Na+在鴨茅適應抗氧化防御和滲透脅迫中的生理作用
2018-08-08 08:08:10梁小玉胡遠彬
江蘇農業科學 2018年14期
關鍵詞:植物
季 楊,梁小玉,易 軍,胡遠彬
(四川省畜牧科學研究院,四川成都 610066)
干旱作為一種水量相對虧缺的自然現象,是限制植物生長發育和產量提高的主要逆境因子之一[1-2]。干旱脅迫下,抗氧化保護系統是植物抵御干旱脅迫最重要的機制之一,大量研究證明耐旱性強的鴨茅(DactylisglomerataL.)、白三葉(TrifoliumrepensL.)、黑麥草(LoliumperenneL.)等植物具有更強的抗氧化保護機制[3-5],從而有效提高植物的耐旱性。同時,滲透調節也是植物抵御逆境脅迫,維持正常生命活動不可或缺的重要生理機制[6]。Na+作為重要的無機滲透調節物質,高濃度的Na+能夠對植物造成鹽害,嚴重時能夠使植物發生質壁分離而死亡[7]。越來越多的研究表明,低濃度的Na+對旱生植物抵御干旱脅迫發揮著積極的作用[8-9]。
目前,有關Na+提高植物抗旱性的研究主要集中在旱生植物中,而在中生植物中報道較少。鴨茅是溫帶叢生冷季禾本科牧草之一,因具有產量豐富、營養價值高、耐牧性強等優點而被廣泛推廣利用。然而,鴨茅喜溫暖和濕潤氣候,耐旱性相對較差,干旱成為限制其潛力發揮的重要逆境因子。因此,本研究以敏感型鴨茅為材料,探討了無機滲透調節物質Na+參與干旱脅迫下鴨茅耐旱機制,為提高鴨茅的耐旱性奠定理論基礎。
1 材料與方法
1.1 試驗材料
供試材料為鴨茅敏感型材料“01998”。精選均勻飽滿種子,經6%次氯酸鈉溶液消毒5 min后,用蒸餾水沖洗3~4次,播種在石英砂上并于24 ℃光照培養箱中進行發芽,待發芽7 d后用Hoagland全營養液繼續培養,溫度設置為白天 22 ℃,夜晚15 ℃,相對含水量80%,光照300 μmol/(m2·s),光—暗周期14 h—10 h。
1.2 試驗設計
當幼苗長至3周齡時,幼苗分為4組:(1)CK(對照,Hoagland全營養液正常培養生長);(2)NaCl+CK(在Hoagland全營養液中加入50 mmol/L的NaCl處理5 d后,更換為正常Hoagland全營養液);(3)PEG(使用含有15% PEG-6000的Hoagland全營養液進行水分脅迫處理);(4)NaCl+PEG(在Hoagland全營養液中加入50 mmol/L的NaCl處理5 d后,更換為15% PEG-6000的Hoagland全營養液進行水分脅迫);每2 d更換1次處理液,9 d后取樣,分別測定各項指標,每個處理取4個重復。
