西藏色季拉山川滇高山櫟群落生態位特征
2018-07-02 11:20:48尼瑪曲珍張璽譚坤魯闖王國嚴徐阿生羅建
生態科學 2018年3期
尼瑪曲珍, 張璽, 譚坤, 魯闖, 王國嚴,*, 徐阿生, 羅建
1. 西藏農牧學院資源與環境學院, 西藏 林芝 86000
2. 西藏農牧學院高原生態研究所, 西藏 林芝 86000
1 前言
物種共存和生物多樣性維持一直是生態學研究的中心論題。探索群落構建(community assembly)背后的過程機制對于預測環境變化背景下的群落動態和開展區域物種多樣性保護至關重要[1]。一個世紀以來, 生態學家提出了眾多解釋物種共存的理論和假說[2], 包括種庫理論、更新生態位理論、資源比率/異質性假說、競爭共存理論、生態漂變假說、中性理論等[3-6], 其中生態位理論和中性理論先后得到了較為廣泛的驗證和關注[7]。大量研究證明, 在溫帶森林群落生態位理論是解釋物種共存的重要生態學機理[8-9]。生態學家基于生態位寬度值和生態位重疊值[10-11]入手,揭示群落構建和物種共存機制。生態位寬度不但反映物種對環境資源利用狀況的尺度, 也表征了物種的生態適應性和分布幅度。物種的生態位寬度指數越大, 說明該物種在群落中的地位越高, 分布越廣,對資源利用充分且對所在環境有較強的適應力[10]。基于生態位理論的群落構建理論可以被認為是物種的篩選過程, 環境條件和生物間的相互作用可以被看作是多個嵌套的篩子(nested sieves); 群落構建將區域物種庫中的物種經過這些嵌套篩子的過濾, 只有那些具有特定性狀并符合各環境篩的特定物種才能進入小局域群落[7]。生態位重疊是指一定資源序列上, 兩個物種利用同級資源而相互重疊的情況。生態位重疊值較大預示著這兩個種可能具有相似的生態學和生物學特性, 或者對資源的需求有一定的互補性。同等資源序列上, 物種間較高的生態位重疊有利于多物種的共存。
藏東南地區是我國面積最大的原始林區之一, 由于地理和交通條件限制, 森林群落受人為干擾相對較少, 是開展群落生態學理論研究的天然實驗室[13]。高山櫟類硬葉常綠闊葉林是全世界分布最廣, 也是青藏高原的主要常綠闊葉林。其中, 川滇高山櫟普遍存在于西藏東南部, 對該區的水土保持和涵養水源方面具有重要的作用[13-14]。近年來, 川滇高山櫟的研究主要集中在種群結構與動態[15]、種群空間格局[16]、種群統計特征[17-18]、葉片特性[19]、遺傳多樣性的變化[20]、營養元素供應[21]等方面, 關于藏東南川滇高山櫟群落生態位的研究尚未見報道。本文通過研究色季拉山川滇高山櫟種群植物生態位特征, 旨在揭示該區川滇高山櫟種群在群落中的功能地位、生態適應性及各種群對資源的利用狀況, 為該區森林群落的經營管理和保護提供理論依據。
