乳酸鈣處理對發芽大豆植酸降解和主要營養物質含量的影響
2018-06-26 09:05:30惠倩汝顧振新楊潤強
食品科學 2018年12期
關鍵詞:大豆
惠倩汝,王 冕,周 婷,王 沛,顧振新,楊潤強*
(南京農業大學食品科學技術學院,南京 江蘇 210095)
植酸又名肌醇六磷酸,廣泛存在于谷、豆類植物籽粒中,是植物磷元素的儲備庫[1]。植酸的化學結構中,含有6 個電負性極強的磷酸基團,具有很強的螯合能力,易與一些金屬離子如Ca2+、Mg2+、Zn2+和Fe2+等形成植酸鹽,與蛋白質結合形成蛋白小體,從而影響人體對礦質離子和蛋白質的吸收。因此,植酸被認為是一種抗營養因子[2]。
發芽被認為是一種低成本、有效降解植酸的技術,同時也可以增強芽苗的營養價值[3]。據El-Adawy[4]報道,發芽相較于水煮、高壓蒸煮與微波蒸煮,對植酸降解最有效。乳酸鈣是一種天然的有機酸鈣鹽,具有極高的生物利用率、可溶性及穩定性,是有效的外源鈣元素補充劑,且不會影響溶液的感官[5]。同時,鈣元素對植物的生長發育至關重要。目前,已有許多植物籽粒發芽過程中加入外源鈣富集功能性成分的報道。Wang Xinkun等[6]報道了6 mmol/L氯化鈣溶液有效提高了大豆芽菜的產量、促進了植酸的降解,同時富集了功能性成分。在豆類籽粒發芽過程中,植酸在植酸酶的作用下降解脫去磷酸基團,既降低了植酸的抗營養作用,也保持了植酸的有益作用,如抗氧化活性等[7]。有報道稱,發芽綠豆中植酸的降解與其子葉中植酸酶活性的升高有密切關系[8]。同時,所形成的低級磷酸肌醇也是重要的信號物質,參與植物體內多種信號通路的傳遞[9],可起到抵御結腸癌、動脈硬化、神經系統疾病及冠心病等功能[10]。大豆是我國常見的糧食作物,其芽菜生產周期短,是廣受國民喜愛的一種蔬菜。然而,大豆植酸含量較高,會影響礦質元素和其他營養物質的吸收。外施乳酸鈣處理可降解大豆芽菜中的植酸,并富集低級磷酸肌醇[11]。但是,對于乳酸鈣處理降解植酸的過程中,大豆芽菜礦物質和主要營養物質含量的變化還鮮見報道。因此,對乳酸鈣處理大豆芽菜營養價值的認識并不全面。
本研究以大豆為試材,研究大豆芽菜的鐵、鋅元素含量、生理生化特性、蛋白質、氨基酸、可溶性糖等營養成分含量變化等,明確乳酸鈣對大豆芽菜營養品質的影響,為高品質大豆芽菜生產提供理論依據。
1 材料與方法
1.1 材料與試劑
大豆(Glycine max L.,品種:云鶴)籽粒購于江蘇省農業科學院,并置于-20 ℃冰柜保藏待用。千粒質量為(161.6±0.8)g。
乳酸鈣 美國Sigma-Aldrich公司;AG1-X4樹脂(100~200 目) 德國Alfa Aesar公司;牛血清白蛋白上海藍季科技發展有限公司;亮氨酸、考馬斯亮藍G-250、茚三酮 國藥集團上海化學試劑有限公司。
1.2 儀器與設備
BX801發芽機 貝欣五金電器廠;755B型分光光度計上海精密科學儀器有限公司;TGL-16GR冷凍離心機、TDL-40B離心機 上海安亭科學儀器廠;PYX-DHS-BS型隔水電熱恒溫培養箱 上海躍進醫療器械廠;L8900AAA-400型氨基酸自動分析儀 日本日立公司;2300全自動凱氏定氮儀 瑞典福斯公司;Optima 2100 DV型電感耦合等離子體發射光譜儀 珀金埃爾默公司。
