基于COI基因序列片段的柔突葉蟬屬部分近緣種分子鑒定與系統(tǒng)發(fā)育
2018-06-22 12:42:06姚亞林陳祥盛
山地農(nóng)業(yè)生物學報 2018年2期
姚亞林,陳祥盛,楊 琳*
(1.貴州大學 林學院,貴州 貴陽 550025;2.貴州大學 昆蟲資源開發(fā)利用省級特色重點實驗室,貴州 貴陽 550025;3.貴州大學 昆蟲研究所,貴州 貴陽 550025;4.貴州大學 貴州山地農(nóng)業(yè)病蟲害省級重點實驗室,貴州 貴陽 550025)
葉蟬類昆蟲均屬植食性,不僅刺吸危害植物,而且其刺吸式口器刺吸植物汁液,影響植物的正常發(fā)育;取食產(chǎn)卵過程造成的植物傷痕影響水分和養(yǎng)分運輸;通過交叉寄主傳播植物病毒;排泄物蜜露堵塞植物氣孔影響光合作用;分泌物可為多種霉菌提供培養(yǎng)基導致霉病爆發(fā),是農(nóng)、林、果樹及經(jīng)濟植物的重要害蟲類群之一。
柔突葉蟬屬AbrusDaietZhang,2002屬半翅目Hemiptera,葉蟬科Cicadellide,角頂葉蟬亞科Deltocephalinae。該屬目前已知19種[1],分別為:安龍柔突葉蟬A.anlongensisChen,Yang Li,2012;竹柔突葉蟬A.bambusanusChen,Yang Li,2012;叉莖柔突葉蟬A.bifurcatusDai Zhang,2002;雙枝柔突葉蟬A.biprocessusLi,2011;短板柔突葉蟬A.breviolusDai Zhang,2008;短莖柔突葉蟬A.brevisDai Zhang,2002;凹板柔突葉蟬A.concavelusLi Wang,2006;錐尾柔突葉蟬A.coneusDai Zhang,2002;大明山柔突葉蟬A.damingshanensisXing Li,2014;道真柔突葉蟬A.daozhenensisChen,Yang Li,2012;寬頂側(cè)突柔突葉蟬A.expansivusXing Li,2014;細莖柔突葉蟬A.graciaedeagusLi,2011;衡山柔突葉蟬A.hengshanensisDai Zhang,2002;黃氏柔突葉蟬A.huangiDai Zhang,2002;崀山柔突葉蟬A.langshanensisYang & Chen,2013;雷公山柔突葉蟬A.leigongshanesisLi Wang,2006;武夷柔突葉蟬A.wuyiensisDai Zhang,2002;習水柔突葉蟬A.xishuiensisYang Chen,2013;云山柔突葉蟬A.yunshanensisChen, Yang Li,2012分布于東洋界和古北界。除叉莖柔突葉蟬大明山柔突葉蟬寬頂側(cè)突柔突葉蟬和武夷柔突葉蟬4個種外,其余14種均報道危害竹子。該屬形態(tài)結(jié)構相似,且部分種的雄性外生殖器結(jié)構差異程度不大,這對該類昆蟲的物種鑒定帶來了困難。