2 研究地區與研究方法
2.1 研究地概況
研究區位于西藏色季拉山, 地理坐標為 29°33′—30°00′ N, 94°19′—94°53′ E。該區屬于較典型的亞高山溫帶半濕潤氣候區, 冬溫夏涼、干濕季分明。是西藏東南部雅魯藏布江大峽谷西北側, 系念青唐古拉山向南延伸的余脈, 與喜馬拉雅山向東發展的山系相連, 為藏東南林區的腹心地帶, 也是自然環境極為特殊多樣的地區。研究的范圍在海拔 3600—4300 m。年平均氣溫-0.73 ℃, 最高月(7月)平均氣溫9.23 ℃, 最低月(1月)平均氣溫-13.98 ℃, 極端最低氣溫-31.6 ℃, 極端最高氣溫24.0 ℃, 年均日照時數1150.6 h, 日照百分率26.1%, 日照時數最高月為12月(151.7 h), 日照百分率為 40%, 年均相對濕度78.83%, 年均降水量1134.1 mm, 蒸發量 544.0 mm,占年均降水量的48.0%。6—9月為雨季, 占全年降水的75%—82%, 其中8月降雨最多, 平均為294.2 mm,占全年降水的 30%, 土壤主要以山地棕壤和酸性棕壤為主, PH值為4—6, 土層較厚, 平均厚度60 cm,腐殖質化程度不明顯[13]。
該區域川滇高山櫟林分結構相對單一, 多以純林分布, 有時與高山松(Pinus densataMast.)混交成為次林層優勢種。研究林分喬木層主要有川滇高山櫟、高山松、糙皮樺(Betula utilisD. Don), 偶爾出現華山松(P. armandiFranch.)、沙棘(Hippophae rhamnoidesL.)等, 個別林下有急尖長苞冷杉(Abies georgeivar.smithiiCheng et L.K. Fu) 或林芝云杉(Picea likiangensisvar.linzhiensisCheng et L.K.Fu)幼苗; 灌木層主要有雅致山馬蝗(Desmodium elegansDC.)、粉背野丁香(Leptodermis potaniniivar.Glauca(Diels) H.Winkl.)、西南野丁香(L.purdomiiHutch.)等; 草本層有秦嶺槲蕨(Drynaria sinicaDiels)、高山唐松草(Thalictrum alpinumL.)、尼泊爾大丁草(Leibnitzia nepalensis(Kunze)Kitam.)等; 藤本有防己葉菝葜(Smilax menispermoideaA.DC.)和繡球藤(Clematis montanaBuch.-Ham.et DC.)等。