1.3 方法
1.3.1 樣品處理
每種處理優選200 顆大小一致且無腐爛、霉變的大豆籽粒。種子表面用去離子水洗去浮沉與污漬,后置于體積分數1%的次氯酸鈉溶液浸泡除菌消毒15 min。用去離子水漂洗至少5 次至pH值為中性,洗凈的籽粒在去離子水中于30 ℃浸泡6 h。泡好的籽粒移入自動發芽機中,溫度30 ℃,相對濕度85%,黑暗培養4 d。
用乳酸鈣溶液在大豆籽粒發芽過程中進行噴淋處理。乳酸鈣溶液每小時噴淋1 次,每次2 min。用去離子水噴淋設置為對照。每24 h更換一次培養液。大豆芽菜分別在發芽0、2 d和4 d取樣,取樣后進行用去離子水漂洗,確保樣品表面沒有噴淋液殘留。樣品每5 株隨機分裝一袋,凍干后,分別磨粉,置于-20 ℃冰柜保藏待用。
乳酸鈣濃度優化:設置濃度為0.01、0.03、0.09、0.27、0.30、0.90、1.50、2.10、2.70 mmol/L。取發芽4 d大豆芽苗測定植酸含量,確定最佳降解植酸的乳酸鈣濃度。
1.3.2 各指標的測定
植酸含量測定參照Hui Qianru等[11]的方法。5 株粉末樣品用30 mL 100 g/L的Na2SO4(溶劑為體積分數1.2%的HCl溶液)30 ℃提取2 h。9 000×g離心20 min后取上清液過陰離子交換樹脂,取5 mL濾液加4 mL顯色液(由質量分數0.03%的FeCl3溶液及0.3%的磺基水楊酸配制),靜置10 min,在波長500 nm處測定吸光度。實驗結果以sprout計。
總鈣含量:采用電感耦合等離子體發射光譜測定,參照Hui Qianru等[12]的方法測定。樣品與優級純硝酸按1∶25(g/mL)混合,進行微波消解。消解后稀釋至有效濃度用電感耦合等離子體發射光譜測定。實驗結果以sprout計。
總鋅、鐵含量:參照總鈣含量測定方法。實驗結果以sprout計。總鐵、鋅含量與植酸含量的關系以其物質的量比表示,參照Karkle等[13]的方法計算。
可溶性糖含量:采用硫酸苯酚法測定[14]。總蛋白質含量:采用凱氏定氮法測定[14]。可溶性蛋白質含量:采用考馬斯亮藍G-250法測定[14]。游離氨基酸含量:采用茚三酮溶液顯色法測定[14]。氨基酸組成分析:采用氨基酸自動分析儀測定氨基酸含量[15]。以上實驗結果以sprout計。
1.4 數據統計與分析
實驗設3 次生物學重復,數據以 ±s表示。采用SAS 9.2軟件Duncan’s多重比較法進行顯著性分析,顯著性水平為P值小于0.05。
2 結果與分析
2.1 乳酸鈣濃度篩選
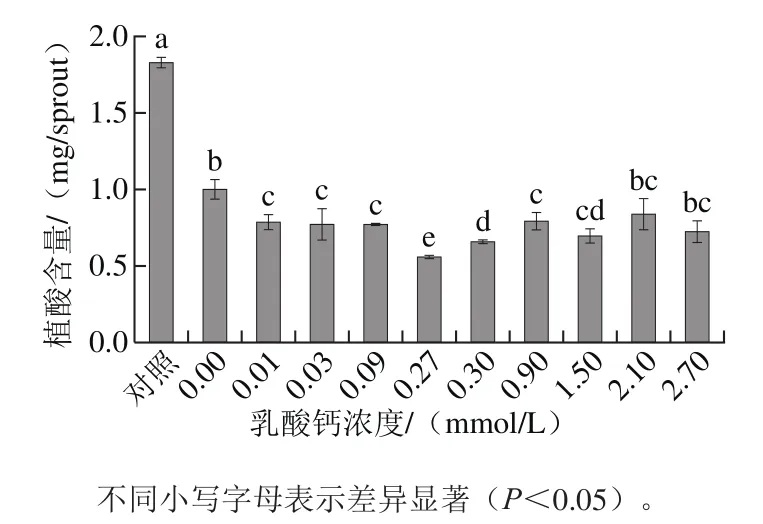
圖1 不同濃度乳酸鈣對發芽4 d大豆芽菜植酸含量的影響Fig.1 Effects of different calcium lactate concentrations on phytic acid content of 4 day-old soybean sprouts
