高產(chǎn)幾丁質(zhì)酶菌株的 誘變選育及發(fā)酵條件的優(yōu)化
2018-05-30 19:02:36鄭家敏梁燕輝葉秀云
食品工業(yè)科技 2018年9期
關(guān)鍵詞:影響
鄭家敏,梁燕輝,朱 凡,葉秀云,林 娟
(福州大學(xué),福建省海洋酶工程重點(diǎn)實(shí)驗(yàn)室,福建福州 350116)
幾丁質(zhì)(chitin)又稱(chēng)甲殼素或殼多糖,是由N-乙酰-D-氨基葡萄糖和少量D-氨基葡萄糖通過(guò)β-1,4-糖苷鍵連接而成的線性多糖[1]。幾丁質(zhì)在自然界中儲(chǔ)量巨大,是僅次于纖維素的可再生資源,主要來(lái)源于海洋中的蝦蟹殼[2]。
幾丁質(zhì)酶(chitinase,EC3.2.1.14)是一類(lèi)能特異性催化幾丁質(zhì)水解生成幾丁寡糖或N-乙酰氨基葡萄糖的酶。1905年,Benecke首次發(fā)現(xiàn)貝內(nèi)克氏菌(Beneckeachitinovora)具有降解幾丁質(zhì)的能力[3],之后科學(xué)家相繼發(fā)現(xiàn)細(xì)菌、真菌、放線菌也能通過(guò)合成幾丁質(zhì)酶來(lái)降解幾丁質(zhì)作為自身能量來(lái)源,一些動(dòng)植物和病毒在生長(zhǎng)特殊時(shí)期也能產(chǎn)生幾丁質(zhì)酶[4-6]。幾丁質(zhì)酶降解幾丁質(zhì)生成的高附加值產(chǎn)物及其衍生物,具有抗菌益菌、降低血糖血脂、抗癌、調(diào)節(jié)人體pH、增強(qiáng)免疫力等生物活性,在食品、醫(yī)藥、農(nóng)業(yè)、化妝品等行業(yè)具有廣闊的應(yīng)用前景[7-9]。
微生物幾丁質(zhì)酶的研究已經(jīng)取得了一定的進(jìn)展,但菌株產(chǎn)酶量低且不穩(wěn)定限制了幾丁質(zhì)酶的生產(chǎn)應(yīng)用。目前,國(guó)內(nèi)外學(xué)者用來(lái)提高幾丁質(zhì)酶活力的方法大多可分為傳統(tǒng)誘變育種技術(shù)、
基因工程技術(shù)和基因重排技術(shù)。傳統(tǒng)誘變育種技術(shù)包括物理和化學(xué)誘變。研究發(fā)現(xiàn),使用紫外線、X射線、微波輻射、激光、離子束等物理方法均可取得良好的誘變效果,具有設(shè)備簡(jiǎn)單、操作方便、安全快速等優(yōu)點(diǎn),但是長(zhǎng)期重復(fù)使用會(huì)導(dǎo)致誘變率降低和抗飽和性[10]。化學(xué)誘變的優(yōu)點(diǎn)是突變頻率較高,但誘變劑量過(guò)高會(huì)有一定毒性,因此會(huì)采用物理-化學(xué)聯(lián)合的方法來(lái)達(dá)到預(yù)期的誘變效果。田強(qiáng)等[11]通過(guò)UV-LiCl復(fù)合誘變獲得一株幾丁質(zhì)酶變異株TUC13,發(fā)酵條件優(yōu)化后酶活提高了236.5%,酶活力為0.11 U/mL。張敏等[12]通過(guò)紫外線、氯化鋰、硫酸二乙酯的復(fù)合誘變,獲得了1株遺傳穩(wěn)定的高產(chǎn)幾丁質(zhì)酶活性菌株,優(yōu)化后酶活力提高到了1.53 U/mL,較出發(fā)菌株提高了347.7%。
本文通過(guò)紫外-LiCl和微波-LiCl兩輪復(fù)合誘變以及發(fā)酵條件優(yōu)化來(lái)提高菌株的產(chǎn)酶能力,為幾丁質(zhì)酶的開(kāi)發(fā)應(yīng)用提供基礎(chǔ)。
1 材料與方法
1.1 材料與儀器
粘質(zhì)沙雷氏菌(Serratiamarcescens)G3-1 從海洋中分離獲得;粘質(zhì)沙雷氏菌GF-21 紫外-LiCl和微波-LiCl復(fù)合誘變后的突變株;細(xì)粉幾丁質(zhì) Sigma公司;胰蛋白胨、酵母粉、瓊脂粉(BR) 上海生物工程有限公司;其余試劑 均為國(guó)產(chǎn)分析純。
U-2910分光光度計(jì)、CF16RXⅡ型高速冷凍離心機(jī) 日本HITACHI公司;78-1型磁力攪拌器 常州國(guó)華電器有限公司;SW-CJ-2F I類(lèi)B型潔凈工作臺(tái) 蘇州安泰空氣技術(shù)有限公司;WP800TL23-K3微波爐 廣東格蘭仕集團(tuán)有限公司;UB-7型pH計(jì) 賽多利斯儀器有限公司。