1.3 測定指標與方法
根系活力(root vitality)采用四氮唑法TTC還原法測定[10];相對含水量(relative water content,RWC)采用烘干法測定[11],相對含水量(%)=(鮮質量-干質量)/(飽和鮮質量-干質量)×100%;丙二醛(malondialdehyde,MDA)含量采用硫代巴比妥酸法測定[12];超氧化物歧化酶(superoxide dismutase,SOD) 活性采用核黃素-NBT 法測定[13];過氧化氫酶(catalase,CAT)活性采用紫外吸收法測定;過氧化物酶(guaiacol peroxidase,POD) 活性測定用愈創木酚顯色法[14];電解質滲透率(electrolyte leakage,EL)采用電導儀測定[15];游離脯氨酸(free proline)含量采用茚三酮比色法測定[16];可溶性糖(soluble sugar)含量采用蒽酮乙酸乙酯比色法測定[16];超氧陰離子(superoxide anion)產生速率和過氧化氫(H2O2)含量按照郝建軍等[17]和Uchida等[18]的方法進行測定。
1.4 統計分析
圖表用Excel軟件進行繪制,利用SAS 8.1軟件進行方差分析和顯著性檢驗。
2 結果與分析
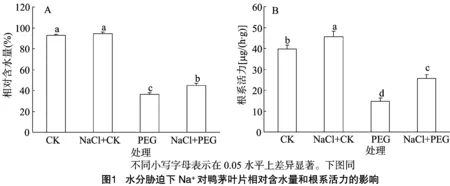
2.1 水分脅迫下Na+對鴨茅葉片相對含水量和根系活力的影響
植物葉片相對含水量反映植物在逆境脅迫下的整體水分虧缺狀況。正常水分條件下添加外源NaCl對鴨茅葉片相對含水量較CK沒有顯著影響。15% PEG水分脅迫9 d后,鴨茅葉片相對含水量顯著下降,但在相同脅迫強度和脅迫時間下,PEG+NaCl葉片相對含水量較PEG處理顯著提高(圖1-A)。如圖1-B所示,4個處理的根系活力存在顯著差異,外源NaCl處理不僅顯著提高了水分脅迫下鴨茅根系活力,也提高了正常水分下鴨茅的根系活力。
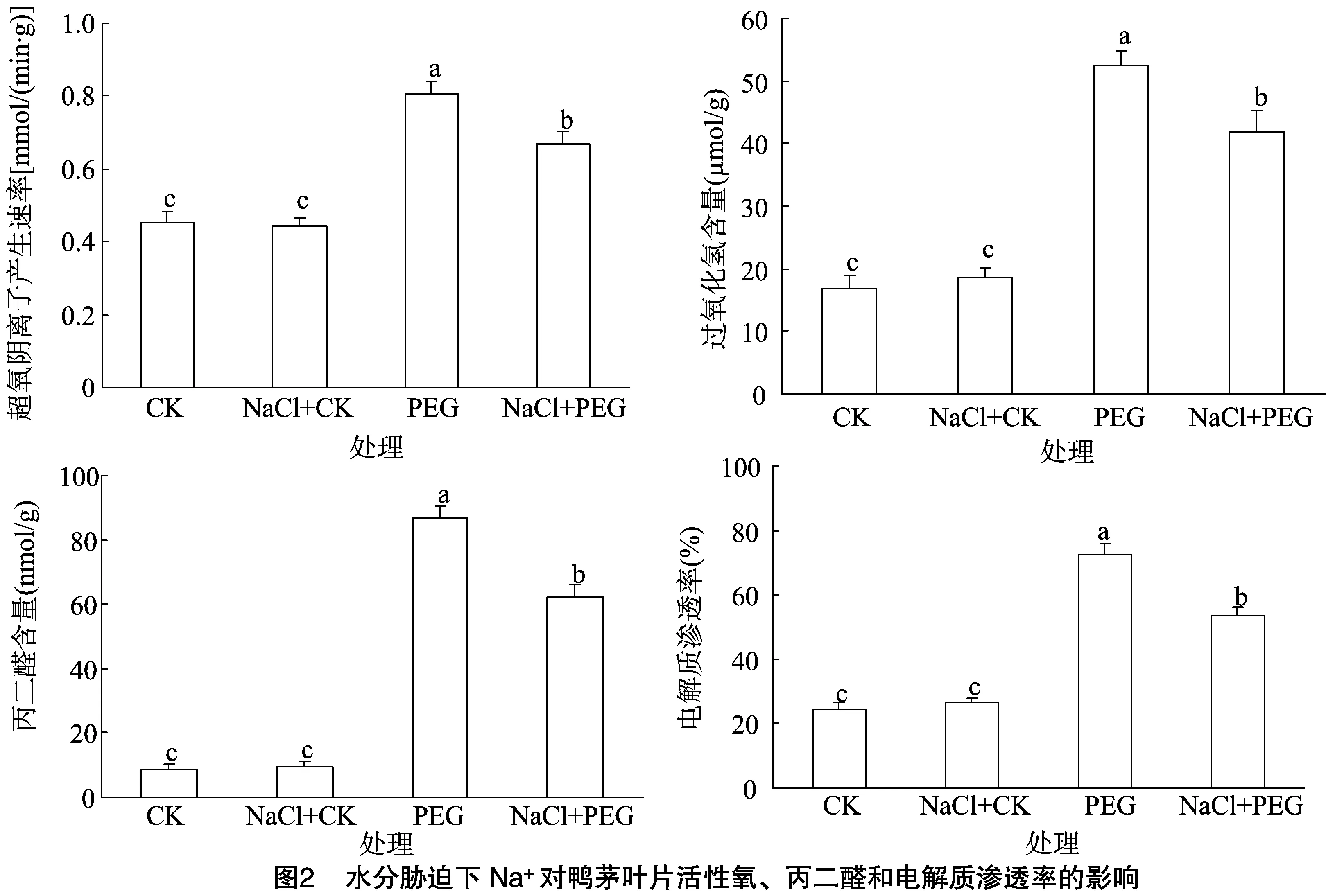
2.2 水分脅迫下Na+對鴨茅葉片活性氧、丙二醛含量和電解質滲透率的影響