2.2 研究方法
調查從雅魯藏布江以北川滇高山櫟在我國的分布西限-工布江達縣金達鎮開始, 向東延伸至林芝縣東久一帶, 覆蓋川滇高山櫟在我國青藏高原的主要分布區[12], 海拔范圍3050—3800 m, 氣候自東向西逐漸從高原濕潤氣候向半濕潤氣候過渡[16]。選擇代表區域種群特征的典型樣地展開調查, 共設置20×20 m2樣地13個, 共計調查面積5200 m2。詳細樣地分布及氣候特征參見文獻[22]。
記錄樣地的海拔高度、經緯度、坡度、坡向、各層次高度、郁閉度和死地被層及土壤等指標。對喬木層進行每木檢尺, 包括種、胸徑、樹高、冠幅、生長狀況以及林內更新狀況等。在每個樣地內按五點取樣法(樣地四角和中心)設置5個2×2 m2的灌木小樣方, 并在每個灌木樣方選取1×1 m2草本小樣方,分別記錄5×5 m2面積內灌木層和1×l m2面積內草本層的種類、株(叢)數、高度和蓋度等。
2.3 數據分析
以群落中物種的重要值代表物種在不同資源位當中的優勢程度。基于不同調查樣地的物種重要值矩陣, 以樣地為資源位測算群落物種生態位寬度和生態位重疊。
1) 重要值數據
13個樣地中共記錄到49個種植物, 數據分析采用重要值作為綜合指標來反映種的特征[22], 計算公式如下:
喬木層重要值=(相對多度+相對優勢度+相對頻度)/3
灌木層重要值=(相對多度+相對優勢度+相對蓋度+相對頻度)/4
草本層重要值=(相對多度+相對優勢度+相對蓋度+相對頻度)/4
2)生態位寬度
除去群落中重要值<1的種群, 建立49×13的數據矩陣進入生態位運算。
Levins指數[23]
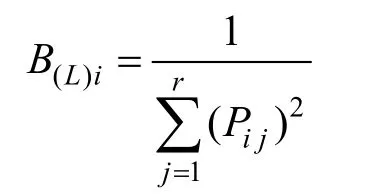
式中,Bi為種群i的生態位寬度為種群i利用資源狀態j的數量(以種群i在第j個樣方的重要值表示),Ni為種群i的總數量,r為樣方數。
信息指數(Shannon-Wiener)指數
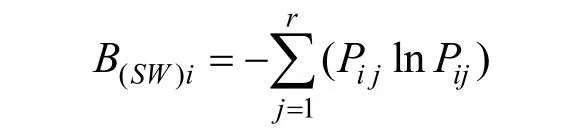
以上的兩個指數iB值越大, 說明生態位越寬。當種i的所有個體以相等的數目利用每一資源狀態時,iB最大, 即該種具有最寬的生態位; 當種i的所有個體都集中在某一個資源狀態下時,iB最小, 該種具有最窄的生態位[24-25]。
3)生態位重疊
Horn(1966)重疊指數
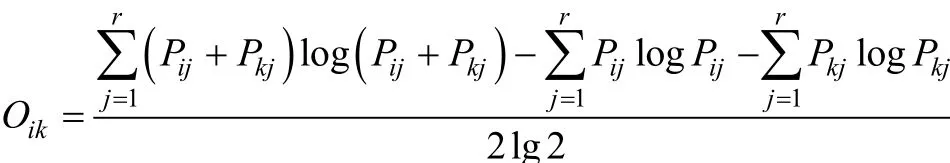
式中,Oik為種類i對種類k的生態位重疊指數,Pij、Pkj為種類i或種類k所利用的整個資源中第j種資源所占比例,r為資源狀態總數[26]。將群落喬、灌、草層次重要值數據分別進行標準化和中心化(均值為0, 標準差為1), 再根據以上公式計算層次間生態位重疊指數。