目前,研究近似種最有效的方法是結(jié)合形態(tài)和分子數(shù)據(jù)進行分類鑒定。關于葉蟬科的分子鑒定方面,一些類群如角頂葉蟬類Deltocephalus-like,彎鉤葉蟬屬Flexamia,黃翅葉蟬屬Dalbulus的16S rRNA基因序列片段可用于區(qū)分近緣種[2-3],然而有些類群如緬甸安小葉蟬Anakaburmensis,COI基因也具有相當?shù)蔫b定功效[4-5],可見不同動物類群的最適條形碼并非一致。在葉蟬類昆蟲的DNA條形碼研究中,相比于COI基因,16S rRNA基因序列片段較易擴增,常被作為一種有效的分子標記,用于區(qū)分形態(tài)近似種或描述新發(fā)現(xiàn)的種類[6-10],然而,柔突葉蟬屬的分子鑒定相關研究尚未有報道。為此,本文提取擴增了采自竹類的柔突葉蟬屬5個近似種(錐尾柔突葉蟬、道真柔突葉蟬、習水柔突葉蟬、安龍柔突葉蟬和短莖柔突葉蟬,圖A1-E5)COI序列片段,對其進行分子鑒定,同時利用鄰接法(NJ)、最大簡約MP法構建系統(tǒng)發(fā)育樹,并結(jié)合形態(tài)結(jié)構與系統(tǒng)發(fā)育樹對其近緣種的物種分化時間估計進行分析和討論,以期為角頂葉蟬亞科的分類學和系統(tǒng)學提供科學依據(jù)。
1 材料與方法
1.1 實驗材料
本研究所用標本的采集信息詳見表1,錐尾柔突葉蟬、道真柔突葉蟬、習水柔突葉蟬、安龍柔突葉蟬和短莖柔突葉蟬的外部形態(tài)特征及雄性外生殖器如圖1、圖2。樣品冷凍保存于貴州大學昆蟲研究所,雄性外生殖器保存于甘油中,以備查用。5種近緣柔突葉蟬的外部形態(tài)[11]如圖1,其雄性陽莖如圖2。
1.2 試劑及儀器
昆蟲DNA提取試劑盒購于Omega BioTek公司(D0926-01);引物由生物工程(上海)股份有限公司合成。Olympus SZ2-ILST型光學體視顯微鏡(德國Leica公司);T100TMThermal Cycler型PCR擴增儀(美國BIO-RAD公司);170-8170 型 UVP 凝膠成像系統(tǒng)(美國 BIO-RAD 公司);Power PacTMHV Power Supply型電泳儀(美國BIO-RAD公司);Mini-10K微型高速離心機(珠海黑馬醫(yī)學儀器有限公司);HVE-50型自動高壓滅菌器(日本HIRAYAMA制造公司)等。

表1 供試昆蟲的采集信息Tab.1 Collecting information for the insect specimens
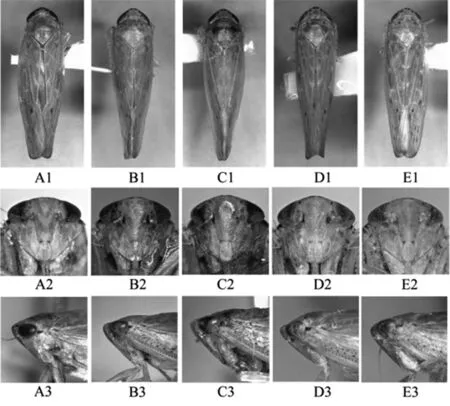
A1- A3:安龍柔突葉蟬A.anlongensis;B1- B3:短莖柔突葉蟬A.brecis;C1- C3:錐尾柔突葉蟬A.coneus;D1- D3:習水柔突葉蟬A.xishuiensis;E1- E3:道真柔突葉蟬A.daozhenensis
圖1 柔突葉蟬屬5個近似種成蟲背面、顏面及頭胸側(cè)面觀
Fig. 1 Habitus of five similar species ofAbrus
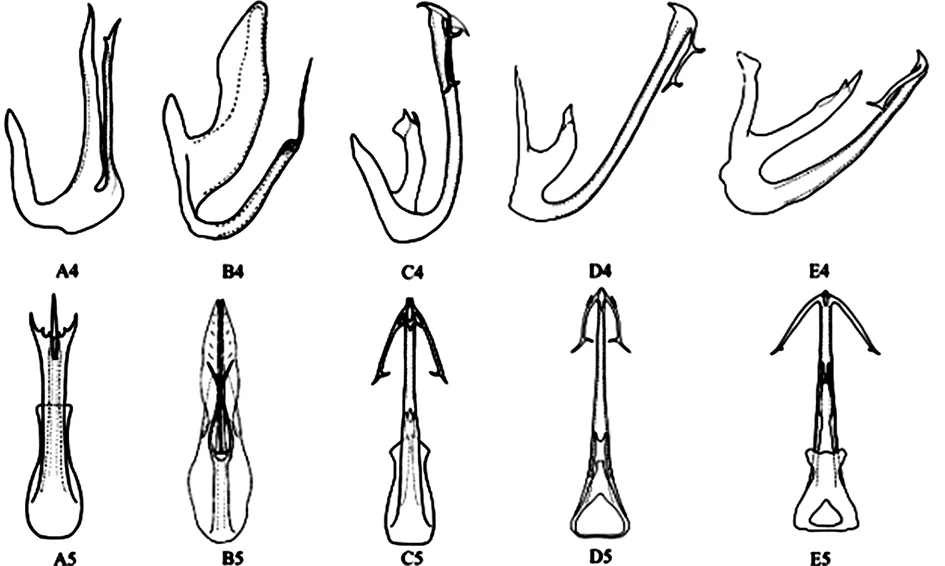
