是一種多系統發育障礙性的常染色體顯性遺傳疾病[7]。生長發育遲緩、面部畸形和智力障礙等是CdLS的表型特征,其中生長發育遲緩是CdLS的重要表現[8]。有關研究報道,CdLS受多種基因及信號通路的調控,一旦這些基因及信號通路發生改變,都可能導致CdLS的發生。其中,NIPBL基因缺陷是導致CdLS最常見的原因,研究人員利用分子遺傳工具創建了NIPBL基因敲除小鼠,試圖了解CdLS的病因學[9]。有關研究報道,Hedgehog信號通路抑制劑限制了骨骼的發育成熟,其包括 Sonic hedgehog(Shh)、Desert hedgehog(Dhh)及Indian hedgehog(Ihh)3 個配體,其中最常見的是 Shh[10]。Shh基因編碼一種信號蛋白,在小鼠胚胎肢芽的發育過程中起不可或缺的作用[11]。Shh基因編碼的Shh蛋白是與ZPA極化功能有關的一種形態發生素,由ZPA中細胞產生,ZPA區域傳遞肢芽早期的發育信號,啟動Shh基因的表達[12]。有研究表明,小鼠胚胎肢芽中Shh蛋白的變化存在于肢體發育前10 h,且Shh蛋白量的改變會影響肢體缺陷的程度[13]。相關研究顯示,NIPBL在ZPA區域和肢芽的間充質細胞對Shh基因進行調控,一旦NIPBL基因缺陷,會引起Shh基因表達的信號通路受到抑制,造成Shh基因的表達缺失,引起ZPA區域染色體位點異常,從而形成了CdLS的特征性改變[14]。因此,本研究通過建立NIPBL基因敲除模型,進一步研究NIPBL和Shh2個基因之間的相互作用機制。
有研究報道,Shh基因通常在正常胎鼠肢芽的E9.5~E11.5天進行表達,且在孕鼠的E10天左右表達量逐漸增多,在E11天大量表達,在E12天左右表達量逐漸減少[15]。本實驗觀察到胎鼠在E9.5天時仍是個橢球形,很難分辨出胚胎的器官,Shh基因也幾乎檢測不出表達;E12.5天時,胎鼠的肢芽也幾乎檢測不到Shh基因的表達。因此,本實驗選取胎鼠的E10、E11、E12天作為實驗分組。本實驗結果表明,在孕鼠的E10、E11、E12天,實驗組和對照組的胎鼠肢芽內均有Shh基因的表達,但是實驗組胎鼠肢芽內Shh基因的ΔCt值大于對照組,表明實驗組胎鼠肢芽內Shh基因的表達水平低于對照組(ΔCt值越高,Shh基因的表達水平越低),說明NIPBL基因缺陷影響胎鼠肢芽內Shh基因表達的正常信號通路,抑制了Shh基因的表達。實驗組和對照組胎鼠肢芽內Shh基因ΔCt值大小依次為E10、E12、E11天,表明在不同的孕期(E10、E11、E12天)內,實驗組和對照組胎鼠肢芽內Shh基因的表達趨勢是E10天時有表達,但表達量較少,隨后表達量逐漸增多,在E11天時達到頂峰,隨后表達量逐漸減少,E12天表達量較E11天減少,但比E10天表達量增多,差異有統計學意義(P<0.01)。
綜上所述,NIPBL+/-基因敲除抑制胎鼠肢芽內Shh基因的表達,而Shh基因在體內正常的軟骨發育中起重要調控作用,由此推斷NIPBL+/-基因敲除胎鼠可能會抑制軟骨的發育成熟,進一步導致CdLS。本研究有助于人們進一步了解NIPBL基因在CdLS發病中的機制和意義,并為CdLS疾病的診斷和干預提供新的策略,幫助研究人員深入了解CdLS疾病,以便探索更好的治療方法。
[1]MASKOEN AM,LAKSONO B,HAJJAH R,et al.Cornelia de lange syndrome with thyroid agenesis of an indonesian patient[J].Cell Mol Biol,2017,63(8):93-94.
[2]INFANTE E,ALKORTA-ARANBURU G,EL-GHARBAWY A,et al.Rare form of autosomal dominant familial Cornelia de Lange syndrome due to a novel duplication in SMC3[J].Clin Case Rep,2017,5(8):1277-1283.
[3]PUISAC B,TERESA-RODRIGO M,BAQUERO-MONTOYA C,et al.mRNA quantification of NIPBL isoforms a and B in adult and fet al human tissues,and a potentially pathological variant affecting only isoform a in two patients with cornelia de lange syndrome[J].Int J Mol Sci,2017,18(3):481.
[4]KAUR M,MEHTA D,NOON SE,et al.NIPBL expression levels in CdLS probands as a predictor of mutation type and phenotypic severity[J].Am J Med Genet C Semin Med Genet,2016,172(2):163-170.
[5]LETTICE LA,DEVENNEY P,DE ANGELIS C,et al.The conserved sonic hedgehog limb enhancer consists of discrete functional elements that regulate precise spatial expression[J].Cell Rep,2017,20(6):1396-1408.
[6]PELUSO S,DOUGLAS A,HILL A,et al.Fibroblast growth factors(FGFs)prime the limb specific shh enhancer for chromatin changes that balance histone acetylation mediated by E26 transformation-specific(ETS)factors[J].Elife,2017,6(1):28590.
[7]NEWKIRK DA,CHEN YY,CHIEN R,et al.The effect of Nipped-B-like(Nipbl)haploinsufficiency on genome-wide cohesin binding and target gene expression:modeling Cornelia de Lange syndrome[J].Clin Epigenetics,2017,9(1):89.
[8]PARENTI I,TERESA-RODRIGO ME,POZOJEVIC JA,et al.Mutations in chromatin regulators functionally Link Cornelia de Lange syndrome and clinically overlapping phenotypes[J].Hum Genet,2017,136(3):307-320.
[9]KAWAUCHI S,SANTOS R,MUTO AA,et al.Using mouse and zebrafish models to understand the etiology of developmental defects in cornelia de lange syndrome[J].Am J Med Genet C Semin Med Genet,2016,172(2):138-145.
[10]ROBINSON GW,KASTE SC,CHEMAITILLY W,et al.Irreversible growth plate fusions in children with medulloblastoma treated with a targeted hedgehog pathway inhibitor[J].Oncotarget,2017,8(41):69295-69302.
[11]AMANO T,SAGAI T,SEKI R,et al.Two types of etiological mutation in the Limb-Specific enhancer of shh[J].G3,2017,7(9):2991-2998.
[12]MATSUBARA H,SAITO D,ABE G,et al.Upstream regulation for initiation of restricted Shh expression in the chick limb bud[J].Dev Dynam,2017,246(5):417-430.
[13]ZHANG R,LEE C,LAWSON LY,et al.SHH protein variance in the limb bud is constrained by feedback regulation and correlates with altered digit patterning[J].G3,2017,7(3):851-858.
[14]MUTO A,IKEDA S,LOPEZ-BURKS ME,et al.Nipbl and mediator cooperatively regulate gene expression to control limb development[J].PLoS Genet,2014,10(9):1004671.
[15]YOKOYAMA S,FURUKAWA S,KITADA S,et al.Analysis of transcription factors expressed at the anterior mouse limb bud[J].PLoS One,2017,12(5):0175673.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