1.2 實(shí)驗(yàn)方法
1.2.1 培養(yǎng)基 篩選培養(yǎng)基:2%(w/v)膠體幾丁質(zhì)500 mL,K2HPO40.7 g,KH2PO40.3 g,MgSO40.5 g,FeSO4·H2O 0.02 g,NaCl 5 g,蛋白胨3 g,蒸餾水定容至1000 mL,pH7.0~7.2,121 ℃,20 min滅菌;
種子培養(yǎng)基:蛋白胨10 g,牛肉膏3 g,NaCl 5 g,蒸餾水定容至1000 mL,pH7.0~7.2;
基礎(chǔ)發(fā)酵培養(yǎng)基[13]:2%(w/v)膠體幾丁質(zhì)500 mL,蛋白胨10 g,K2HPO40.7 g,KH2PO40.3 g,MgSO40.5 g,FeSO4·H2O 0.02 g,NaCl 5 g,蒸餾水定容至1000 mL,pH7.0~7.2。
1.2.2 出發(fā)菌株G3-1生長(zhǎng)曲線的測(cè)定 將菌株G3-1經(jīng)活化后轉(zhuǎn)接于種子培養(yǎng)基中,30 ℃、200 r/min培養(yǎng),每隔2 h取菌液測(cè)定OD600并記錄。
1.2.3 幾丁質(zhì)酶活力的測(cè)定
1.2.3.1 N-乙酰-D-氨基葡萄糖標(biāo)準(zhǔn)曲線的繪制 準(zhǔn)確稱(chēng)取0.2 g烘干至恒重的N-乙酰-D-氨基葡萄糖,用蒸餾水定容至100 mL,再取出10 mL定容至100 mL,配制成終濃度為0.2 mg/mL的N-乙酰-D-氨基葡萄糖標(biāo)準(zhǔn)溶液,采用DNS法測(cè)定[9,14],按表1加入試劑,混勻后沸水浴5 min,冷水冷卻至室溫,在540 nm下測(cè)定吸光值,根據(jù)吸光值與質(zhì)量的關(guān)系繪制標(biāo)準(zhǔn)曲線,得到回歸方程為y=5.3327x-0.0656,R2=0.9991。
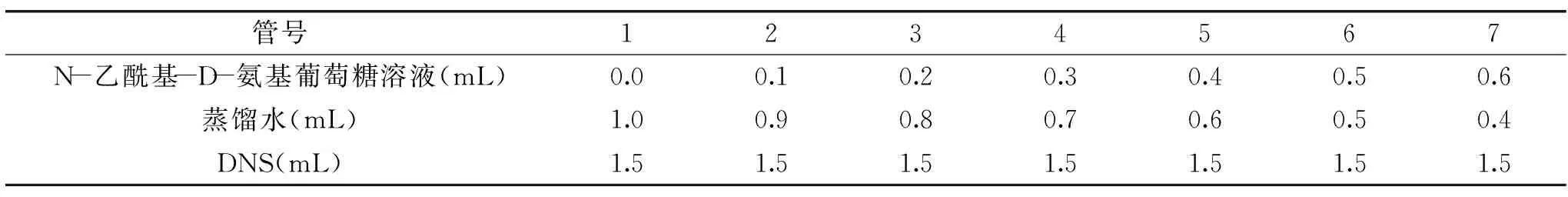
表1 N-乙酰基-D-氨基葡萄糖標(biāo)準(zhǔn)曲線繪制Table 1 The standard curve of N-Acetyl-D-glucosamine
1.2.3.2 酶活力的測(cè)定 采用DNS(3,5-二硝基水楊酸)法測(cè)定還原糖含量[9,14]。取0.1 mL酶液與0.9 mL的0.5%膠體幾丁質(zhì)底物,于50 ℃水浴反應(yīng)30 min后,加入1.5 mL DNS終止反應(yīng)并煮沸5 min顯色;同時(shí)以滅活酶液組作為空白對(duì)照。6000 r/min離心5 min,取上清液于540 nm測(cè)定吸光值,根據(jù)N-乙酰-D-氨基葡萄糖標(biāo)準(zhǔn)曲線計(jì)算還原糖含量。酶活力單位定義為:在該反應(yīng)條件下,1 min催化底物產(chǎn)生1 μmol N-乙酰-D-氨基葡萄糖所需的酶量作為1個(gè)酶活力單位(U)。
1.2.4 紫外-LiCl復(fù)合誘變 取對(duì)數(shù)生長(zhǎng)期菌液離心(10000 r/min,5 min)后,沉淀用生理鹽水稀釋成濃度為108個(gè)/mL的菌懸液。將裝有5 mL菌懸液的無(wú)菌平皿置于磁力攪拌器上,在實(shí)驗(yàn)前打開(kāi)紫外燈預(yù)熱10 min,在距離紫外燈(15 W)30 cm處進(jìn)行誘變。分別照射0、3、6、9、15、30、45 s,每個(gè)照射時(shí)間各取0.1 mL處理后的菌液涂布于篩選平板上,30 ℃避光培養(yǎng)5 d,計(jì)算致死率,確定最佳紫外照射時(shí)間[11]。
致死率(%)=(未經(jīng)誘變處理菌落數(shù)-誘變處理后菌落數(shù))/未經(jīng)誘變處理菌落數(shù)×100
式(1)
將經(jīng)過(guò)紫外誘變后的菌懸液涂布在含有0.5% LiCl的篩選平板上,置于30 ℃培養(yǎng)。