A4- A5:安龍柔突葉蟬A.anlongensis;B4- B5:短莖柔突葉蟬A.brecis;C4- C5:錐尾柔突葉蟬A.coneus;D4- D5:習水柔突葉蟬A.xishuiensis;E4- E5:道真柔突葉蟬A.daozhenensis
圖2 柔突葉蟬屬5個近似種雄性外生殖器特征
Fig. 2 Morphology of male external genitals of five similar species ofAbrus
1.3 DNA提取及PCR擴增
冷凍保存標本解凍后,將腹部放入甘油中保存,剩余的蟲體取頭胸足等部分研磨至粉末狀,然后用Omega BioTek公司生產(chǎn)的E.Z.N.A.TMInsect DNA kit試劑盒提取總DNA,產(chǎn)物-20℃保存。
PCR上下游擴增引物參照Folmer等[5]設計,COI片段引物為:5′-GGTCAACAAATCATAAAGATATTG-3′;5′-TAAACTTCAGGGTGACCAAAAAAT-3′。PCR反應體系為30 μL,其中模板DNA 3 μL,上下游引物各1 μL、Taq PCR Master Mix 15 μL、ddH2O補至30 μL。PCR擴增程序:94℃預變性3 min,94℃變性30 s,50℃退火30 s,72℃延伸1 min,33個循環(huán),最后72℃補償延伸10 min,4℃保存。PCR產(chǎn)物取,3 μL采用1%瓊脂糖凝膠電泳檢測,純化步驟參照DNA凝膠回收試劑盒Omega BioTek公司(D0926-01)程序。回收產(chǎn)物送至生工生物工程(上海)股份有限公司進行雙向測序。
1.4 數(shù)據(jù)分析
將獲得的測序峰圖利用DNAstar 5.0軟件進行正反鏈校對和編輯,手動去除序列兩端的引物區(qū),獲得有效片段的樣本序列。Meglign進行排序后利用Clustal X進行多序列比對,采用DnaSP v5計算信息位點、變異位點數(shù),通過MEGA 6.06分析堿基組成、變異位點,并基于Kimura-2-parameter(K2P)模型進行遺傳距離分析,并以叉突平額葉蟬Flatfrontapronga為外群,采用鄰接法(Neighbor-Joining,NJ法)和最大簡約法(MP法)構建系統(tǒng)發(fā)育樹,系統(tǒng)發(fā)育樹的節(jié)點支持率采用自展值進行估計,重復檢驗1000次。同時結(jié)合貝葉斯方法BPP[12-15]和MCMCTREE[16]對這5個近緣種的物種分化時間進行討論。
2 結(jié)果與分析
2.1 堿基組成及序列變異
利用MEGA 6. 06軟件對本研究中獲得的COI基因序列進行比對后,保留622 bp的同源性序列,對其分析結(jié)果顯示:COI序列中,A、T、G、C含量分別是30.5% 、36.8% 、14.7% 、18.0%,A+ T的平均含量為67.3%,G + C的平均含量為32.7%;共檢測到保守位點407個,變異位點215個,簡約信息點90個,自裔位點125個,變異位點約占總序列的34.56%。
2.2 遺傳距離