3 結果
3.1 重要值與生態位寬度
在喬木層中, 川滇高山櫟的重要值最大, 為82.425,生態位寬度最大(表1)。在灌木層中, 雅致山螞蝗的重要值為16.554, 具有優勢。小葉忍冬在2、3、9、10號樣地中與川滇高山櫟大量混生, 蓋度較大, 適應能力強, 生態位寬度大。在草本層中秦嶺槲蕨的
重要值較大 8.252。秦嶺槲蕨、雙參與尼泊爾蓼在13個樣地中出現頻數都為 7, 屬于川滇高山櫟群落中的優勢種, 分布較普遍, 生態位寬度也較大; 相反, 高山唐松草在群落中數量相對少, 13個樣地中僅有一次出現, 重要值也較小, 分布范圍較窄, 生態位寬度很小。
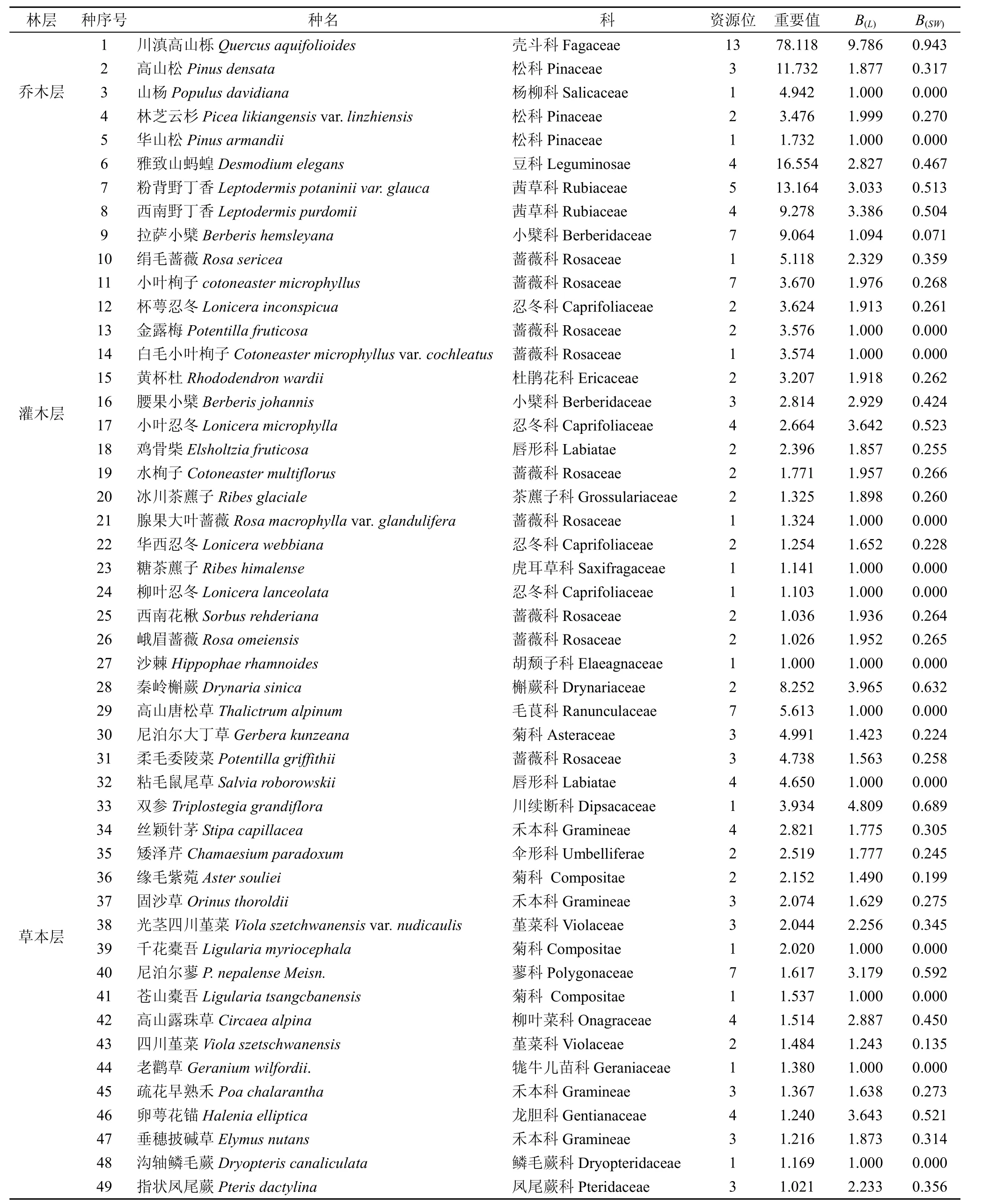
表1 色季拉山川滇高山櫟群落植物重要值及生態位寬度Tab. 1 Importance values and niche breadths of main plant species in the Quercus aquifolioides community in Sejila Mountains
在川滇高山櫟群落中生態位寬度()LB值排前10位的主要種群順序為: 川滇高山櫟>雙參>秦嶺槲蕨>卵萼花錨>小葉忍冬>西南野丁香>尼泊爾蓼>粉背野丁香>腰果小檗;()SWB值的順序為: 川滇高山櫟>雙參>秦嶺槲蕨>尼泊爾蓼>小葉忍冬>卵萼花錨>粉背野丁香>西南野丁香>雅致山馬蝗>高山露珠草。兩指數計測結果排列順序雖有差異, 但總體一致, 川滇高山櫟、雙參、秦嶺槲蕨、尼泊爾蓼、小葉忍冬、卵萼花錨、粉背野丁香、西南野丁香的生態位寬度值均較高, 在群落中占優勢地位。
3.2 生態位重疊
3.2.1 群落各林層內的生態位重疊
從表 2可知, 在川滇高山櫟群落的喬木樹種當中, 林芝云杉與華山松的生態位重疊較高 0.612, 而川滇高山櫟與高山松、山揚、林芝云杉、華山松的生態位重疊普遍較低, 多分布在0.31—0.35。群落灌木層中, 冰川茶藨子、腺果大葉薔薇、華西忍冬、糖茶藨子、柳葉忍冬、西南花楸、峨眉薔薇、沙棘9個種之間的生態位重疊值較大(見表 3, 0.855—0.998)。表4顯示, 群落草本層中四川堇菜、老鸛草、卵萼花錨、垂穗披堿草、溝軸鱗毛蕨、指狀鳳尾蕨的生態位重疊值較大(0.817—0.998)。
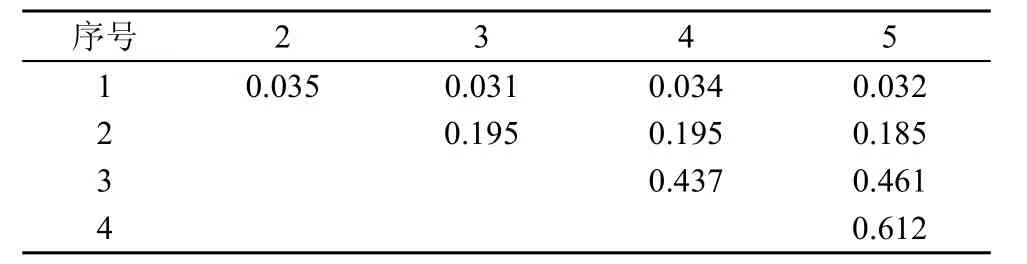
表2 喬木樹種生態位重疊值Tab. 2 Niche overlaps between tree species