1.2.5 微波-LiCl復(fù)合誘變 以紫外-LiCl復(fù)合誘變的突變株G-12作為微波誘變的出發(fā)菌株。將裝有5 mL菌懸液的無(wú)菌試管置于微波爐中進(jìn)行誘變處理,微波輻射采用高檔火(最大功率700 W,微波波段2450 MHz),分別輻射0、15、30、45、60、75、90、120、180 s,并每隔3 s用冰浴法消除熱效應(yīng)。每個(gè)輻射時(shí)間分別吸取0.1 mL處理后的菌懸液涂布于篩選平板上,30 ℃培養(yǎng)5 d,根據(jù)公式(1)計(jì)算致死率,確定最佳微波輻射時(shí)間[15]。
將經(jīng)過(guò)微波誘變后的菌懸液涂布在含有0.5% LiCl的篩選平板上,置于30 ℃培養(yǎng)。
1.2.6 培養(yǎng)基成分對(duì)突變株GF-21產(chǎn)酶的影響 培養(yǎng)基初始pH7.0,溫度30 ℃,搖床轉(zhuǎn)速200 r/min,接種齡8 h,接種量8%,在此發(fā)酵條件下,對(duì)培養(yǎng)基成分進(jìn)行優(yōu)化。
1.2.6.1 碳源對(duì)產(chǎn)酶的影響 在基礎(chǔ)發(fā)酵培養(yǎng)基中分別添加5 g/L的環(huán)糊精、淀粉、乳糖、蔗糖、果糖、葡萄糖、糯米粉、馬鈴薯淀粉、麩皮作為碳源,以基礎(chǔ)發(fā)酵培養(yǎng)基作為對(duì)照組,研究不同碳源對(duì)GF-21產(chǎn)酶的影響,確定最適碳源。
向基礎(chǔ)培養(yǎng)基中分別添加1、2、3、4、5、6、7 g/L的乳糖,確定乳糖添加量。
1.2.6.2 氮源對(duì)產(chǎn)酶的影響 在添加6 g/L乳糖的基礎(chǔ)上,分別以5 g/L的(NH4)2SO4、NH4Cl、NH4HPO3、NH4NO4、花生餅粉、玉米粉、尿素、蛋白胨、酵母膏取代基礎(chǔ)發(fā)酵培養(yǎng)基中的氮源,以添加6 g/L乳糖的基礎(chǔ)發(fā)酵培養(yǎng)基作為對(duì)照組,研究不同氮源對(duì)GF-21產(chǎn)酶的影響,確定最適氮源[13]。
向基礎(chǔ)培養(yǎng)基中分別添加2、4、6、8、10、12、14 g/L的尿素,確定尿素添加量。
1.2.6.3 金屬離子對(duì)產(chǎn)酶的影響 在添加6 g/L乳糖和8 g/L尿素的基礎(chǔ)上,分別添加0.2 mmol/L和1 mmol/L的K+、Ca2+、Fe3+、Fe2+、Mn2+、Ni+、Zn2+、Cu2+、Mg2+和10 mmol/L的K+、Ca2+,研究不同金屬離子對(duì)GF-21產(chǎn)酶的影響,確定對(duì)產(chǎn)酶促進(jìn)作用最明顯的金屬離子。
向基礎(chǔ)培養(yǎng)基中分別添加0.2、0.4、0.6、0.8、1.0、1.5 mmol/L的K+,確定K+添加量。
1.2.6.4 其他培養(yǎng)基成分添加量的影響 在添加6 g/L乳糖、8 g/L尿素和0.8 mmol/L K+的基礎(chǔ)上,研究作為誘導(dǎo)劑的膠體幾丁質(zhì)添加量(0、2、4、6、8、10、12、14 g/L)以及作為滲透壓調(diào)節(jié)劑的NaCl添加量(1、2、3、4、5、6、7 g/L)對(duì)GF-21產(chǎn)酶的影響。
1.2.6.5 培養(yǎng)基成分的優(yōu)化 在培養(yǎng)基成分單因素實(shí)驗(yàn)基礎(chǔ)上,選擇乳糖、尿素、KCl、膠體幾丁質(zhì)進(jìn)行4因素4水平L16(45)正交實(shí)驗(yàn)[16-17]。正交實(shí)驗(yàn)設(shè)計(jì)表如2所示。

表2 正交實(shí)驗(yàn)設(shè)計(jì)表Table 2 Orthogonal experimental design
1.2.7 發(fā)酵條件對(duì)突變株GF-21產(chǎn)酶的影響
1.2.7.1 培養(yǎng)基初始pH對(duì)幾丁質(zhì)酶合成的影響 分別設(shè)置優(yōu)化后的發(fā)酵培養(yǎng)基初始pH為4.0、5.0、6.0、7.0、8.0、9.0、10.0、11.0,以8%(v/v)接種量,200 r/min發(fā)酵培養(yǎng)48 h后測(cè)定發(fā)酵液酶活,研究不同初始pH對(duì)GF-21產(chǎn)酶的影響[12]。