如圖1所示,設發芽0 d大豆為對照,外源乳酸鈣對發芽4 d大豆芽菜的植酸具有顯著的降解作用。其中0.27 mmol/L的乳酸鈣溶液處理條件下植酸含量最低,比去離子水噴淋低44.22%,比未發芽的大豆低69.77%。表明發芽和乳酸鈣噴淋對植酸的降解具有顯著的疊加效果。因此,選0.27 mmol/L的乳酸鈣溶液用于大豆發芽期間噴淋處理。
2.2 總鈣含量測定結果
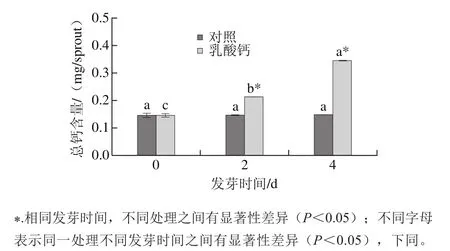
圖2 乳酸鈣處理條件下大豆芽菜總鈣含量的變化Fig.2 Changes in total calcium content with calcium lactate treatment
隨著發芽時間的延長,外源鈣在大豆芽菜中不斷積累,總鈣含量也因此不斷升高。由圖2可知,乳酸鈣處理條件下,大豆芽菜在發芽4 d時的總鈣含量是0 d的1.36 倍,而對照組隨著發芽時間的延長,總鈣含量并無顯著性變化。表明大豆籽粒在發芽過程中,可以吸收外源乳酸鈣,且總鈣含量隨著發芽時間延長不斷增加。
2.3 總鋅、鐵含量及植酸與其物質的量比值測定結果
植酸易與金屬離子鋅鐵螯合形成植酸鹽的形式積累。由圖3可知,相同發芽時間條件下,乳酸鈣處理的芽菜總鋅、總鐵含量并無顯著變化。但是,隨發芽時間的延長,植酸-鋅和植酸-鐵物質的量比值顯著降低。在發芽第4天時,植酸-鋅和植酸-鐵物質的量比值較對照分別降低58.20%和58.75%,而發芽第2天時,乳酸鈣處理與對照無顯著差異。
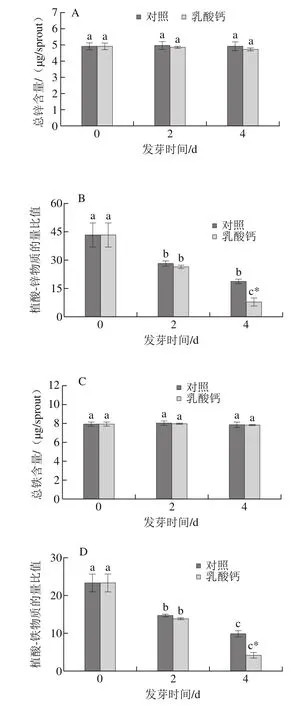
圖3 乳酸鈣處理條件下大豆芽菜總鋅含量(A)、植酸-鋅物質的量比值(B)、總鐵含量(C)及植酸-鐵物質的量比值(D)的變化Fig.3 Changes in total zinc content (A), molar ratio of PA to Zn (B),total iron content (C) and molar ratio of PA to Fe (D) with calcium lactate treatment
2.4 可溶性糖含量測定結果
由圖4可知,隨著發芽時間的延長,可溶性糖含量顯著下降。其中,在乳酸鈣處理條件下,可溶性糖含量顯著低于對照。在發芽4 d時,乳酸鈣處理的大豆芽菜可溶性糖比0 d低45.66%,表明乳酸鈣處理條件下的大豆芽菜對可溶性糖利用更多。
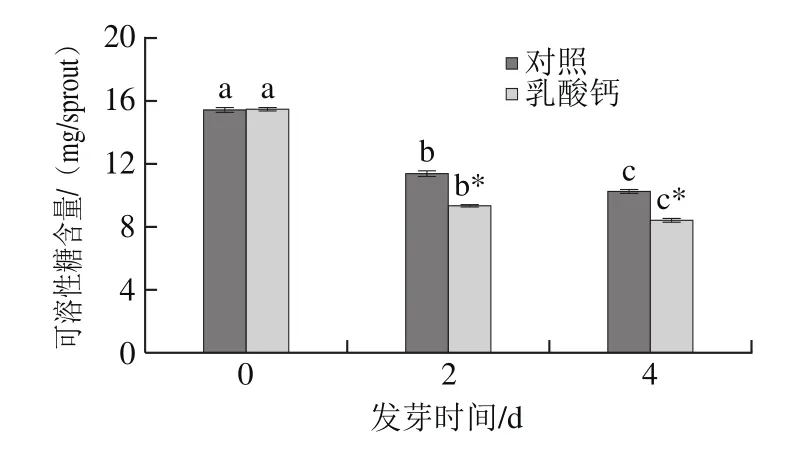
圖4 乳酸鈣處理條件下大豆芽菜可溶性糖含量的變化Fig.4 Changes in soluble sugar content with calcium lactate treatment
2.5 總蛋白質含量測定結果
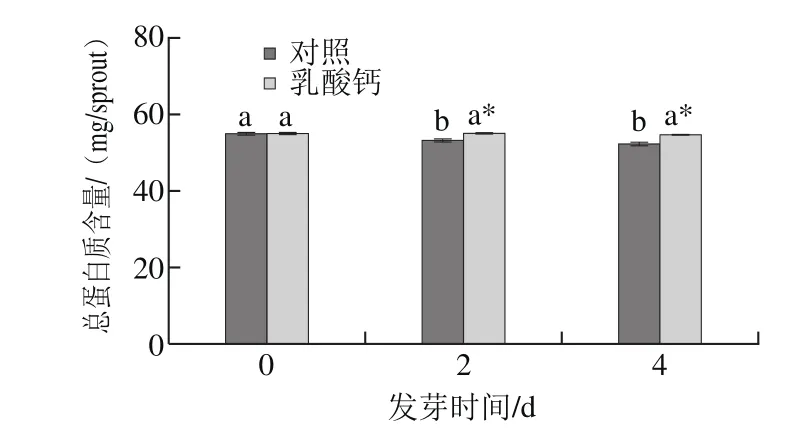
圖5 乳酸鈣處理條件下大豆芽菜總蛋白質含量的變化Fig.5 Changes in total protein content with calcium lactate treatment