利用MEGA 6.06軟件計算不同個體的COI基因序列間的遺傳距離(不同模型下的遺傳距離,表3) 。習水柔突葉蟬與道真柔突葉蟬種間的遺傳距離最小為0.011,錐尾柔突葉蟬與道真柔突葉蟬遺傳距離最大為0.095。柔突葉蟬屬與平額葉蟬屬間的遺傳距離為0.69~0.96。從不同模型下的遺傳距離 (表3~7) 可以看出,K2P轉(zhuǎn)換+G遺傳距離與Pdistance+U遺傳距離模型下短莖柔突葉蟬與安龍柔突葉蟬種間遺傳距離最小為0.011,最大為0.103~0.335,此時從遺傳距離界定這兩種較困難。錐尾柔突葉蟬和道真柔突葉蟬間遺傳距離在K2P轉(zhuǎn)換+顛換+G模型下最大,為0.367,而在JC69 +Gamma模型下為0.366,Pdistance+U遺傳距離最小為0.095。表明轉(zhuǎn)換與顛換以及位點的替代速率分布的異質(zhì)性較強,對遺傳距離的估計影響較大,利用COI基因序列間遺傳差異來鑒定柔突葉蟬效率不太高,總體來說,COI基因序列可以暫時被作為該屬近緣種分子鑒定的候選基因序列片段。

表3 供試5種柔突葉蟬COI基因序列的Pdistance遺傳距離Tab.3 Genetic distances under the Pdistance model of COI sequence of five Abrus species
注:右上半三角部分為遺傳距離的標準誤S.E,下同。
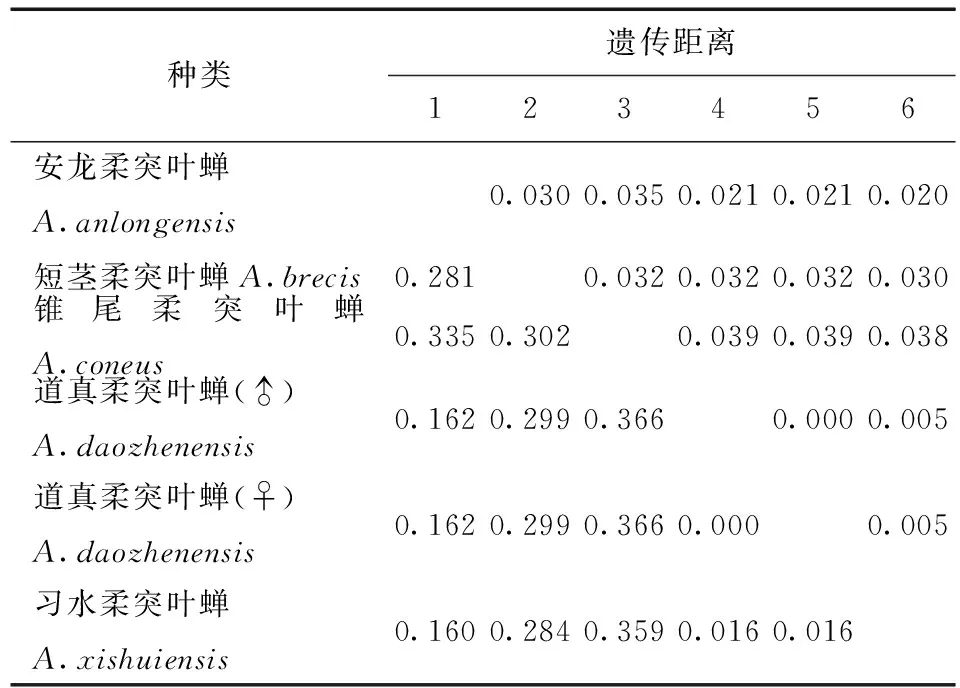
表4 供試5種柔突葉蟬COI基因序列的JC69+G遺傳距離Tab.4 Genetic distances under the JC69+U model of COI sequence of five Abrus species
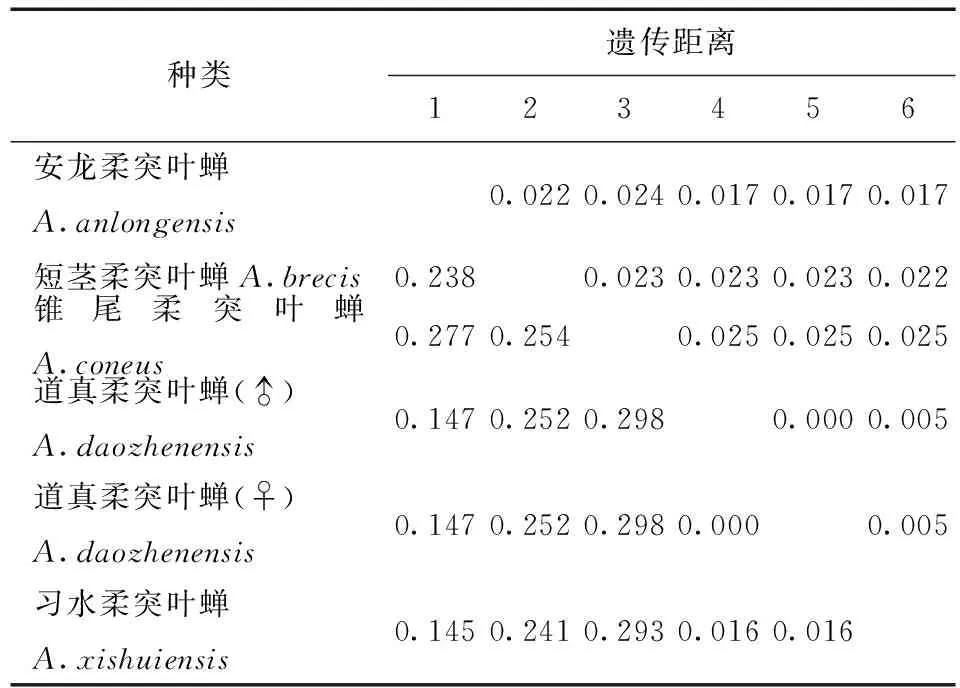
表5 供試5種柔突葉蟬COI基因序列的JC69+U遺傳距離Tab.5 Genetic distances under the K2P+Ts+Tv+G model of COI sequence of five Abrus species

表6 供試5種柔突葉蟬COI基因序列的K2P轉(zhuǎn)換+顛換+G遺傳距離Tab.6 Genetic distances under the K2P+Ts +G model of COI sequence of five Abrus species

表7 供試5種柔突葉蟬COI基因序列的K2P轉(zhuǎn)換+G遺傳距離Tab.7 Genetic distances under the K2P+Ts +G model of COI sequence of five Abrus species
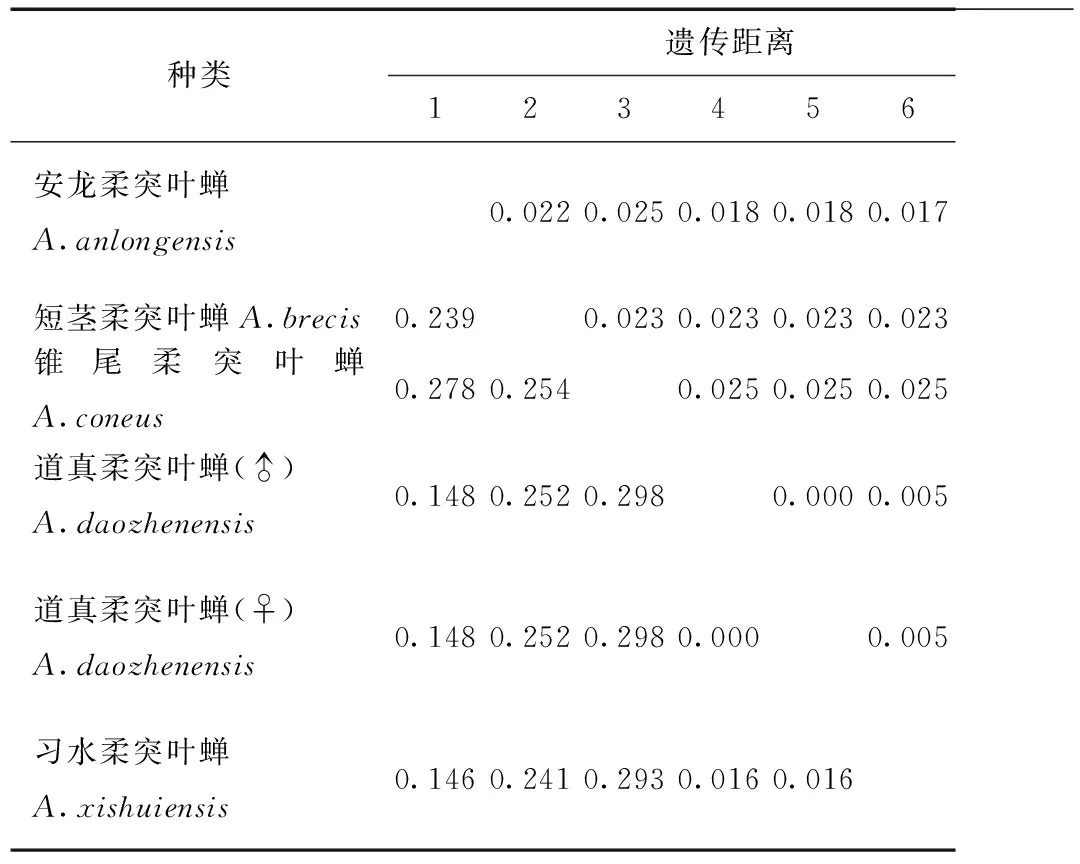
表8 供試5種柔突葉蟬COI基因序列的K2P轉(zhuǎn)換+U遺傳距離Tab.8 Genetic distances under the K2P+Ts +U model of COI sequence of five Abrus species
2.3 堿基替換飽和度分析