1.2.7.2 發(fā)酵溫度對(duì)幾丁質(zhì)酶合成的影響 培養(yǎng)基初始pH為9.0,以8%(v/v)接種量,200 r/min發(fā)酵培養(yǎng)48 h后測(cè)定發(fā)酵液酶活,研究不同發(fā)酵溫度(23、25、27、30、33、35、37 ℃)對(duì)GF-21產(chǎn)酶的影響。
1.2.7.3 搖床轉(zhuǎn)速對(duì)幾丁質(zhì)酶合成的影響 培養(yǎng)基初始pH為9.0,以8%(v/v)接種量,30 ℃發(fā)酵培養(yǎng)48 h后測(cè)定發(fā)酵液酶活,研究不同搖床轉(zhuǎn)速(150、180、200、220、250 r/min)對(duì)GF-21產(chǎn)酶的影響。
1.2.7.4 接種齡對(duì)幾丁質(zhì)酶合成的影響 培養(yǎng)基初始pH為9.0,以8%(v/v)接種量,30 ℃,200 r/min發(fā)酵培養(yǎng)48 h后測(cè)定發(fā)酵液酶活,研究種子液不同菌齡(3、4、5、6、7、8、9 h)對(duì)GF-21產(chǎn)酶的影響。
1.2.7.5 接種量對(duì)幾丁質(zhì)酶合成的影響 培養(yǎng)基初始pH為9.0,30 ℃,200 r/min發(fā)酵培養(yǎng)48 h后測(cè)定發(fā)酵液酶活,研究不同接種量(2%、3%、4%、5%、6%、7%、8%、9%、10%、11%、12%、13%、14%)對(duì)GF-21產(chǎn)酶的影響。
1.3 數(shù)據(jù)處理
實(shí)驗(yàn)中每個(gè)樣品設(shè)3個(gè)平行,采用Origin 8.0軟件作圖和SPSS 20.0軟件進(jìn)行數(shù)據(jù)的顯著性分析。
2 結(jié)果與分析
2.1 出發(fā)菌株G3-1生長(zhǎng)曲線的測(cè)定
如圖1所示,菌株G3-1在4 h后進(jìn)入對(duì)數(shù)生長(zhǎng)期,10 h后進(jìn)入穩(wěn)定期,選擇培養(yǎng)8 h后的菌液進(jìn)行誘變。

圖1 菌株G3-1的生長(zhǎng)曲線Fig.1 Growth curve of strain G3-1
2.2 紫外-LiCl復(fù)合誘變
如圖2所示,在3~15 s之間,隨著紫外照射時(shí)間的延長(zhǎng),致死率迅速增加,15 s后趨于平衡。在以紫外線作誘變劑時(shí),一般采用致死率為80%~85%左右的劑量,因此選擇最佳紫外誘變時(shí)間為12 s。

圖2 菌株G3-1紫外誘變的致死曲線Fig.2 Lethal curve of UV mutation
經(jīng)紫外-LiCl復(fù)合誘變,獲得15株酶活明顯提高的正突變菌株(表3)。其中突變株G-12產(chǎn)酶能力提高最多,酶活力達(dá)到1.41 U/mL,比出發(fā)菌株G3-1提高了69.9%。選取4株酶活力有較大提高的突變菌株GU-4、G-12、G-16、GU-17在活化平板上連續(xù)傳代5次,測(cè)定其遺傳穩(wěn)定性。

表3 紫外-LiCl復(fù)合誘變突變菌株的幾丁質(zhì)酶活力提高率Table 3 Improvement rate of chitinase activity of strains by UV-LiCl mutagenesis
由表4可以看出,突變株G-12和G-16的遺傳穩(wěn)定性較好,由于在傳代實(shí)驗(yàn)中G-12的第一代酶活略高些,因此選擇突變株G-12作為微波-LiCl復(fù)合誘變的出發(fā)菌株。
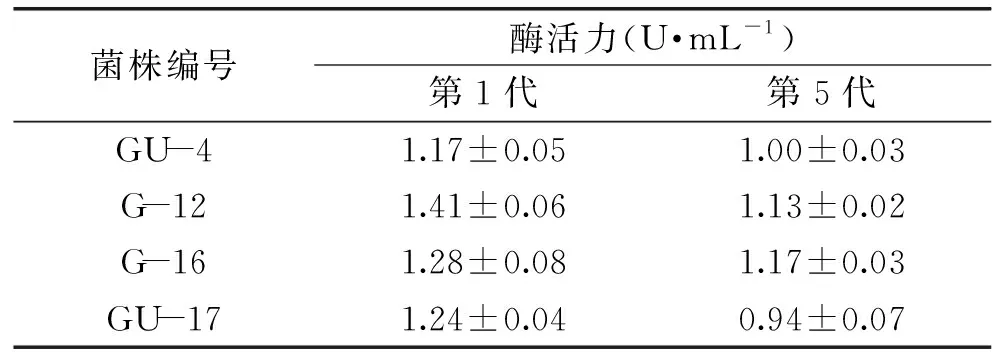
表4 紫外-LiCl復(fù)合誘變突變菌株遺傳穩(wěn)定性Table 4 The genetic stability of mutant strains
2.3 微波-LiCl復(fù)合誘變
如圖3所示,微波輻射時(shí)間在0~60 s之間,微波對(duì)G-12的影響較小,60 s時(shí)致死率僅為15.31%;但在60~75 s之間細(xì)菌迅速死亡,這可能是由于微波輻射熱效應(yīng)的積累對(duì)G-12影響導(dǎo)致的。最后選擇致死率在75%左右的微波輻射時(shí)間75 s作為最佳誘變劑量。