蛋白質是大豆籽粒中最重要的營養物質,約占大豆鮮質量的40%左右。如圖5所示,發芽2 d,對照組中總蛋白質含量顯著下降,然而乳酸鈣處理對總蛋白質含量卻無顯著影響。發芽至第4天,對照組的總蛋白含量與乳酸鈣處理組的總蛋白含量相比顯著下降。表明乳酸鈣處理不僅不會造成總蛋白質的流失,而且可以保存蛋白質。
2.6 可溶性蛋白含量測定結果
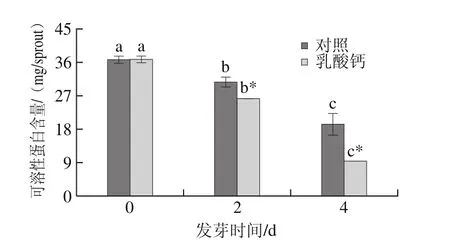
圖6 乳酸鈣處理條件下大豆芽菜可溶性蛋白含量的變化Fig.6 Changes in soluble protein content with calcium lactate treatment
如圖6所示,發芽過程中,對照及乳酸鈣處理均可顯著降低可溶性蛋白的含量,但在相同發芽時間條件下,乳酸鈣處理大豆芽菜的可溶性蛋白含量顯著低于對照。發芽4 d,乳酸鈣處理的可溶性蛋白含量比0 d降低74.56%。
2.7 游離氨基酸含量測定結果
隨著發芽時間的延長,游離氨基酸含量顯著增加。其中,發芽4 d時,乳酸鈣處理的大豆芽菜中游離氨基酸含量比對照增加了30.35%;與0 d相比增加到5.6 倍。這可能與可溶性蛋白的水解有關。
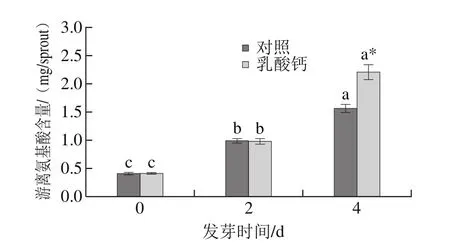
圖7 乳酸鈣處理條件下大豆芽菜游離氨基酸含量的變化Fig.7 Changes in free amino acids with calcium lactate treatment
2.8 氨基酸組成變化
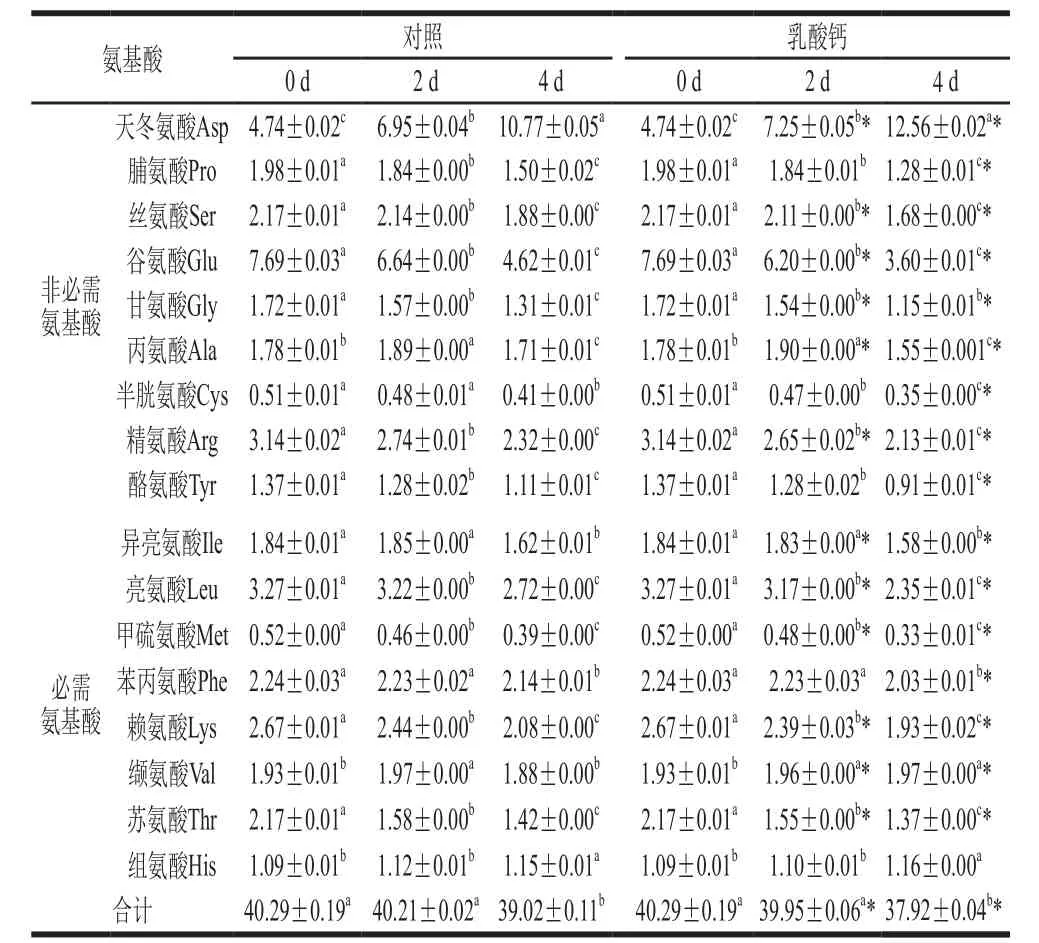
表1 乳酸鈣處理條件下氨基酸組成的變化Table1 Changes in amino acid composition with calcium lactate treatment%
探究在乳酸鈣處理條件下發芽4 d大豆芽菜的17 種氨基酸組成的變化,如表1所示。其中包括8 種必需氨基酸和9 種非必需氨基酸。發芽0 d 時,谷氨酸含量最高,發芽4 d后,其含量比對照和乳酸鈣處理分別下降了39.92%和53.19%。但發芽4 d后,天冬氨酸成為含量最高的氨基酸,與未發芽大豆相比,對照和乳酸鈣處理中天冬氨酸含量分別增加了1.27 倍和1.65 倍。在發芽過程中,乳酸鈣處理與對照均只有天冬氨酸和組氨酸含量隨著發芽時間的延長而增加;丙氨酸含量在發芽第2天達到最高,隨后下降;其余氨基酸含量都呈下降趨勢。但纈氨酸含量在對照中先升高后下降,而在乳酸鈣處理條件下一直上升,因此在4 d與對照相比增加4.79%。乳酸鈣處理條件下,除了天冬氨酸、纈氨酸和組氨酸,其余氨基酸含量均在發芽4 d時顯著低于對照。
3 討 論