由圖1A可知, 該群落喬木植物生態位重疊值與灌木植物重疊值主要集中在 0.5—0.9, 分別占100%和87%。說明該群落喬木植物內部與灌木植物內部生態位普遍重疊, 并且重疊程度較高。該群落草本植物生態位重疊值主要集中在 0.8—1, 所占比例達到 65%。各林層內部生態位重疊程度順序為:草本層>灌木層>喬木層。
3.2.2 群落各林層之間的生態位重疊
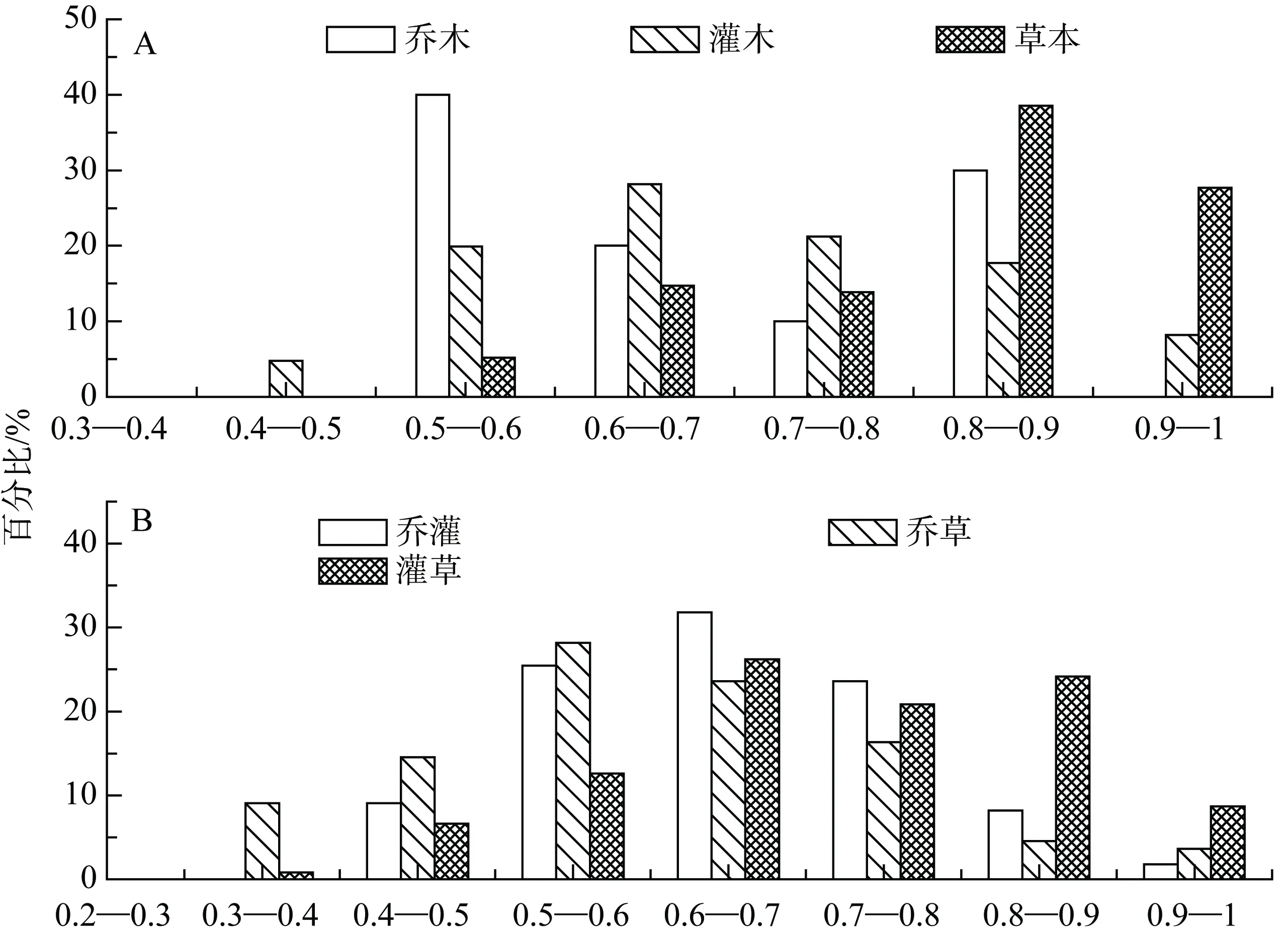
圖1 林層內(A)與層間(B)物種生態位重疊分配格局Fig. 1 Distribution patterns of niche overlaps of plant populations within (A) and between (B) forestry layers
由圖1B可知, 該群落喬灌植物之間與喬草植物之間生態位重疊值主要集中在 0.5—0.8, 其中喬灌植物之間生態位重疊值為 0.6—0.7所占比例最高,達32%。喬草植物之間生態位重疊值為0.5—0.6所占比例最高28%。該群落灌草植物之間生態位重疊值主要集中在0.6—0.9, 所占比例高達70%。各林層之間生態位重疊程度順序為: 灌草>喬灌>喬草。
4 討論
生態位寬度是度量種群對環境資源利用能力的尺度[27]、是種群動態的一個間接測度和種群在群落中的地位和作用的數量表達, 能夠較好地解釋群落演替過程中種群的環境適應性和資源利用能力[28]。該群落中生態位寬度較大的物種有川滇高山櫟、雙參、秦嶺槲蕨、尼泊爾蓼、小葉忍冬、卵萼花錨、粉背野丁香、西南野丁香等。灌木層中生態位寬度最大的是小葉忍冬; 草本層生態位寬度最大的為秦嶺槲蕨, 說明這些植物在川滇高山櫟群落中生態幅較寬, 具有較強的環境適應能力和資源利用能力,在藏東南不同的資源位條件下都能很好地生長, 在川滇高山櫟群落構建中起著重要的作用。川滇高山櫟的重要值和生態位寬度都較大, 形成單優勢種群落, 在群落中占有主導地位。即使川滇高山櫟與高山松等混生, 居于林下, 數量亦較多, 在立地中具有一定優勢, 這與其它建群種植物具有一定的相似性, 如珙桐[29]、矮牡丹[30]、云貴鵝耳櫟[31]、秦嶺冷杉[32]。川滇高山櫟作為藏東南優勢先鋒樹種, 在干擾和火燒跡地廣泛分布, 在演替中后期林下出現云杉或高山松的幼苗或幼樹, 然而由于后者生長相對緩慢且群落演替過程復雜, 在相當長一個時期內依舊由川滇高山櫟占優勢。因此, 作為藏東南地區重要的水土保持森林群落之一, 川滇高山櫟在其群落中的地位及在發揮群落的水土保持與荒漠化防治作用中意義突出。