圖3 G-12微波誘變的致死曲線Fig.3 Lethal curve of microwave mutation
經(jīng)微波-LiCl復(fù)合誘變,獲得9株酶活力提高較多的正突變菌株(表5)。選取酶活力提高最多的4株突變株GF-21、GF-25、GF-28、GF-32在活化平板上連續(xù)傳代5次,測(cè)其遺傳穩(wěn)定性,結(jié)果如圖4所示。4株突變株都有較好的遺傳穩(wěn)定性,結(jié)合4株突變株的酶活力,選擇酶活力提高率為87.31%的突變株GF-21作為后續(xù)研究的實(shí)驗(yàn)菌株。

表5 微波-LiCl復(fù)合誘變突變菌株的幾丁質(zhì)酶活力提高率Table 5 Improvement rate of chitinase activity of strains by microwave-LiCl mutagenesis
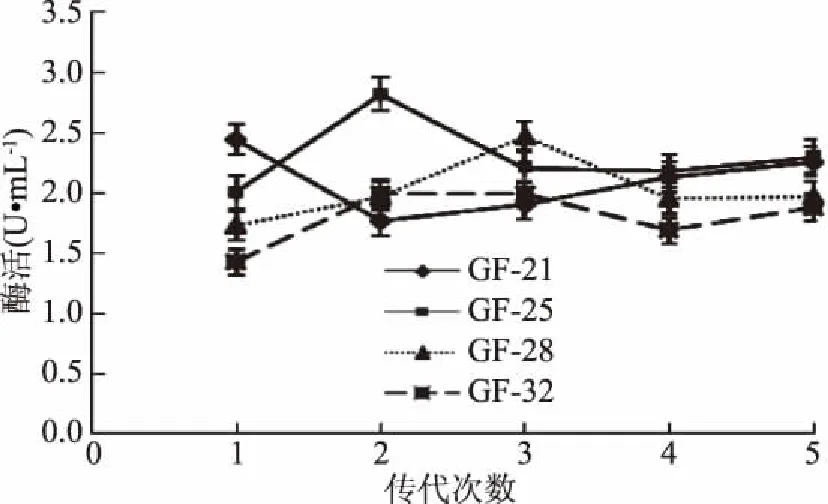
圖4 微波-LiCl誘變突變菌株的遺傳穩(wěn)定性Fig.4 The genetic stability of mutant strains
2.4 培養(yǎng)基成分對(duì)突變株GF-21產(chǎn)酶的影響
2.4.1 碳源對(duì)GF-21產(chǎn)酶的影響 碳源為微生物細(xì)胞物質(zhì)合成提供碳骨架成分,不同碳源對(duì)幾丁質(zhì)酶的影響結(jié)果表明,以乳糖作為碳源,GF-21的酶活力最高,說(shuō)明其效果最好(圖5),當(dāng)乳糖添加量為6 g/L時(shí),產(chǎn)酶能力達(dá)到最高(圖6),因此確定乳糖適宜添加量為6 g/L。
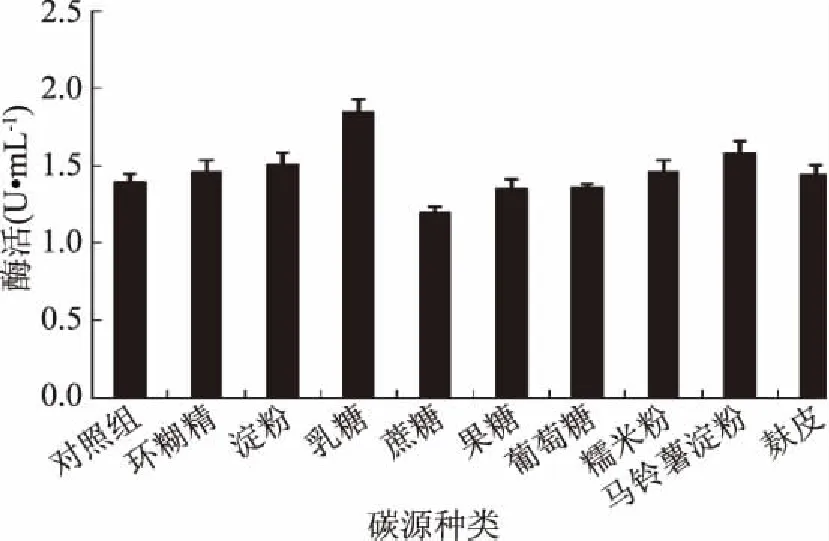
圖5 碳源種類(lèi)對(duì)幾丁質(zhì)酶合成的影響Fig.5 Effects of carbon sourceson chitinase production

圖6 乳糖添加量對(duì)幾丁質(zhì)酶合成的影響Fig.6 Effects of lactose addition on chitinase production
2.4.2 氮源對(duì)GF-21產(chǎn)酶的影響 氮源是微生物合成核酸和蛋白質(zhì)的重要物質(zhì),不同氮源對(duì)幾丁質(zhì)酶的影響結(jié)果表明,尿素對(duì)產(chǎn)酶影響最大(圖7),當(dāng)尿素添加量為2~6 g/L時(shí),GF-21產(chǎn)酶能力隨著尿素添加量的增加而緩慢增加;當(dāng)尿素添加量超過(guò)12 g/L時(shí),產(chǎn)酶能力迅速下降(圖8)。因此確定尿素的添加量為8 g/L。

圖7 不同氮源對(duì)幾丁質(zhì)酶合成的影響Fig.7 Effects of nitrogen sources on chitinase production
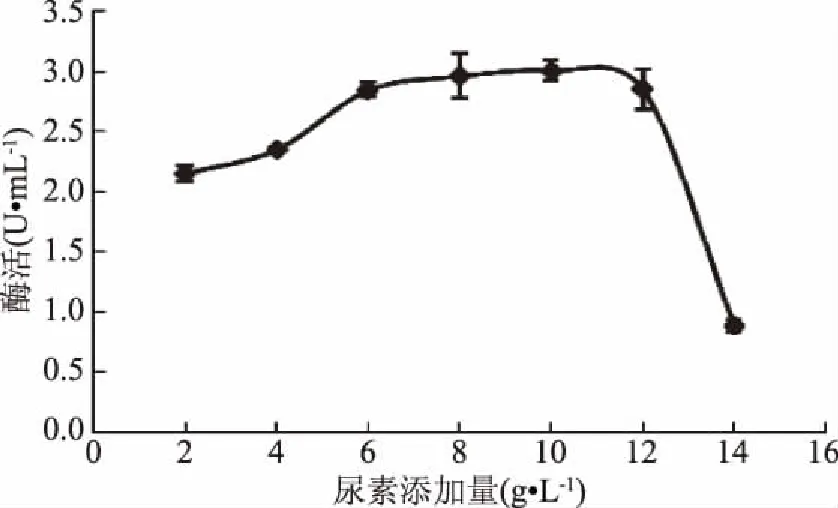
圖8 尿素添加量對(duì)幾丁質(zhì)酶合成的影響Fig.8 Effects of urea addition on chitinase production
2.4.3 金屬離子對(duì)GF-21產(chǎn)酶的影響 金屬離子參與了細(xì)胞內(nèi)結(jié)構(gòu)物質(zhì)和某些酶的活性基團(tuán)的組成,并且影響細(xì)胞的跨膜運(yùn)輸方式。結(jié)果表明,K+對(duì)產(chǎn)酶有較大的促進(jìn)作用,而Cu2+對(duì)產(chǎn)酶幾乎100%抑制,Ni+的抑制效果次之,而Ca2+、Mn2+、Zn2+、Mg2+、Fe2+對(duì)GF-21產(chǎn)酶幾乎沒(méi)有影響(圖9)。

圖9 不同金屬離子對(duì)幾丁質(zhì)酶合成的影響Fig.9 Effects of different metal ions on chitinase production
選擇對(duì)產(chǎn)酶促進(jìn)作用最明顯的K+進(jìn)行添加量實(shí)驗(yàn),結(jié)果如圖10所示。產(chǎn)酶能力隨K+濃度增加而增加,在0.8 mmol/L時(shí)GF-21產(chǎn)酶能力最高,之后趨于平衡,故選擇K+濃度為0.8 mmol/L。