鈣是植物的必需元素之一,對其生長發育有著重要的作用[16]。本研究在大豆籽粒發芽過程中施用0.27 mmol/L乳酸鈣處理,有效地降解了植酸(圖1),同時探明了幾種基本營養成分含量的變化。由于乳酸鈣良好的溶解性、穩定性以及生物利用率,其被認為是一種優質的鈣元素補充劑。在農業生產與食品加工中,無機鈣鹽如氯化鈣被廣泛應用,但其用量大濃度高,生物利用率低,難以滿足生產需要。Wang Xinkun[6]和Yin Yongqi[17]等均施用6 mmol/L的氯化鈣降解植酸來提高大豆芽菜品質、緩解鹽脅迫等。但0.27 mmol/L乳酸鈣具有更好的植酸降解效果,能更好地提高大豆芽菜品質。其原因可能是:第一,鈣元素是植物細胞的結構組成元素,鈣元素的補充促進大豆生長;第二,鈣參與能量三磷酸腺苷(adenosinetriphosphate,ATP)的合成,而ATP的合成需要大量的磷元素,因此促進了植酸降解[9];第三,鈣是細胞內的第二信使,是植物體內多種酶類和輔酶的活化因子,可以通過影響酶的活性來調節多種生理過程[18]。同時,乳酸的作用也不容忽視。據報道,乳酸可以有效激活大麥的植酸酶活性,降低大麥中的植酸含量,從而促進其體內低級磷酸肌醇的形成[19]。因此,乳酸鈣的補充適用于植物籽粒在發芽過程中植酸的降解。
在種子萌發過程中,外施乳酸鈣被植物大量吸收(圖2),大豆芽菜生長狀態佳,而芽菜旺盛的生長狀態必然和呼吸速率的增強以及子葉中營養物質大量的消耗相關。此時,植酸降解相關酶被激活[20],植酸降解及其磷代謝等也均受到Ca2+的調控,因此在乳酸鈣處理條件下植酸隨著發芽時間的延長而大量降解[9]。隨之生成低級磷酸肌醇、無機磷、肌醇、二價金屬離子以及蛋白等物質,以供給植株生長所需[21]。總鐵、總鋅含量在發芽過程中基本保持不變,但植酸降解釋放了鐵、鋅元素從而降低了大豆的抗營養作用。在幾種易被植酸螯合的礦質元素中,鋅與植酸形成的植酸鋅是最穩定的,因此鋅元素的生物利用率也是受植酸影響最大的[22]。鐵、鋅與植酸的物質的量之比可有效反映鐵、鋅的吸收情況。據報道,植酸-鋅物質的量比值超過10~15會抑制鋅的吸收;而植酸-鐵物質的量比值超過14會造成鐵缺乏[23-24]。發芽0 d的大豆籽粒的鋅、鐵元素利用率極差,但在乳酸鈣處理條件下,發芽4 d的大豆芽菜植酸-鋅與植酸-鐵物質的量比值均有了很大變化,鋅、鐵元素的吸收也達到了最佳效果,并且顯著優于對照組(圖3B、D)。
碳水化合物約占大豆籽粒干質量的35%左右,多為蔗糖、棉子糖、水蘇糖等可溶性糖及纖維素類[25]。其中可溶性糖是大豆中非結構性碳水化合物的重要組成部分,包括絕大部分單糖和寡糖。而非結構性碳水化合物也是大豆籽粒中重要的能量來源。其含量隨著發芽過程逐漸下降(圖4),且與植株發育程度和生長環境有關。據報道,淀粉作為另一種重要的非結構性碳水化合物,在大豆發芽過程中含量也在不斷下降[26]。蔗糖、淀粉被大量利用,提供發芽過程中所需的能量。
植酸與蛋白質也易結合形成植酸-蛋白質復合體,蛋白質結構發生改變而產生聚沉作用,導致其溶解度降低,同時被蛋白酶水解的程度也下降[27]。在大豆籽粒中,蛋白質是其中最重要的營養物質并占有較大比例。隨著籽粒休眠被打破,蛋白酶被激活,籽粒中貯藏的蛋白質被水解成多肽、氨基酸及酰胺。因此,在發芽過程中可溶性蛋白含量不斷下降(圖6),游離氨基酸含量逐漸增加(圖7),而外施乳酸鈣處理組變化更為明顯,可溶性蛋白消耗更多,游離氨基酸生成也更多。總蛋白含量在乳酸鈣處理發芽過程中基本沒有變化,因為無外源添加的情況下氮元素在植物體內僅被轉化利用,并不影響總含量。