從圖2可以看出,正常水分條件下,外源NaCl對鴨茅葉片內活性氧成分、MDA含量和電解質滲透率幾乎沒有任何影響,說明低濃度NaCl對鴨茅生長沒有造成脅迫傷害。鴨茅葉片內活性氧成分、MDA含量和電解質滲透率在水分脅迫條件下迅速增加,并顯著高于正常水分處理。其中,PEG處理和PEG+NaCl處理的超氧陰離子產生速率分別是對照CK的1.8、1.5 倍,H2O2含量分別是CK的3.2、2.5 倍,MDA含量分別是CK的10.0、7.2倍,電解質滲透率分別是CK的3.0、2.2倍。結果說明,外源NaCl顯著降低了干旱脅迫下葉片內超氧陰離子產生速率、葉片內H2O2和MDA積累以及電解質外滲,維持了脅迫下鴨茅葉片細胞膜系統的穩定性,有效緩解了水分脅迫造成的氧脅迫傷害。
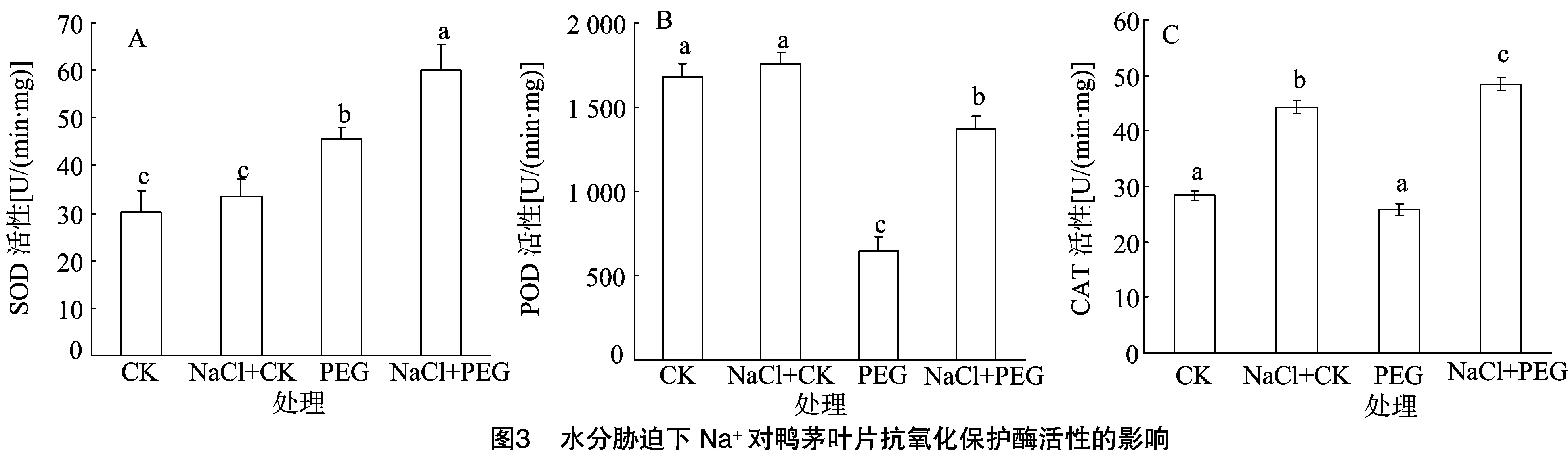
2.3 水分脅迫下Na+對鴨茅葉片抗氧化保護酶活性的影響
SOD、CAT和POD是植物體內清除活性氧的重要酶類。結果表明,水分脅迫條件下,鴨茅葉片SOD活性較對照顯著增加。當添加外源NaCl處理后,PEG+NaCl處理的SOD活性比PEG處理的活性增加24%,差異達顯著水平(圖3-A)。水分脅迫使鴨茅葉片POD活性顯著降低,在添加外源NaCl處理后下降幅度顯著減小,表明外源NaCl的添加有效緩解了水分脅迫對POD活性的抑制(圖3-B)。從圖3-C可以看出,水分脅迫處理CAT活性較對照差異不顯著,然而添加外源NaCl顯著提高了正常水分和水分脅迫下的CAT活性。
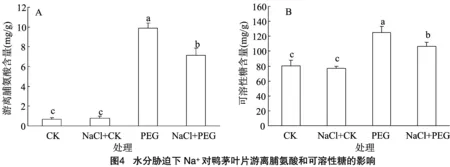
2.4 水分脅迫下Na+對鴨茅葉片內有機滲透調節物質的影響
游離脯氨酸和可溶性糖是植物體內重要的有機滲透調節物質。從圖4-A可以看出,正常水分條件下(CK和CK+NaCl處理)鴨茅葉片內游離脯氨酸含量均較低,且差異不顯著;然而在水分脅迫條件下(PEG和PEG+NaCl)含量均急劇增加,其中PEG處理的脯氨酸含量高于PEG+NaCl處理,差異達顯著水平。同時,水分脅迫使鴨茅葉片積累了更多的可溶性糖,其中PEG+NaCl處理的可溶性糖含量顯著低于直接水分脅迫處理(圖4-B)。
3 討論與結論
干旱是限制植物生長和發育最主要的逆境因子之一,水分脅迫往往會造成植物體內大量活性氧的積累,導致膜質過氧化,其中MDA是膜質過氧化作用的產物之一,MDA含量高低和細胞質膜透性變化是反映細胞膜脂過氧化作用強弱和質膜破壞程度的重要指標,被廣泛地用作氧化損傷的分析[19]。同時,電解質滲透率直接反映植物細胞膜穩定性,其值大小是反映干旱脅迫對植物的傷害程度[20]。抗氧化酶系統是主要清除活性氧的保護酶系統,通過催化超氧陰離子發生歧化作用而轉化為H2O2和O2,并將不同細胞定位的H2O2轉化為H2O和O2,該系統需要活性氧清除酶系統的關鍵酶SOD、CAT、POD發揮協同作用,以降低氧化脅迫導致的植物毒害[21]。