圖10 K+添加量對(duì)幾丁質(zhì)酶合成的影響Fig.10 Effects of K+ addition on chitinase production
2.4.4 其他培養(yǎng)基成分添加量對(duì)GF-21產(chǎn)酶的影響 膠體幾丁質(zhì)作為GF-21發(fā)酵產(chǎn)酶過(guò)程中的誘導(dǎo)劑,同時(shí)提供碳源和氮源,膠體幾丁質(zhì)添加量對(duì)GF-21產(chǎn)酶有較大影響。如圖11所示,當(dāng)膠體幾丁體添加量為10 g/L時(shí),GF-21產(chǎn)酶能力達(dá)到最高;當(dāng)膠體幾丁質(zhì)大于12 g/L,GF-21產(chǎn)酶能力下降,因?yàn)槟z體幾丁質(zhì)濃度太高導(dǎo)致發(fā)酵液變得粘稠,不利于溶氧,抑制細(xì)菌生長(zhǎng)。因此選擇膠體幾丁質(zhì)添加量為10 g/L。

圖11 膠體幾丁質(zhì)添加量對(duì)幾丁質(zhì)酶合成的影響Fig.11 Effects of colloid chitin concentration addition on chitinase production
因出發(fā)菌株G3-1來(lái)源于海洋,故研究NaCl對(duì)其產(chǎn)酶的影響。結(jié)果表明,當(dāng)NaCl濃度為5 g/L時(shí),GF-21產(chǎn)酶能力達(dá)到最高,增加或減少NaCl濃度,GF-21產(chǎn)酶能力都下降,因此選擇NaCl添加量為5 g/L(圖12)。
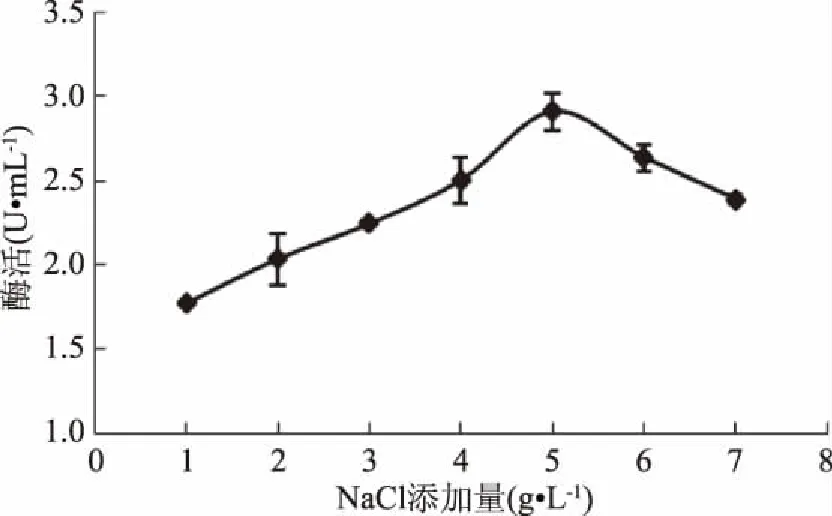
圖12 NaCl添加量對(duì)幾丁質(zhì)酶合成的影響Fig.12 Effects of NaCl addition on chitinase production
2.4.5 正交實(shí)驗(yàn) 在單因素實(shí)驗(yàn)的基礎(chǔ)上,選擇乳糖、尿素、KCl、膠體幾丁質(zhì)進(jìn)行4因素4水平L16(45)正交實(shí)驗(yàn),實(shí)驗(yàn)結(jié)果如表6所示。由極差分析可得各因素對(duì)產(chǎn)酶的影響大小為:

表6 L16(45)正交實(shí)驗(yàn)結(jié)果Table 6 Results of L16(45)orthogonal experiment
D(膠體幾丁質(zhì))>A(乳糖)>B(尿素)>C(KCl);通過(guò)比較K值大小確定最優(yōu)水平組合為A2B4C3D4,即乳糖6 g/L,尿素10 g/L,KCl 1.0 mmol/L,膠體幾丁質(zhì)10 g/L。在該條件下進(jìn)行驗(yàn)證實(shí)驗(yàn),酶活力為4.73 U/mL,比優(yōu)化前提高了109.3%。由方差分析可知(表7),膠體幾丁質(zhì)對(duì)產(chǎn)酶影響極顯著(p<0.01),其次為尿素和乳糖(p<0.1),KCl影響不顯著(p>0.1)。

表7 方差分析表Table 7 Analysis of variance
2.5 發(fā)酵條件對(duì)突變株GF-21產(chǎn)酶的影響
2.5.1 培養(yǎng)基初始pH對(duì)GF-21產(chǎn)酶的影響 當(dāng)培養(yǎng)基初始pH為4.0~9.0時(shí),GF-21產(chǎn)酶能力逐漸增加;當(dāng)pH為9.0時(shí),產(chǎn)酶能力達(dá)到最高。因此選擇培養(yǎng)基初始pH為9.0(圖13)。
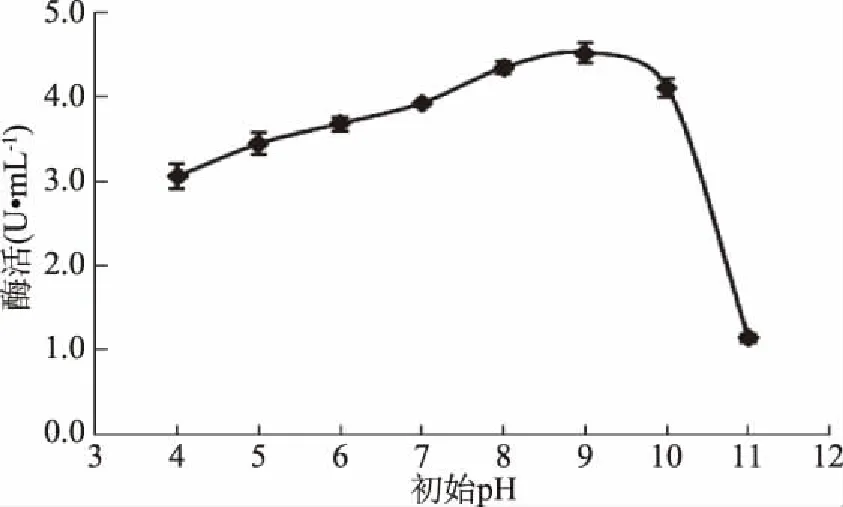
圖13 培養(yǎng)基初始pH對(duì)幾丁質(zhì)酶合成的影響Fig.13 Effects of initial pH on chitinase production
2.5.2 溫度對(duì)GF-21產(chǎn)酶的影響 當(dāng)溫度為25~33 ℃之間時(shí),GF-21產(chǎn)酶能力較高且相對(duì)穩(wěn)定,故選擇30 ℃作為最佳發(fā)酵溫度(圖14)。
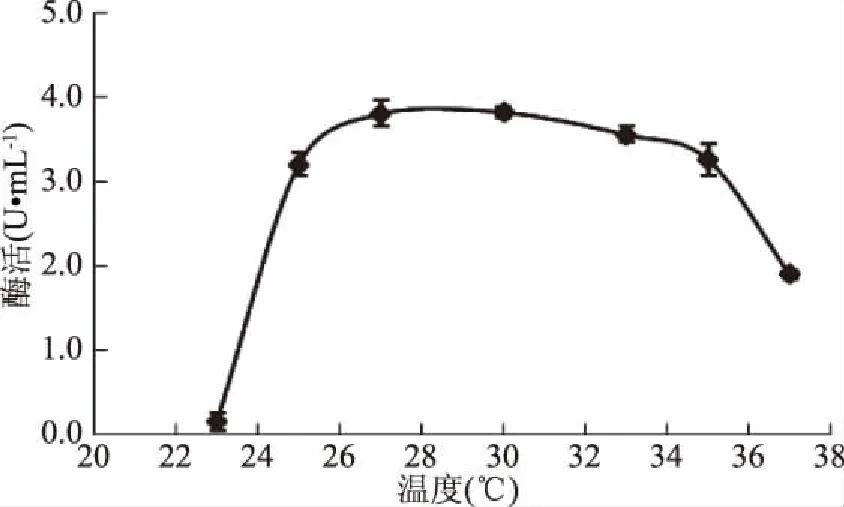