大豆中含有人體所必需的各類氨基酸,其中8 種必需氨基酸的含量與肉制品相似。因此,大豆氨基酸組成平衡,其營養價值高于其他谷豆類植物性食品[28]。氨基酸組成的變化可以從某種程度上反映出蛋白質的轉化情況。本研究共檢測17 種氨基酸的變化,其中有9 種非必需氨基酸以及8 種必需氨基酸。大豆籽粒在未發芽時,谷氨酸含量最高。谷氨酸是一種酸性氨基酸,廣泛存在于谷豆類蛋白質中,是蛋白質代謝的重要中間產物之一。隨著發芽時間的延長,谷氨酸含量逐漸下降,其中乳酸鈣處理的大豆芽菜中下降更為明顯(表1)。可能原因是,在大豆萌發過程中游離谷氨酸的不斷釋放,為谷氨酸脫羧酶(glutamate decarboxylase,GAD)提供了充足的底物。GAD是γ-氨基丁酸(γ-aminobutyric acid,GABA)生成的關鍵酶,伴隨著發芽的進行,GABA也在芽菜體內不斷積累[6]。GAD活性受到Ca2+調控,而植酸降解釋放的Ca2+與外源吸收的Ca2+不斷增加,也因此大大提高GAD活性[29]。據報道,發芽與施鈣會促進大豆芽菜中酚類物質的積累[6]。苯丙氨酸是植物體內酚類物質包括大豆異黃酮的前體[30],在乳酸鈣處理發芽過程中,苯丙氨酸含量也可能因此而不斷下降。與谷氨酸、苯丙氨酸不同的是,天冬氨酸含量顯著升高,而施加乳酸鈣的處理組升高更為明顯(表1)。天冬氨酸是一種非必需氨基酸,也是數種氨基酸合成的前體,包括必需氨基酸甲硫氨酸、蘇氨酸、異亮氨酸、賴氨酸等。因此,在籽粒發芽過程中天冬氨酸要不斷積累,供給大豆生長發育之用。鈣的施用,提高了生化反應速率,也加速了天冬氨酸的合成。另外,天冬氨酸也具有和Ca2+結合的能力,大豆芽菜吸收了大量Ca2+后,更多的天冬氨酸生成,可以更好地在芽體內貯藏、分流多余鈣源。組氨酸是另一種伴隨發芽過程含量上升的氨基酸,發芽4 d,乳酸鈣處理組和對照組組氨酸均顯著高于未發芽大豆的含量(表1)。組氨酸是嬰兒的必需氨基酸,因此富含鈣的芽類食品也可作為嬰兒輔食進行進一步開發。
在大豆發芽過程中施用0.27 mmol/L乳酸鈣處理可以顯著降解大豆籽粒體內的植酸,并且提高鋅、鐵元素的生物利用率,利于人體吸收。此外,基本營養物質蛋白質、碳水化合物等代謝速率也加快,總蛋白質與氨基酸總量基本保持穩定,氨基酸組成發生了較大變化,使氨基酸組成更為合理,提高了大豆的營養品質。
[1] BRINCH-PEDERSEN H, MADSEN C K, HOLME I B, et al.Increased understanding of the cereal phytase complement for better mineral bio-availability and resource management[J]. Journal of Cereal Science, 2014, 59(3)∶ 373-381. DOI∶10.1016/j.jcs.2013.10.003.