本研究中,正常水分條件下添加外源Na+處理鴨茅葉片內MDA含量、電解質滲透率和活性氧成分均未發生明顯變化,說明 50 mmol/L NaCl沒有對鴨茅造成氧化脅迫傷害,而根系活力、CAT活性顯著高于對照CK,該結果與李州等的研究結果[4,22]一致,即吸收適量的Na+有助于促進植物生長。在水分脅迫條件下添加外源NaCl處理能夠有效緩解了水分脅迫導致的氧化損傷,顯著提高了鴨茅對水分脅迫的耐受力。該結果與部分旱生植物耐旱機制一致,蔡建一等研究表明 50 mmol/L NaCl能夠顯著提高滲透脅迫下霸王SOD、CAT、POD活性,從而減輕活性氧對細胞膜的傷害,有效降低水分脅迫對植物的傷害[7]。Martíínez等對濱藜[Atriplexpatens(Litv. ) Iljin]進行干旱處理發現,適量的Na+可以減輕水分脅迫造成的氧化脅迫,提高植物適應環境的能力[23]。由此看來,Na+在適量水平時,對植物的抗逆性有重要作用。
長期以來,無機滲透調節物質Na+被認為是造成植物鹽害的主要因素,因此對Na+的研究主要集中在植物鹽害方面。近年來,越來越多的研究關注于Na+對旱生植物的滲透調節作用。已有研究表明,Na+在荒漠植物梭梭[Haloxylonammodendron(C. A. Mey.) Bunge]的滲透調節中貢獻最大,是梭梭適應干旱環境時最主要的滲透調節物質[24]。然而適量的Na+對中生植物耐旱性研究比較少,缺乏充足的證據。本試驗結果顯示,水分脅迫下加入50 mmol/L NaCl可以使鴨茅葉片內游離脯氨酸和可溶性糖含量顯著降低。這與張金林等的結果較為一致,即中生植物和多漿旱生植物并不依靠脯氨酸來調節滲透勢,Na+對滲透調節的貢獻要遠大于K+、Ca2+和有機滲透調節物質等[25-26]。由此可以推測,鴨茅在響應外源Na+進行滲透調節時可能與多槳旱生植物具有相似特性[27],通過積累滲透調節物質Na+,可以平衡液泡滲透勢,減少合成其他有機滲透調節物質,維持植物生長和抵御干旱脅迫。
綜上所述,50 mmol/L NaCl預處理能顯著增強水分脅迫下鴨茅葉片抗氧化防御系統,有效緩解水分脅迫導致的細胞氧化性損傷。外源NaCl處理顯著提高鴨茅抗旱性的原因可能與旱生植物一樣是通過富集Na+提高其滲透調節能力來發揮作用,并非促進了植物體內有機滲透調節物質的積累。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