圖14 溫度對(duì)幾丁質(zhì)酶合成的影響Fig.14 Effects of temperature on chitinase production
2.5.3 轉(zhuǎn)速對(duì)GF-21產(chǎn)酶的影響 搖瓶轉(zhuǎn)速影響供氧水平,結(jié)果表明,轉(zhuǎn)速為150~220 r/min時(shí)對(duì)GF-21產(chǎn)酶能力影響不大,但高于220 r/min,產(chǎn)酶能力下降,因此選擇搖瓶轉(zhuǎn)速為220 r/min(圖15)。
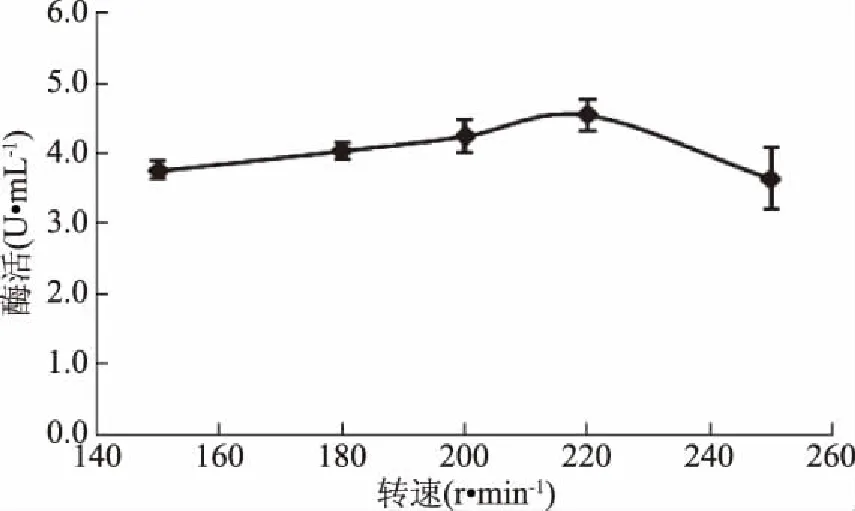
圖15 不同轉(zhuǎn)速對(duì)幾丁質(zhì)酶合成的影響Fig.15 Effects of rotation speed on chitinase production
2.5.4 接種齡和接種量對(duì)GF-21產(chǎn)酶的影響 接種齡和接種量會(huì)影響菌體的生長(zhǎng)周期,但在該實(shí)驗(yàn)中,接種齡和接種量對(duì)GF-21產(chǎn)酶無(wú)明顯影響(圖16,圖17)。處于對(duì)數(shù)生長(zhǎng)期的菌體生長(zhǎng)較快,為了使菌體快速生長(zhǎng)達(dá)到一定的菌濃度,在后續(xù)實(shí)驗(yàn)中,選擇接種齡為5 h,接種量為10%。
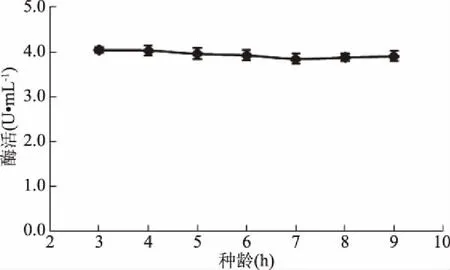
圖16 接種齡對(duì)幾丁質(zhì)酶合成的影響Fig.16 Effects of inoculation age on chitinase production

圖17 不同接種量對(duì)幾丁質(zhì)酶合成的影響Fig.17 Effects of inoculum size on chitinase production
3 結(jié)論與討論
本文以產(chǎn)幾丁質(zhì)酶細(xì)菌G3-1作為出發(fā)菌株,通過(guò)紫外-LiCl和微波-LiCl兩輪復(fù)合誘變,最終獲得一株產(chǎn)酶能力高且遺傳穩(wěn)定的突變株GF-21,并對(duì)其發(fā)酵培養(yǎng)基和產(chǎn)酶條件進(jìn)行優(yōu)化。研究結(jié)果表明,最優(yōu)發(fā)酵培養(yǎng)基配方為:乳糖6 g/L,尿素10 g/L,KCl 1.0 mmol/L,NaCl 5 g/L,膠體幾丁質(zhì)10 g/L。在該條件下進(jìn)行驗(yàn)證實(shí)驗(yàn),酶活力為4.73 U/mL,比優(yōu)化前提高了109.3%,較出發(fā)菌株提高了470%。最優(yōu)發(fā)酵條件為:培養(yǎng)基初始pH9.0,溫度30 ℃,搖床轉(zhuǎn)速220 r/min,接種齡5 h,接種量10%。
根據(jù)已報(bào)道的文獻(xiàn),目前幾丁質(zhì)酶活力偏低,限制了幾丁質(zhì)的開(kāi)發(fā)利用。如施騰鑫等[18]從海塘土壤中分離得到一株產(chǎn)幾丁質(zhì)酶的粘質(zhì)沙雷氏菌菌株,酶活力為1.0 U/mL。王慧敏等[19]對(duì)海洋細(xì)菌產(chǎn)幾丁質(zhì)酶條件進(jìn)行優(yōu)化,優(yōu)化后的酶活力為0.63 U/mL。陶勇[20]對(duì)產(chǎn)幾丁質(zhì)酶細(xì)菌發(fā)酵條件進(jìn)行優(yōu)化,在最優(yōu)條件下酶活力為2.68 U/mL。Singh等[21]篩選到的Paenibacillussp. D1幾丁質(zhì)酶活力為0.8 U/mL。Wang等[22]表明在最優(yōu)產(chǎn)酶條件下進(jìn)行發(fā)酵,其酶活力為0.38 U/mL。本文通過(guò)兩輪復(fù)合誘變獲得一株高產(chǎn)幾丁質(zhì)酶的菌株,產(chǎn)酶穩(wěn)定且酶活力(4.73 U/mL)高,為幾丁質(zhì)酶的生產(chǎn)應(yīng)用奠定了良好的基礎(chǔ)。
[1]郝之奎.Chitinolyticbactermeiyuanensis的篩選鑒定及其發(fā)酵產(chǎn)幾丁質(zhì)酶研究[D]. 無(wú)錫:江南大學(xué),2011.