[2] SANZ-PENELLA J M, LAPARRA J M, SANZ Y, et al. Bread supplemented with amaranth (Amaranthus cruentus)∶ effect of phytates on in vitro iron absorption[J]. Plant Foods for Human Nutrition, 2012,67(1)∶ 50-56. DOI∶10.1007/s11130-011-0269-6.
[3] AGUILERA Y, DíAZ M F, JIMéNEZ T, et al. Changes in nonnutritional factors and antioxidant activity during germination of nonconventional legumes[J]. Journal of Agricultural and Food Chemistry, 2013, 61(34)∶ 8120-8125. DOI∶10.1021/jf4022652.
[4] EL-ADAWY T A. Nutritional composition and antinutritional factors of chickpeas (Cicer arietinum L.) undergoing different cooking methods and germination[J]. Plant Foods for Human Nutrition, 2002,57(1)∶ 83-97. DOI∶10.1023/A∶1013189620528.
[5] PATHOMRUNGSIYOUNGGUL P, GRANDISON A S, LEWIS M J. Effect of calcium carbonate, calcium citrate, tricalcium phosphate,calcium gluconate and calcium lactate on some physicochemical properties of soymilk[J]. International Journal of Food Science &Technology, 2010, 45(11)∶ 2234-2240. DOI∶10.1111/j.1365-2621.2010.02399.x.
[6] WANG X K, YANG R Q, JIN X L, et al. Effect of supplemental Ca2+on yield and quality characteristics of soybean sprouts[J]. Scientia Horticulturae, 2016, 198∶ 352-362. DOI∶10.1016/j.scienta.2015.11.022.
[7] 江洪, 馬續紅, 吳謀成. 磷酸肌醇酯的生理活性及其研究進展[J]. 食品科技, 2007, 32(8)∶ 265-268. DOI∶10.3969/j.issn.1005-9989.2007.08.074.
[8] MANDAL N C, BISWAS B B. Metabolism of inositol phosphates 1.Phytase synthesis during germination in cotyledons of mung beans,Phaseolus aureus[J]. Plant Physiology, 1970, 45(1)∶ 4-7. DOI∶10.1104/pp.45.1.4.
[9] 靳曉琳, 王新坤, 楊潤強, 等. 高等植物體中植酸合成、代謝及其生理作用[J]. 植物生理學報, 2014, 50(6)∶ 711-716. DOI∶10.13592/j.cnki.ppj.2014.0048.
[10] REALE A, MANNINA L, TREMONTE P, et al. Phytate degradation by lactic acid bacteria and yeasts during the wholemeal dough fermentation∶ a31P NMR study[J]. Journal of Agricultural and Food Chemistry, 2004, 52(20)∶ 6300-6305. DOI∶10.1021/jf049551p.
[11] HUI Q R, YANG R Q, SHEN C, et al. Mechanism of calcium lactate facilitating phytic acid degradation in soybean during germination[J].Journal of Agricultural and Food Chemistry, 2016, 64(27)∶ 5564-5573.DOI∶10.1021/acs.jafc.6b01598.
[12] HUI Q R, WANG M, WANG P, et al. Gibberellic acid promoting phytic acid degradation in germinating soybean under calcium lactatetreatment[J]. Journal of the Science of Food and Agriculture, 2017,98(2)∶ 644-651. DOI∶10.1002/jsfa.8509.
[13] KARKLE E N L, BELEIA A. Effect of soaking and cooking on phytate concentration, minerals, and texture of food-type soybeans[J].Food Science and Technology (Campinas), 2010, 30(4)∶ 1056-1060.DOI∶10.1590/S0101-20612010000400034.
[14] 李合生. 植物生理生化實驗原理和技術[M]. 北京∶ 高等教育出版社,2000∶ 61-201.