本研究使用F84遺傳距離對數(shù)據(jù)組進行了替換飽和分析,結(jié)果表明,轉(zhuǎn)換(S)與顛換(V)的比率均隨著遺傳距離的增加而上升,顛換逐漸多于轉(zhuǎn)換,與線粒體基因中堿基CT及AG突變較多的情況相吻合。轉(zhuǎn)換與顛換的曲線均呈現(xiàn)出向平臺期過渡的趨勢,即出現(xiàn)飽和狀態(tài)的傾向。整體而言,盡管序列整體上較為保守,但構建的數(shù)據(jù)集飽和度在較低的水平上,說明作為常用的分子標記,COI基因包含足夠的遺傳信息可以用于解析本研究所構建數(shù)據(jù)集的分類關系(圖3)。

表9 供試5種柔突葉蟬COI基因序列的Pdistance+U遺傳距離Tab.9 Genetic distances under the Pdistance+U model of COI sequence of five Abrus species
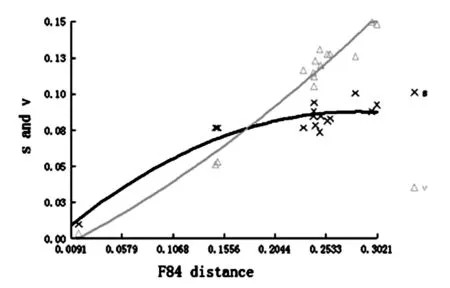
橫坐標:F84模型下的遺傳距離;縱坐標:轉(zhuǎn)換和顛換值
圖3 序列堿基替換飽和度分析
Fig.3 Transitions (s) and transversions (v) versus divergences by COI sequences.
2.4 系統(tǒng)發(fā)育分析
以叉突平額葉蟬為外群,基于COI構建的系統(tǒng)發(fā)育樹中(圖4、5),得到了一致的拓撲結(jié)構,并得到了較高的檢驗支持率。道真柔突葉蟬和習水柔突葉蟬的3個種聚為一支,與安龍柔突葉蟬形成姐妹群關系,短莖柔突葉蟬和錐尾柔突葉蟬位于較根部,其中道真柔突葉蟬與習水柔突葉蟬親緣關系最近,而其與錐尾柔突葉蟬和短莖柔突葉蟬親緣關系較遠。基于貝葉斯方法的BPP物種定界與物種樹估計的結(jié)果(圖6)也表明錐尾柔突葉蟬與道真柔突葉蟬的親緣關系遠,盡管它們的形態(tài)極為相近,但COI構建的系統(tǒng)發(fā)育樹中并未按屬聚一簇,其單系性并不明確。
MCMCTREE的估計結(jié)果中(圖7),短莖柔突葉蟬、錐尾柔突葉蟬、道真柔突葉蟬與習水柔突葉蟬三種在系統(tǒng)樹中的關系與BPP的一致,但其三物種的最近共同祖先(most recent common ancestor,MRCA)分化時間比BPP估計的高,道真柔突葉蟬與習水柔突葉蟬的分化時間約為4-10ka BP左右,而錐尾柔突葉蟬和道真柔突葉蟬+習水柔突葉蟬+安龍柔突葉蟬+錐尾柔突葉蟬的MRCA分化時間約120 ka BP,都是處于全新世初,冰后期。錐尾柔突葉蟬位于根部,這也與其常見寄主差異有關,錐尾柔突葉蟬常見的寄主并非方竹,而是平竹,故在系統(tǒng)發(fā)育樹中體現(xiàn)出與其他柔突葉蟬的差異而出現(xiàn)在根部。
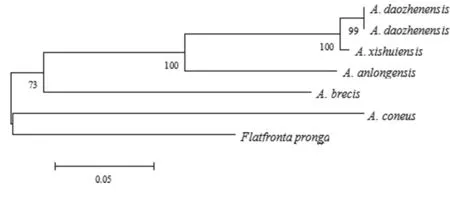
圖4 Abrus 5個近緣種的COI 基因序列進化樹(NJ tree)
Fig.4 eighbor-joining (NJ) analysis of Kimura-2-parameter (K2P) distances of fiveAbrusspecies. Values above the branches are NJ bootstrap values (> 60%).