生態位重疊表明物種間在資源利用上的交叉和重疊程度, 反映群落物種共存方式, 對生物多樣性的維持具有重要意義。藏東南川滇高山櫟群落喬木層中, 川滇高山櫟與其他四種喬木的重疊指數都不高(<0.035), 也證明了川滇高山櫟在群落喬木層的絕對優勢地位。林芝云杉和華山松的生態位重疊較高, 兩者分別為耐陰和陽性樹種在藏東南地區的陽坡地帶有時形成混交林, 對光照的不同生態位有助于在同一地段共存且減小競爭。
該群落各個林層內部重疊程度順序為: 草本層>灌木層>喬木層。草本層內部物種生態位普遍重疊,且重疊程度很高。各層次的物種數與生態位重疊程度呈負相關關系, 與大量溫帶森林群落的特征相似[8-9], 證明了生態位重疊對多物種共存及生物多樣性維持的重要意義。高山地區植物分布多受降水條件控制, 結合不同樣地的環境狀況與之前對川滇高山櫟群落的物種多樣性分布研究結果[22], 色季拉山小氣候特征隨地形變化, 草本層對降水的反應比其他植物更為敏感, 數量、高度和冠幅受降水的影響變化較大, 在群落中分布不均勻。隨著降水量增大, 草本層在群落中的重要值、生態位寬度及生態位重疊值都增大, 反之隨降水量減少這些指標也減少, 說明降水是影響色季拉山川滇高山櫟群落草本植物分布的關鍵資源, 對群落和更新演替起著控制作用。與我國暖溫帶落葉闊葉林群落相比[10,24], 色季拉山森林群落物種豐富度更低, 而喬木層物種間生態位重疊值較低, 灌木和草本層物種生態位重疊度則更高, 反映出藏東南地區受高寒氣候影響, 物種數量整體較少, 森林多為單優勢種類型, 喬木樹種生態位分化明顯。同時高山地區強烈的環境梯度和環境篩選作用導致林下廣布種相對較少, 大量的灌木和草本層種間以共同生生境偏好而同時出現于同一資源位的概率較高, 出現較高的生態位重疊。
群落各個林層之間重疊程度順序為: 灌草>喬灌>喬草。灌草層植物之間生態普遍重疊, 且重疊程度很高。相鄰兩個生活型之間重疊度高于距離較遠的生活型之間的重疊程度, 說明空間分層作為群落物種生態位分化的最顯著特征, 層間物種關系以資源利用方式互補甚至互利的形式存在, 空間分層大大促進了群落物種的共存。
生態位作為解釋物種多樣性共存的重要假說之一, 雖然近年來受到群落中性理論的沖擊和爭論,但大量研究證明, 在溫帶森林群落隨機因素貢獻較熱帶植物群落低很多, 生態位理論依舊是解釋物種共存的重要生態學機理[1]。藏東南林區作為我國重要的原始林區之一, 由于經濟、交通等原因, 其生物多樣性研究還剛剛起步。本研究分析了藏東南川滇高山櫟群落主要物種的生態位特征, 初步揭示了川滇高山櫟群落物種的生態位特征, 發現川滇高山櫟在其群落中地位高, 分布廣, 對資源利用充分且對所在環境有較強的適應力; 高山地區強烈的環境過濾作用導致的生態位分化和對生境需求相似的物種的生態位重疊在藏東南川滇高山櫟群落物種共存中發揮重要作用。川滇高山櫟群落是藏東南地區的重要水土保持植物群落, 認識其群落物種共存規律對該地區的生態維持和受損生態系統恢復具有重要意義。