[2]王曉輝. 海洋細(xì)菌Dl-6幾丁質(zhì)酶和幾丁質(zhì)結(jié)合蛋白的生化性質(zhì)與功能研究[D]. 大連:大連理工大學(xué),2016.
[3]Benecke U.Bacilluschitinovoruseien chitin zersetzenden spaltpilz[J]. Bot Ztg,1995,63(8):227.
[4]Li A,Yu K,Liu H,et al. Two novel thermostable chitinase genes from thermophilic fungi:Cloning,expression and characterization[J]. Bioresource Technology,2010,101(14):5546-51.
[5]Singh AK,Chhatpar HS. Purification and characterization of chitinase fromPaenibacillussp. D1[J]. Applied Biochemistry and Biotechnology,2011,164(1):77-88.
[6]Zhang H,Liu M,Tian Y,et al. Comparative characterization of chitinases from silkworm(Bbombyxmori)and bollworm(Helicoverpaarmigera)[J]. Cell Biochemistry Biophysics,2011(61):267-275.
[7]紀(jì)瑩. 殼寡糖對(duì)巨噬細(xì)胞IL-1β、TNF-α基因表達(dá)的影響[D]. 杭州:浙江大學(xué),2002.
[8]楊銘鐸,曲志華,李元瑞,等. 幾丁質(zhì)、幾丁聚糖及其衍生物在食品中的應(yīng)用[J]. 食品科學(xué),2001,1(5):97-99.
[9]García-Fraga B,Silva AFD,López-Seijas J,et al. A novel family 19 chitinase from the marine-derived pseudoalteromonas tunicata ccug 44952t:Heterologous expression,characterization and antifungal activity[J]. Biochemistry Engineering Journal,2015(93):84-93.
[10]施巧琴. 工業(yè)微生物育種學(xué)[M]. 北京:科學(xué)出版社,2003:68-70.
[11]田強(qiáng),張彩,吳子健. 幾丁質(zhì)酶生產(chǎn)菌的誘變育種及搖瓶發(fā)酵條件優(yōu)化[J]. 食品科學(xué),2007,28(9):405-409.
[12]張敏,胡曉,萬(wàn)津瑜,等. 高產(chǎn)幾丁質(zhì)酶的枯草芽孢桿菌誘變育種及發(fā)酵條件研究[J]. 中國(guó)農(nóng)學(xué)通報(bào),2010,26(11):279-283.
[13]Xia JL,Xiong J,Zhang RY,et al. Production of chitinase and its optimization from a novel isolate serratia marcescens,XJ-01[J]. Indian Journal of Microbiology,2011,51(3):301-306.
[14]Silva AFD,García-Fraga B,López-Seijas J,et al. Characterization and optimization of heterologous expression in escherichia coli,of the chitinase encoded by the chia,gene of bacillus halodurans,c-125[J]. Process Biochemistry,2014(49):1622-1629.
[15]張?jiān)撇?劉海濤,趙雅坤. 一株高效石油降解菌的紫外微波復(fù)合誘變改良[J]. 化學(xué)與生物工程,2013,30(7):71-73.
[16]李志西,杜雙奎. 實(shí)驗(yàn)優(yōu)化設(shè)計(jì)與統(tǒng)計(jì)分析[M]. 北京:科學(xué)出版社,2010:143-177.
[17]梁飛豹. 應(yīng)用統(tǒng)計(jì)方法[M]. 北京:北京大學(xué)出版社,2010:274.
[18]施騰鑫,黃秀菁,劉嘉,等. 黏質(zhì)沙雷氏菌胞外幾丁質(zhì)酶的純化及特性[J]. 食品與發(fā)酵工業(yè),2012,38(7):114-119.
[19]王慧敏,祖國(guó)仁,尹璐,等. 響應(yīng)面法優(yōu)化海洋細(xì)菌產(chǎn)幾丁質(zhì)酶培養(yǎng)條件[J]. 中國(guó)釀造,2012,31(2):103-106.
[20]陶勇. 一株產(chǎn)幾丁質(zhì)酶細(xì)菌的鑒定、酶的性質(zhì)及其基因的克隆與表達(dá)[D]. 成都:四川大學(xué),2006.
[21]Singh AK,Chhatpar HS. Purification and characterization of chitinase fromPaenibacillussp. D1[J]. Applied Biochemistry and Biotechnology,2011,164(1):77-88.
[22]Wang K,Yan PS and Cao LX. Chitinase from a novel strain ofserratiamarcescensjpp1 for biocontrol of aflatoxin:molecular characterization and production optimization using response surface methodology[J]. Biomed research international,2014(3):482623.
猜你喜歡
中學(xué)生數(shù)理化·八年級(jí)物理人教版(2022年3期)2022-03-16 05:55:08
當(dāng)代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(shù)(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國(guó)塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(shù)(2016年3期)2016-03-26 07:13:38
知識(shí)經(jīng)濟(jì)·中國(guó)直銷(xiāo)(2016年3期)2016-02-27 16:15:49
現(xiàn)代檢驗(yàn)醫(yī)學(xué)雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學(xué)低年級(jí)版)(2011年3期)2011-01-01 00:00:00