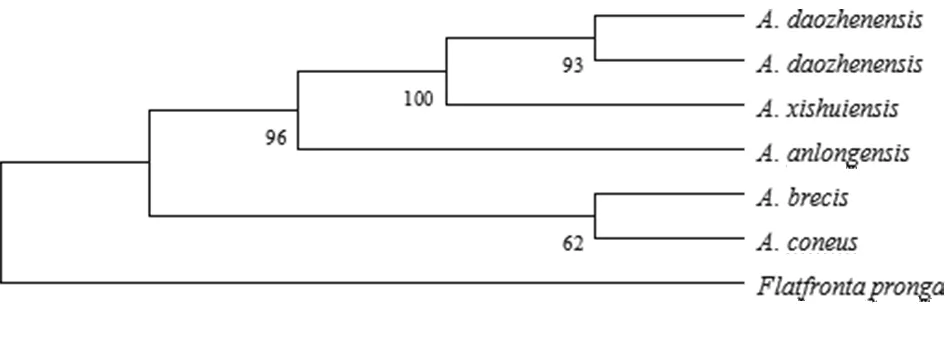
圖5 Abrus 5個近緣種的COI基因序列進化樹(MP tree)
Fig.5 Strict consensus phylogenetic tree based on maximum parsimony (MP) analysis of fiveAbrusspecies. Values above the branches are MP bootstrap values (> 60%).
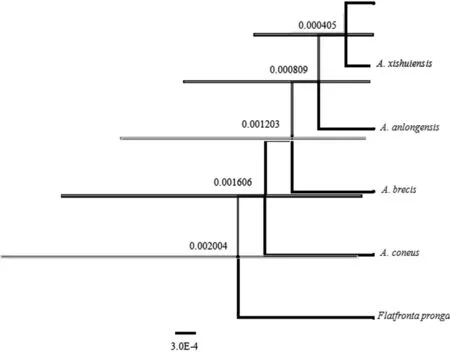
圖6 Abrus 5個近緣種的BPP物種定界
與物種樹估計的最優(yōu)結(jié)果
Fig.6 The optimal results of species delimitation and species tree estimation by BPP of fiveAbrusspecies
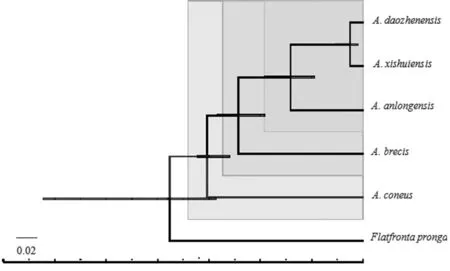
圖7 Abrus 5個近緣種的MCMCTREE物種分化時間估計
Fig.7 Species divergence time of the fiveAbrusspecies by MCMCTREE software
3 結(jié)論與討論
目前已知柔突葉蟬Abrus共19種,全部分布于中國,其形態(tài)相似度高,且大部分寄主為竹類。本研究的5種柔突葉蟬的寄主有貴州都勻斗篷山特有的毛環(huán)方竹Chimonobambusahirtinoda分布,習水的方竹Chimonobambusaangustifolia以及寬闊水有分布的平竹Qiongzhueacommunis和一些其他的方竹,由寄主的獨有性以及柔突葉蟬在世界分布來看,尤其是該屬與方竹共有特有區(qū)的分布特點來看,該屬似乎與竹類的進化有較強的協(xié)同性,但其實際協(xié)同關系還待深入研究。
依據(jù)經(jīng)典分類方法利用雄性外生殖器的形態(tài)學特征對這5種柔突葉蟬進行鑒定時,錐尾柔突葉蟬(C1~C3)與習水柔突葉蟬(D1~D3)道真柔突葉蟬(E1~E3)三者的雄性外生殖器也極為相似(表2),使得在農(nóng)林生產(chǎn)實踐中基于形態(tài)學來區(qū)分這三物種較困難。本研究首次利用COI序列進行分子鑒定,從遺傳距離來看,COI做分子鑒定是可行的,但在不同模型下的距離差異較大可能預示著當樣本數(shù)量增大可能表現(xiàn)并不可觀,今后應該結(jié)合其他基因片段評價其鑒定效率。同時,COI的系統(tǒng)發(fā)育結(jié)果表明錐尾柔突葉蟬和道真柔突葉蟬與習水柔突葉蟬存在很大差異。從形態(tài)來看,其主要區(qū)別不僅在于其翅的外觀色澤、顏面額唇基的飽滿度等特征等,而雄性外生殖器的連鎖形態(tài)、陽莖端部的反向齒突也明顯不同,COI卻能很好的體現(xiàn)這些隱存的差異,F(xiàn)ang等[17]提出“Y”連鎖更具祖征的結(jié)論,本文中錐尾柔突葉蟬和道真柔突葉蟬及習水柔突葉蟬的連鎖卻差異較大,前者是Abrus中唯一一個主干較細的,其余均較粗大(表1),因此出現(xiàn)COI的差異表現(xiàn)并不意外。
分子鑒定理想的條形碼種內(nèi)遺傳差異會小于近緣物種種間差異[18],即種內(nèi)、種間遺傳差異的幅度確定了物種的界限[19]。若種內(nèi)、種間遺傳差異存在重疊,則說明該分子標記無法準確區(qū)分物種。理論上,有效的DNA條形碼應存在條形碼間隔,即種內(nèi)、種間遺傳距離差異明顯。Hebert等[20]提出,種間遺傳差異應達到種內(nèi)遺傳差異的10倍。本研究所涉及種類的COI基因片段序列均有明顯條形碼間隔,未出現(xiàn)種內(nèi)、種間遺傳差異存在重疊的種群,證實了它們作為慈竹葉蟬類昆蟲DNA條形碼標準基因的可行性。盡管有研究表明角頂葉蟬類Deltocephalus-like,彎鉤葉蟬屬Flexamia,黃翅葉蟬屬Dalbulus 16S rRNA基因序列片段可用于區(qū)分近緣種[2-3]。但本文研究結(jié)果表明至少竹類柔突葉蟬的COI基因序列片段被作為一種有效的分子標記,用于區(qū)分形態(tài)近似種。