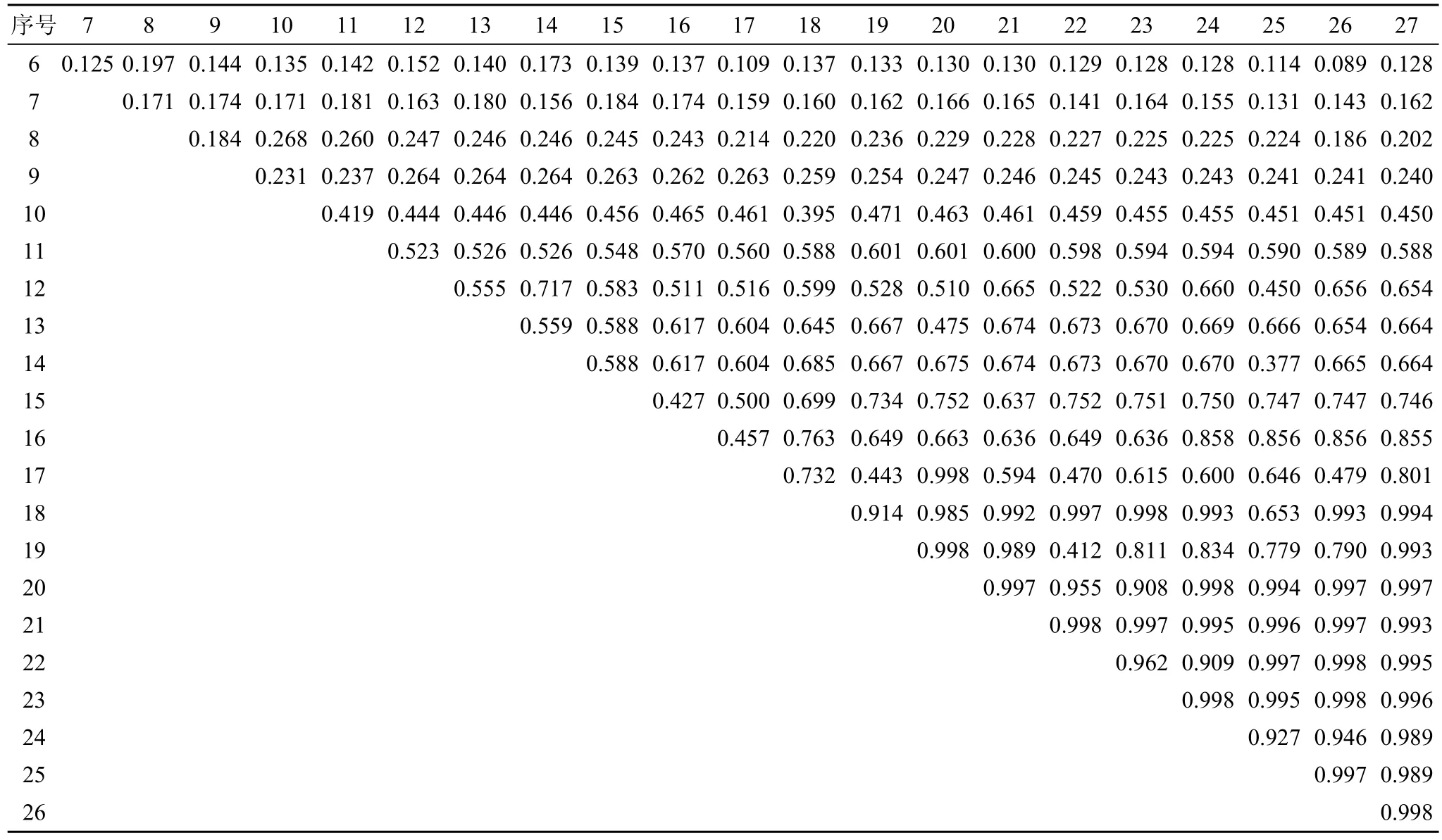
表3 主要灌木樹種生態位重疊值Tab. 3 Niche overlaps of dominant shrub species
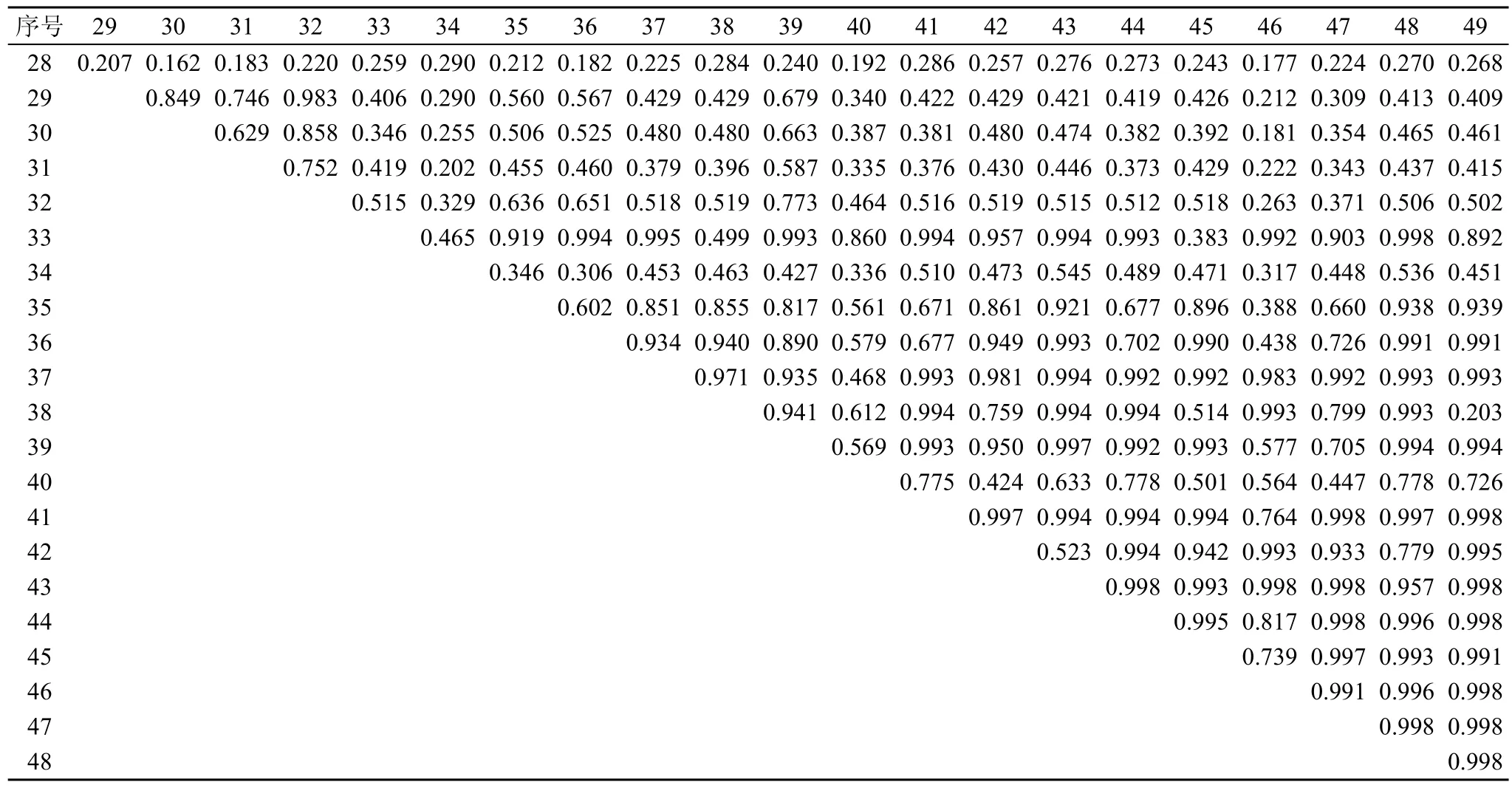
表4 主要草本種生態位重疊值Tab. 4 Niche overlaps of dominant herb species
[1] G?TZENBERGER L, DE BELLO F, BR?THEN K A, et al. Ecological assembly rules in plant communitiesapproaches, patterns and prospects[J]. Biological Reviews of the Cambridge Philosophical Society, 2012, 87(1):111-127.
[2] WRIGHT J S. Plant diversity in tropical forests: a review of mechanisms of species coexistence[J]. Oecologia, 2002,130: 1-14.
[3] 侯繼華, 馬克平. 植物群落物種共存機制的研究進展[J].植物生態學報, 2002, 26: 1-8.
[4] HUBBELL S P. The Unified Neutral Theory of Biodiversity and Biogeography[M].New Jersey: Princeton University Press, 2001: 12-16.
[5] TAYLOR D R, AARSSEN L W, LOEHLE C. On the relationship between r/K selection and environmental carrying capacity: a new habitat templet for plant life history strategies[J]. Oikos, 1990, 58: 239-250.
[6] CONNELL J H. On the role of natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees[J]. Dynamics of Populations, 1971,298: 312-317.