[15] LISIEWSKA Z, S?UPSKI J, SKOCZE?-S?UPSKA R, et al.Content of amino acids and the quality of protein in brussels sprouts, both raw and prepared for consumption[J]. International Journal of Refrigeration, 2009, 32(2)∶ 272-278. DOI∶10.1016/j.ijrefrig.2008.05.011.
[16] BATISTI? O, KUDLA J. Analysis of calcium signaling pathways in plants[J]. Biochimica et Biophysica Acta, 2012, 1820(8)∶ 1283-1293.DOI∶10.1016/j.bbagen.2011.10.012.
[17] YIN Y Q, YANG R Q, HAN Y B, et al. Comparative proteomic and physiological analyses reveal the protective effect of exogenous calcium on the germinating soybean response to salt stress[J]. Journal of Proteomics, 2015, 113∶ 110-126. DOI∶10.1016/j.jprot.2014.09.023.
[18] METZLER-ZEBELI B U, DECKARDT K, SCHOLLENBERGER M,et al. Lactic acid and thermal treatments trigger the hydrolysis of myoinositol hexakisphosphate and modify the abundance of lower myoinositol phosphates in barley (Hordeum vulgare L.)[J]. PLoS ONE,2014, 9(6)∶ e101166. DOI∶10.1371/journal.pone.0101166.
[19] BERRIDGE M J. Inositol trisphosphate and calcium signalling[J].Nature, 1993, 361∶ 927-933.
[20] GIBBINS L, NORRIS F. Phytase and acid phosphatase in the dwarf bean, Phaseolus vulgaris[J]. Biochemical Journal, 1963, 86(1)∶ 67-71.
[21] 羅羽洧, 解衛華, 馬娟, 等. 不同發芽條件對蠶豆植酸酶活性的影響[J]. 中國糧油學報, 2012, 27(10)∶ 32-36. DOI∶10.3969/j.issn.1003-0174.2012.10.007.
[22] WEAVER C M, KANNAN S. Food phytates[M]. Boca Raton∶ CRC Press, 2002∶ 211-223.
[23] LESTIENNE I, MOUQUET-RIVIER C, ICARD-VERNIèRE C, et al. The effects of soaking of whole, dehulled and ground millet and soybean seeds on phytate degradation and Phy/Fe and Phy/Zn molar ratios[J]. International Journal of Food Science & Technology, 2005,40(4)∶ 391-399. DOI∶10.1111/j.1365-2621.2004.00941.x.
[24] ADEYEYE E, AROGUNDADE L, AKINTAYO E, et al. Calcium, zinc and phytate interrelationships in some foods of major consumption in Nigeria[J]. Food Chemistry, 2000, 71(4)∶ 435-441. DOI∶10.1016/S0308-8146(00)00159-X.
[25] MEDIC J, ATKINSON C, HURBURGH J R C R. Current knowledge in soybean composition[J]. Journal of the American Oil Chemists’Society, 2014, 91(3)∶ 363-384. DOI∶10.1007/s11746-013-2407-9.
[26] SALDIVAR X, WANG Y J, CHEN P, et al. Changes in chemical composition during soybean seed development[J]. Food Chemistry,2011, 124(4)∶ 1369-1375. DOI∶10.1016/j.foodchem.2010.07.091.
[27] 王新坤, 仲磊, 楊潤強, 等. 植物籽粒中植酸及其降解方法與產物研究進展[J]. 食品科學, 2014, 35(3)∶ 301-306. DOI∶10.7506/spkx1002-6630-201403059.
[28] 韓立德, 蓋鈞鎰, 張文明. 大豆營養成分研究現狀[J]. 種子, 2003(5)∶58-60. DOI∶10.3969/j.issn.1001-4705.2003.05.023.
[29] 尹永祺. NaCl及其聯合Ca2+處理下發芽大豆生理變化與GABA富集調控機理[D]. 南京∶ 南京農業大學, 2014.
[30] BALASUNDRAM N, SUNDRAM K, SAMMAN S. Phenolic compounds in plants and agri-industrial by-products∶ antioxidant activity, occurrence, and potential uses[J]. Food Chemistry, 2006,99(1)∶ 191-203. DOI∶10.1016/j.foodchem.2005.07.042.
猜你喜歡
農業科技通訊(2023年1期)2023-02-12 07:09:18
今日農業(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年7期)2022-08-31 01:29:28
中國化肥信息(2022年5期)2022-08-30 01:58:26
今日農業(2021年20期)2021-11-26 01:23:56
今日農業(2021年14期)2021-10-14 08:35:34
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
農產品市場周刊(2017年4期)2017-03-03 19:40:05
兒童故事畫報·智力大王(2015年10期)2016-01-27 01:01:35
讀寫算(中)(2015年10期)2015-11-07 07:24:12