COI基因的普遍替代速率從0.0168到0.023每個位點/每百萬年[21-22]。真核生物的替代速率從0.00172到0.0258每個位點/每百萬年[23-24]。昆蟲的16S或12S-16S的替代速率從0.005到0.011[22,25]。本研究使用COI的替代速率0.0168到0.023。從MCMCTREE估計的結(jié)果來看(圖7),短莖柔突葉蟬、錐尾柔突葉蟬、道真柔突葉蟬與習水柔突葉蟬的分化時間約為4ka BP左右,而錐尾柔突葉蟬與道真柔突葉蟬與習水柔突葉蟬的祖先分化時間約16ka BP,都是處于全新世初,冰后期,但是估計結(jié)果偏低,這很可能與這些物種的分化早期出現(xiàn)過水平基因流或基因漸滲等復雜演歷程有關,而本文選擇的COI突變頻率的先驗分布盡管是信息較少的先驗,但是數(shù)據(jù)量少使得估計結(jié)果精度低。全新世至近期經(jīng)歷了多次的冷暖波動[26],植被的垂直帶譜逐漸完善的過程中,一些特殊的植物北移,如水青岡等,箭竹等高海拔植物下移[27],而寬闊水國家級自然保護區(qū)正好擁有水青岡-箭竹林群落,同時也具有錐尾柔突葉蟬與道真柔突葉蟬兩種柔突葉蟬昆蟲的分布,這兩種柔突葉蟬的生殖器明顯和貴州雷公山的安龍柔突葉蟬,以及貴州都勻斗篷山的短頸柔突葉蟬的陽莖不同,而這三個地方的方竹自冰后期全新世以來,在經(jīng)歷分化并形成當前的區(qū)系分布格局的進程中,應該對柔突葉蟬屬的分布格局產(chǎn)生重大影響,如果這推斷成立,那么從這種若隱若現(xiàn)而有趣的寄主-昆蟲多樣性聯(lián)系中可以深入分析柔突葉蟬類群的演化及協(xié)調(diào)進化機制,不過今后還需進一步結(jié)合該屬的更多的地理種群樣本從譜系地理學來研究探討更明晰的結(jié)論。
參 考 文 獻:
[1] Zahniser N. 2007-present. An online interactive key and searchable database of Deltocephalinae (Hemiptera: Cicadellidae). http://zahniser.speciesfile.org/
[2] Fang Q, Black C, Blocker D,etal. A phylogeny of New World Deltocephalus-like leafhopper genera based on mitochondrial 16S ribosomal DNA sequences [J].MolecularPhylogeneticsandEvolution, 1993,2 (2):119-131.
[3] Dietrich C, Whitcomb R, Black C. Phylogeny of the grassland leafhopper genus Flexamia (Homoptera: Cicadellidae) based on mitochondrial DNA sequences [J].MolecularPhylogeneticsandEvolution, 1997(8):139-149.
[4] Gaunt W, Miles A. An insect molecular clock dates the origin of the insects and accords with palaeontological and biogeographic landmarks [J].MolecularBiologyandEvolution, 2002,19(5):748-761.
[5] 周寧寧. 基于線粒體 CO I和COⅡ基因的茶園假眼小綠葉蟬地理種群的遺傳分化研究[D]. 杭州:中國計量學院,2014.