[7] 牛克昌, 劉懌寧, 沈澤昊, 等. 群落構建的中性理論和生態位理論[J]. 生物多樣性, 2009, 17(6): 579-593.
[8] 陳云, 郭凌, 姚成亮, 等. 暖溫帶-北亞熱帶過渡區落葉闊葉林的群落特征[J]. 生態學報, 2017, 37(17): 1-10.
[9] 龔容, 高瓊, 王亞林. 圍封對溫帶半干旱典型草原群落種間關聯的影響[J]. 植物生態學報, 2016, 40(6): 554-563.
[10] 白曉航, 張金屯. 小五臺山森林群落優勢種的生態位分析[J]. 應用生態學報, 2017, 28(12): 1-16.
[11] 吳會峰, 宋麗娟, 杜峰, 等. 黃土丘陵區草地主要種群生態位及其環境解釋[J]. 應用生態學報, 2017, 28(11):1-14.
[12] 王國嚴, 徐阿生. 川滇高山櫟研究綜述[J]. 四川林業科技, 2008, 29(2): 23-29.
[13] 徐鳳翔. 西藏高原森林生態研究[M]. 沈陽: 遼寧大學出版社, 1995: 1-41.
[14] 石小亮, 張穎, 單永娟, 等. 云南省高原典型森林植被涵養水源功能研究[J]. 長江流域資源與環境, 2015, 24(8):1366-1372.
[15] 劉興良. 川西巴郎山地川滇高山櫟林群落生態學的研究[D].北京: 北京林業大學, 2006: 38-45.
[16] 沈志強, 華敏, 丹曲, 等. 藏東南川滇高山櫟種群不同生長階段的空間格局與關聯性[J]. 應用生態學報, 2016,27(2): 387-394.
[17] 劉興良, 岳永杰, 鄭紹偉, 等. 川滇高山櫟種群統計特征的海拔梯度變化[J]. 四川林業科技, 2005, 26(4): 9-15.
[18] 楊朗生, 劉興良, 劉世榮, 等. 臥龍巴郎山川滇高山櫟群落植物生活型海拔梯度特征[J]. 生態學報, 2017, 37(21):1-11.
[19] 劉興良, 何飛, 樊華, 等. 臥龍巴郎山川滇高山櫟群落植物葉特性海拔梯度特征[J]. 生態學報, 2013, 33(22):7148-7156.
[20] 程小毛, 李響, 姜永雷, 等. 基于AFLP的滇西北玉龍雪山不同海拔川滇高山櫟遺傳多樣性分析[J]. 華中農業大學學報, 2017, 36(01): 22-27.
[21] 朱萬澤, 王三根, 郝云慶. 川滇高山櫟灌叢萌生過程中的營養元素供應動態[J]. 植物生態學報, 2010, 34(10):1185-1195.
[22] 王國嚴, 羅建, 徐阿生, 等. 藏東南川滇高山櫟群落物種多樣性格局[J]. 林業科學研究, 2012, 25(06): 703-711.
[23] LEVINS R. Evolution in Changing Environments: Some Theoretical Exploration[D]. Princeton: Princeton University Press, 1968: 25-33.
[24] 林勇, 艾訓儒, 姚蘭, 等. 木林子自然保護區不同群落類型主要優勢種群的生態位研究[J]. 自然資源學報, 2017,32(02): 223-234.
[25] 周亞琦, 官鳳英, 范少輝, 等. 天寶巖竹闊混交林毛竹及其伴生樹種生態位的研究[J]. 北京林業大學學報, 2017,39(07): 46-53.
[26] 張金屯. 數量生態學[M]. 北京: 科學出版社, 2004:135-143.
[27] 王霞, 鐵軍, 李燕芬, 等. 山西陵川南方紅豆杉群落優勢種種間關聯性分析[J]. 生態學雜志, 2016, 35(10):2575-2582.
[28] 李亞, 胡小柯, 魏懷東, 等. 騰格里沙漠南緣天然群落主要種群空間分布格局研究[J]. 西北林學院學報, 2017,32(2): 67-72.
[29] 劉海洋, 金曉玲, 沈守云, 等. 湖南珍稀瀕危植物--珙桐種群數量動態[J]. 生態學報, 2012, 32(24): 7738-7746.
[30] 張峰. 瀕危植物矮牡丹致瀕原因分析[J]. 生態學報,2003, (7): 1436-1441.
[31] 馮祥麟, 胡剛, 劉正華, 等. 貴陽山地森林喬木層結構及主要種群生態位特征初步研究[J]. 貴州科學, 2011, 29(1):40-46.
[32] 韋博良, 陳云, 許寧, 等. 小秦嶺國家級自然保護區維管植物群落結構與組成[J]. 河南農業大學學報, 2015, 49(3): 335-342.
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
吉林廣播電視大學學報(2021年4期)2022-01-14 02:35:48
保健醫苑(2021年7期)2021-08-13 08:48:02
作文成功之路·小學版(2020年5期)2020-06-11 12:48:26
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
小天使·一年級語數英綜合(2018年11期)2018-11-23 09:47:26
汽車觀察(2018年10期)2018-11-06 07:05:26
資源再生(2017年3期)2017-06-01 12:20:59