[6] Schuchert P. The European athecate hydroids and their medusae (Hydrozoa, Cnidaria): Capitata Part 1 [M]. Revue Suisse de Zoologie, 2006(113):325-410.
[7] Miglietta P, Schuchert P, Cunningham W. Reconciling genealogical and morphological species in a worldwide study of the family Hydractiniidae (Cnidaria, Hydrozoa) [J].ZoologicaScripta, 2009(38):403-430.
[8] Miranda S, Collins G, Marques C. Molecules clarify a Cnidarian life cycle﹣the “Hydrozoan” Microhydrula limopsicola is an early life stage of the Staurozoan Haliclystus antarcticus[J].PLoSONE, 2010(5):e10182.DOI: 10.1371/journal.pone.0010182.
[9] Moura J, Cunha R, Porteiro M,etal. Polyphyly and cryptic diversity in the hydrozoan families Lafoeidae and Hebellidae (Cnidaria: Hydrozoa) [J].InvertebrateSystematics, 2011a(25):454-470.
[10] Moura J, Cunha R, Porteiro M,etal. The use of the DNA barcode gene 16S mRNA for the clarification of taxonomic problems within the family Sertulariidae (Cnidaria, Hydrozoa) [J].ZoologicaScripta, 2011b(40):520-537.
[11] 陳祥盛,楊琳,李子忠. 中國竹子葉蟬[M]. 北京:中國林業(yè)出版社,2012.
[12] Yang Z, Rannala B. Bayesian species delimitation using multilocus sequence data [J].ProceedingsoftheRoyalSocietyB-BiologicalSciences,USA, 2010(107):9264-9269.
[13] Rannala B, Yang Z. Improved reversible jump algorithms for Bayesian species delimitation [J].Genetics, 2013(194):245-253.
[14] Yang Z, Rannala B. Unguided species delimitation using DNA sequence data from multiple loci [J].MolecularBiologyandEvolution, 2014(31):3125-3135.
[15] Rannala B, Yang Z. Efficient Bayesian species tree inference under the multispecies coalescent [J].SystermaticBiology, 2017(66):823-842.
[16] Yang Z. PAML 4: phylogenetic analysis by maximum likelihood. Molecular Biology and Evolution, 2007,24(8):1586-91.
[17] Fang Q, Black C, Bbcker D. Cladistic Analysis of Nearctic Deltocephalus-Like Leafhoppers (Homoptera: Cicadellidea) Using Morphological and Molecular Data [J].AnnualsofEntomologySocietyofAmerica, 1995,88(5): 316-323.
[18] Del-Prado R, Cubas P, Lumbsch T,etal. Genetic distances within and among species in monophyletic lineages of Parmeliceae (Ascomycota) as a tool for taxon delimitation [J].MolecularPhylogeneticsandEvolution, 2010(56):125-133.
[19] K?倠hler F. From DNA taxonomy to barcoding: how a vague idea evolved into a biosystematic tool [J].MitteilungenausdemMuseumfürNaturkundeBerlinZoologischeReihe, 2007(83):44-51.
[20] Hebert N, Ratnasingham S, Waard R. Barcoding animal life: cytochrome c oxidase subunit I divergences among closely related species [J].ProceedingsoftheRoyalSocietyB-BiologicalSciences,USA, 2003(270):S96-S99.
[21] Allegrucci G, Trucchi E, Sbordoni V. Tempo and mode of species diversification in Dolichopoda cave crickets (Orthoptera, Rhaphidophoridae) [J].MolecularPhylogeneticsandEvolution. 2011,60(1): 108-121
[22] Papadopoulou A, Anastasiou I, Vogler P. Revisiting the Insect Mitochondrial Molecular Clock: The Mid-Aegean Trench Calibration [J].MolecularBiologyandEvolution, 2010,27(7): 1659-1672.
[23] Kasuga T, White J, Taylor W. Estimation of nucleotide substitution rates in eurotiomycete fungi [J].MolecularBiologyandEvolution, 2001,19(12): 2318-2324.
[24] Percy M, Kiisiowicz R, Panzera F,etal. Origin and phylogeography of the Chagas disease main vector Triatoma infestans base on nuclear rDNA sequences and genome size[J].InfectionGeneticsandEvolution, 2006(1):46-62.
[25] Brower Z. Rapid Morphological Radiation and Convergence among Races of the Butterfly Heliconius-Erato Inferred from Patterns of Mitochondrial-DNA Evolution [J].ProceedingsoftheRoyalSocietyB-BiologicalSciences,USA, 1994,91(14):6491-6495.
[26] 方修琦,侯光良. 中國全新世氣溫序列的集成重建[J]. 地理科學, 2011,31(4):386-691.
[27] 呂伊娜,熊康寧,容 麗,等. 梵凈山生物生態(tài)演化的世界自然遺產(chǎn)價值對比分析[J]. 世界地理研究, 2016,25(5):131-141.
猜你喜歡
課堂內(nèi)外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
音樂探索(2022年2期)2022-05-30 21:01:37
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
中國特種設備安全(2018年11期)2019-01-08 02:08:32
汽車觀察(2018年10期)2018-11-06 07:05:26
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
